Nonalcoholic fatty liver disease (NAFLD) encompasses a broad spectrum of pathological hepatic conditions ranging from simple steatosis to nonalcoholic steatohepatitis (NASH), which may predispose to liver cirrhosis and hepatocellular carcinoma (HCC). The pathogenesis of NAFLD is closely related to insulin resistance (IR), adiposity and physical inactivity as well as genetic and epigenetic factors corroborate to the development and progression of hepatic steatosis and liver injury. Emerging evidence has outlined the implication of gut microbiota and gut-derived endotoxins as actively contributors to NAFLD pathophysiology probably due to the tight anatomo-functional crosstalk between the gut and the liver. Obesity, nutrition and environmental factors might alter intestinal permeability producing a favorable micro-environment for bacterial overgrowth, mucosal inflammation and translocation of both invasive pathogens and harmful byproducts, which, in turn, influence hepatic fat composition and exacerbated pro-inflammatory and fibrotic processes.
- nonalcoholic fatty liver disease
- gut microbiota
- gut–liver axis
- leaky gut
- intestinal permeability
- tight junctions
- endotoxemia
- probiotics
1. Introduction
2. Insight into the Gut Microbiota in NAFLD
The human gastrointestinal lumen is the physiological habitat for more than 100 trillion microorganisms, which is approximately ten-times the number of somatic cells in the human body, hosting a wide variety of microbial species (archaea, fungi, yeast, bacteria and viruses) [16]. The gut microflora is a large reservoir of commensal microbes that live synergistically with the host and provide biological and metabolic functions benefiting the host. It includes more than 160 different bacterial species, including anaerobes and they carry more than three million unique genes [17,18][17][18]. Among them, bacteria predominate with the phyla of the Gram-positive Firmicutes and Gram-negative Bacteroidetes, mainly involved in the short-chain fatty acids (SCFAs), i.e., acetate, butyrate and propionate and hydrogen production, respectively. The other phyla are represented by Actinobacteria, Fusobacteria, Proteobacteria and Verrucomicrobia [19,20,21][19][20][21]. The precise function of the intestinal flora remains largely uncharted. However, it processes complexed and indigestible polysaccharides to SCFAs, providing energy to the host and it also participates in vitamin (i.e., vitamin B and K), bile acid and amino acid synthesis, drug and toxin metabolism and intestinal barrier preservation. In particular, the term ‘dysbiosis’ indicates all imbalances between beneficial and pathogen bacteria or modifications in intestinal flora taxonomic composition and/or function [22]. Perturbations in intestinal microbiota homeostasis has been already described not only in NAFLD, but also in ALD [10], T2DM [23], obesity [24,25][24][25] and many other diseases [26,27,28,29][26][27][28][29]. Along the gastrointestinal tract (GIT) from the mouth to colon, the bacterial concentration and composition is strikingly diverse (increasing from stomach to colon), showing even higher variability depending on the age, lifestyle, medications and diets. Indeed, a diet enriched in animal fat and sugars as well as the Western diet may predispose to bacterial overgrowth, immune system activation and mucosal inflammation both in preclinical [30,31][30][31] and clinical studies [32,33][32][33]. Several approaches have been developed to study the intestinal flora community diversity, exploiting quantitative real time polymerase chain reaction (qRT-PCR), sequencing of the 16S ribosomal RNA (rRNA) gene through next-generation DNA sequencing or partial 16S rRNA sequencing in the V6–V8 region through pyrosequencing, excepting for Enterobacteriaceae and Enterococcaceae families [34]. These tools provide information about the abundance and the taxonomy of microbial species in mucosa-associated colonic tissue biopsies and in fecal samples. All these techniques are also coupled with the more expensive metagenomics or metatranscriptomics shotgun approaches [35]. Nonetheless, to study the host-microbiome interactions, intestinal, systemic, uric, and fecal bacterial-products and metabolites, such as bile acids, SCFAs and endotoxins, can be assessed by using proteomic and metabolomic methods and may represent diagnostic noninvasive markers, reflecting the microbiota composition [36].3. Gut–Liver Axis: New Awareness in NAFLD Pathogenesis and Progression
The gut–liver axis has many implications in NAFLD onset as the major contributor of the intestinal dysbiosis, possibly due to the tight anatomo-functional crosstalk of the two organs. The liver is perpetually exposed to gut microbial end-products and nutrients via the portal vein (70% of blood supply) and, in turn, participates to bacterial composition through bile acids cycling released into the duodenum lumen with the enterohepatic circulation [10]. Alongside, the gut microbiome composition is crucial to modulate innate and adaptive immune response both locally and systemically, facilitating host defense against pathogens. The bowel wall plays an essential role as a selective barrier that regulates the bidirectional flux between the gut and the liver, since it is constituted by tight and adherent junctions (occludins, claudins and Zonula Occludens 1 (ZO-1)) and desmosomes, which hold together the epithelial cells. Furthermore, it exerts many immunological functions as it is constituted by multiple layers and specialized cells, such as Goblet, Paneth and plasma cells secreting mucus, antimicrobial peptides (i.e., defensins, lysozyme and c-lectin Reg3b/g) and Immunoglobulin A (IgA), respectively. Together they protect the host from invasive pathogens and avoid bacterial overgrowth and systemic translocation [10]. The excessive erosion of the protective mucus layer as well as the reduction of antimicrobial mediators has been associated with translocation of pathogenic microorganisms in both preclinical and human studies [70,71][37][38]. Disturbance of the intestinal barrier integrity, a phenomenon known as leaky gut, along with shifting in metabolic function of gut microbiota, are frequently present in patients with NAFLD-related dysbiosis [51,72][39][40] and correlate with NAFLD severity. Indeed, a relative abundance in Bacteroides and Ruminococcus have been independently associated with NASH and fibrosis [73][41]. As a consequence of enhanced gut permeability, much more bacteria and potentially harmful byproducts translocate into circulation and reach the liver thus contributing to the increase of circulating gut-derived toxins (endotoxemia) and the establishment of chronic low-grade inflammatory state that features metabolic disorders such as obesity and NAFLD [74,75][42][43]. Several endogenous molecules as ethanol, ammonia and acetaldehyde, whose circulating increased levels result from dysbiotic microbiota (i.e., Escherichia coli abundance), are able to stimulate hepatic Kupffer cells to produce pro-inflammatory cytokines with similar mechanisms occurring in ALD [10,57][10][44]. Likewise, LPS and peptidoglycans derived from Gram-negative and Gram-positive bacteria walls are the most representative pathogen-associated molecular patterns (PAMPs), which activate Toll-like receptors (TLRs) signaling. In particular, LPS-induced TLR-4 cascade in hepatocytes, Kupffer cells and hepatic stellate cells (HSCs) leading to elevated systemic levels of tumor necrosis factor alpha (TNF-α) and interleukin 6 (IL6) via nuclear receptor kappa B (NF-kB) thus promoting IR, inflammation and fibrosis [9,76][9][45]. Otherwise, circulating free fatty acids (FFAs), whose levels are commonly higher in NAFLD, may independently stimulate TLR4 and TLR2 inflammatory pathways [77,78][46][47]. Furthermore, peptidoglycans and damage-associated molecular patterns (DAMPs) contribute to liver damage through the crosstalk between TLRs (e.g., TLR2 and TLR5) and inflammasome via intracellular nucleotide-binding and oligomerization domain (NOD)-like receptors (NLRs), which increase IL1 and IL8 production in hepatocytes, Kupffer cells and HSCs [60][48]. Moreover, alteration of gut microflora communities contributes to liver pathology and disruption of intestinal barrier integrity. For instance, dysbiosis may affect lipid metabolism and trafficking in both liver and adipose tissue by upregulating lipogenic enzymes or lipoprotein lipase (LPL) thus participating to obesity and steatosis development. Interestingly, several intestinal bacteria species dampen the production of the Fating-Induced Adipocyte Factor (FIAF), whose downregulation is associated with increased adiposity and hepatic de novo lipogenesis [79][49]. Enrichment in Cytophaga–Flavobacter–Bacteroides phyla influences the development of fatty liver and hepatic inflammation favoring IL7 release from T-helper cells (Th17) [80][50]. Dietary choline is further metabolized by enteric bacteria in trimethylamine and then it is converted in the hepatotoxic trimethylamine N-oxide (TMAO) end-product. Indeed, choline shortage or increased TMAO production have been associated with higher levels of Gram-negative Gammaproteobacteria and Erysipelotrichi and with steatosis since its levels are crucial to favor very-low density lipoprotein (VLDL) assembly and secretion. Microbial SCFAs may affect the intestinal barrier integrity and mucosal immune tolerance raising levels of intestinal SCFAs-producing species strengthen barrier integrity supporting tight junctions and mucins production and operating as energy source for intestinal mucosal cells [81,82][51][52]. For example, the reduction of produce butyric acid, produced by Faecalibacterium prausnitzii, weakens the few connections between intestinal epithelial cells, by decreasing the expression of the tight junction proteins and mucins. The restoration of the physiological abundance of microorganisms-producing butyrate, in turn, may ameliorate the gut high permeability and systemic inflammation [83][53]. The molecular features of the gut–liver axis in NAFLD/NASH are schematically represented in Figure 1A.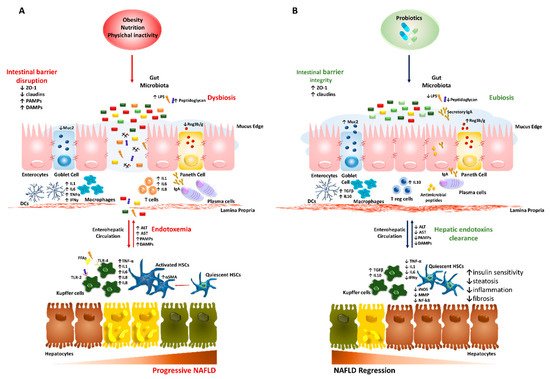
4. Probiotics: Cunning Double-Crossers Against Their Household
References
- Day, C.P. From fat to inflammation. Gastroenterology 2006, 130, 207–210.
- Wong, R.J.; Aguilar, M.; Cheung, R.; Perumpail, R.B.; Harrison, S.A.; Younossi, Z.M.; Ahmed, A. Nonalcoholic steatohepatitis is the second leading etiology of liver disease among adults awaiting liver transplantation in the United States. Gastroenterology 2015, 148, 547–555.
- Younossi, Z.M. Non-alcoholic fatty liver disease—A global public health perspective. J. Hepatol. 2019, 70, 531–544.
- Byrne, C.D.; Targher, G. NAFLD: A multisystem disease. J. Hepatol. 2015, 62, S47–S64.
- Dongiovanni, P.; Valenti, L. A Nutrigenomic Approach to Non-Alcoholic Fatty Liver Disease. Int. J. Mol. Sci. 2017, 18, 1534.
- Dongiovanni, P.; Romeo, S.; Valenti, L. Genetic Factors in the Pathogenesis of Nonalcoholic Fatty Liver and Steatohepatitis. BioMed Res. Int. 2015, 2015, 460190.
- Dongiovanni, P.; Meroni, M.; Longo, M.; Fargion, S.; Fracanzani, A.L. miRNA Signature in NAFLD: A Turning Point for a Non-Invasive Diagnosis. Int. J. Mol. Sci. 2018, 19, 3966.
- Meroni, M.; Longo, M.; Rametta, R.; Dongiovanni, P. Genetic and Epigenetic Modifiers of Alcoholic Liver Disease. Int. J. Mol. Sci. 2018, 19, 3857.
- Buzzetti, E.; Pinzani, M.; Tsochatzis, E.A. The multiple-hit pathogenesis of non-alcoholic fatty liver disease (NAFLD). Metab. Clin. Exp. 2016, 65, 1038–1048.
- Meroni, M.; Longo, M. Alcohol or Gut Microbiota: Who Is the Guilty? Int. J. Mol. Sci. 2019, 20, 4568.
- Del Chierico, F.; Nobili, V.; Vernocchi, P.; Russo, A.; De Stefanis, C.; Gnani, D.; Furlanello, C.; Zandona, A.; Paci, P.; Capuani, G.; et al. Gut microbiota profiling of pediatric nonalcoholic fatty liver disease and obese patients unveiled by an integrated meta-omics-based approach. Hepatology 2017, 65, 451–464.
- Delarue, J.; Lalles, J.P. Nonalcoholic fatty liver disease: Roles of the gut and the liver and metabolic modulation by some dietary factors and especially long-chain n-3 PUFA. Mol. Nutr. Food Res. 2016, 60, 147–159.
- Bibbo, S.; Ianiro, G.; Dore, M.P.; Simonelli, C.; Newton, E.E.; Cammarota, G. Gut Microbiota as a Driver of Inflammation in Nonalcoholic Fatty Liver Disease. Mediat. Inflamm. 2018, 2018, 9321643.
- Puri, P.; Sanyal, A.J. The Intestinal Microbiome in Nonalcoholic Fatty Liver Disease. Clin. Liver Dis. 2018, 22, 121–132.
- Borrelli, A.; Bonelli, P.; Tuccillo, F.M.; Goldfine, I.D.; Evans, J.L.; Buonaguro, F.M.; Mancini, A. Role of gut microbiota and oxidative stress in the progression of non-alcoholic fatty liver disease to hepatocarcinoma: Current and innovative therapeutic approaches. Redox Biol. 2018, 15, 467–479.
- Hillman, E.T.; Lu, H.; Yao, T.; Nakatsu, C.H. Microbial Ecology along the Gastrointestinal Tract. Microbes Environ. 2017, 32, 300–313.
- Guarner, F.; Malagelada, J.R. Gut flora in health and disease. Lancet 2003, 361, 512–519.
- Proctor, L.M. The Human Microbiome Project in 2011 and beyond. Cell Host Microbe 2011, 10, 287–291.
- Eckburg, P.B.; Bik, E.M.; Bernstein, C.N.; Purdom, E.; Dethlefsen, L.; Sargent, M.; Gill, S.R.; Nelson, K.E.; Relman, D.A. Diversity of the human intestinal microbial flora. Science 2005, 308, 1635–1638.
- Ley, R.E.; Backhed, F.; Turnbaugh, P.; Lozupone, C.A.; Knight, R.D.; Gordon, J.I. Obesity alters gut microbial ecology. Proc. Natl. Acad. Sci. USA 2005, 102, 11070–11075.
- Ley, R.E.; Hamady, M.; Lozupone, C.; Turnbaugh, P.J.; Ramey, R.R.; Bircher, J.S.; Schlegel, M.L.; Tucker, T.A.; Schrenzel, M.D.; Knight, R.; et al. Evolution of mammals and their gut microbes. Science 2008, 320, 1647–1651.
- Bajaj, J.S. Alcohol, liver disease and the gut microbiota. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 235–246.
- Larsen, N.; Vogensen, F.K.; van den Berg, F.W.; Nielsen, D.S.; Andreasen, A.S.; Pedersen, B.K.; Al-Soud, W.A.; Sorensen, S.J.; Hansen, L.H.; Jakobsen, M. Gut microbiota in human adults with type 2 diabetes differs from non-diabetic adults. PLoS ONE 2010, 5, e9085.
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 2006, 444, 1027–1031.
- Turnbaugh, P.J.; Backhed, F.; Fulton, L.; Gordon, J.I. Diet-induced obesity is linked to marked but reversible alterations in the mouse distal gut microbiome. Cell Host Microbe 2008, 3, 213–223.
- Hold, G.L.; Smith, M.; Grange, C.; Watt, E.R.; El-Omar, E.M.; Mukhopadhya, I. Role of the gut microbiota in inflammatory bowel disease pathogenesis: What have we learnt in the past 10 years? World J. Gastroenterol. 2014, 20, 1192–1210.
- Kassinen, A.; Krogius-Kurikka, L.; Makivuokko, H.; Rinttila, T.; Paulin, L.; Corander, J.; Malinen, E.; Apajalahti, J.; Palva, A. The fecal microbiota of irritable bowel syndrome patients differs significantly from that of healthy subjects. Gastroenterology 2007, 133, 24–33.
- Nadal, I.; Donat, E.; Ribes-Koninckx, C.; Calabuig, M.; Sanz, Y. Imbalance in the composition of the duodenal microbiota of children with coeliac disease. J. Med. Microbiol. 2007, 56, 1669–1674.
- Goodman, B.; Gardner, H. The microbiome and cancer. J. Pathol. 2018, 244, 667–676.
- Martinez-Medina, M.; Denizot, J.; Dreux, N.; Robin, F.; Billard, E.; Bonnet, R.; Darfeuille-Michaud, A.; Barnich, N. Western diet induces dysbiosis with increased E coli in CEABAC10 mice, alters host barrier function favouring AIEC colonisation. Gut 2014, 63, 116–124.
- Agus, A.; Denizot, J.; Thevenot, J.; Martinez-Medina, M.; Massier, S.; Sauvanet, P.; Bernalier-Donadille, A.; Denis, S.; Hofman, P.; Bonnet, R.; et al. Western diet induces a shift in microbiota composition enhancing susceptibility to Adherent-Invasive E. coli infection and intestinal inflammation. Sci. Rep. 2016, 6, 19032.
- Singh, R.K.; Chang, H.W.; Yan, D.; Lee, K.M.; Ucmak, D.; Wong, K.; Abrouk, M.; Farahnik, B.; Nakamura, M.; Zhu, T.H.; et al. Influence of diet on the gut microbiome and implications for human health. J. Transl. Med. 2017, 15, 73.
- Hills, R.D., Jr.; Pontefract, B.A.; Mishcon, H.R.; Black, C.A.; Sutton, S.C. Gut Microbiome: Profound Implications for Diet and Disease. Nutrients 2019, 11, 1613.
- Suchodolski, J.S.; Dowd, S.E.; Wilke, V.; Steiner, J.M.; Jergens, A.E. 16S rRNA gene pyrosequencing reveals bacterial dysbiosis in the duodenum of dogs with idiopathic inflammatory bowel disease. PLoS ONE 2012, 7, e39333.
- Aguiar-Pulido, V.; Huang, W.; Suarez-Ulloa, V.; Cickovski, T.; Mathee, K.; Narasimhan, G. Metagenomics, Metatranscriptomics, and Metabolomics Approaches for Microbiome Analysis. Evolut. Bioinform. Online 2016, 12, 5–16.
- Yang, Y.; Misra, B.B.; Liang, L.; Bi, D.; Weng, W.; Wu, W.; Cai, S.; Qin, H.; Goel, A.; Li, X.; et al. Integrated microbiome and metabolome analysis reveals a novel interplay between commensal bacteria and metabolites in colorectal cancer. Theranostics 2019, 9, 4101–4114.
- Lindstedt, G.; Lindstedt, S.; Gustafsson, B.E. Mucus in Intestinal Contents of Germfree Rats. J. Exp. Med. 1965, 121, 201–213.
- Wang, L.; Fouts, D.E.; Starkel, P.; Hartmann, P.; Chen, P.; Llorente, C.; DePew, J.; Moncera, K.; Ho, S.B.; Brenner, D.A.; et al. Intestinal REG3 Lectins Protect against Alcoholic Steatohepatitis by Reducing Mucosa-Associated Microbiota and Preventing Bacterial Translocation. Cell Host Microbe 2016, 19, 227–239.
- Miele, L.; Valenza, V.; La Torre, G.; Montalto, M.; Cammarota, G.; Ricci, R.; Masciana, R.; Forgione, A.; Gabrieli, M.L.; Perotti, G.; et al. Increased intestinal permeability and tight junction alterations in nonalcoholic fatty liver disease. Hepatology 2009, 49, 1877–1887.
- Giorgio, V.; Miele, L.; Principessa, L.; Ferretti, F.; Villa, M.P.; Negro, V.; Grieco, A.; Alisi, A.; Nobili, V. Intestinal permeability is increased in children with non-alcoholic fatty liver disease, and correlates with liver disease severity. Dig. Liver Dis. Off. J. Ital. Soc. Gastroenterol. Ital. Assoc. Stud. Liver 2014, 46, 556–560.
- Boursier, J.; Diehl, A.M. Implication of gut microbiota in nonalcoholic fatty liver disease. PLoS Pathogens 2015, 11, e1004559.
- Cani, P.D.; Osto, M.; Geurts, L.; Everard, A. Involvement of gut microbiota in the development of low-grade inflammation and type 2 diabetes associated with obesity. Gut Microbes 2012, 3, 279–288.
- Boulange, C.L.; Neves, A.L.; Chilloux, J.; Nicholson, J.K.; Dumas, M.E. Impact of the gut microbiota on inflammation, obesity, and metabolic disease. Genome Med. 2016, 8, 42.
- Zhu, L.; Baker, S.S.; Gill, C.; Liu, W.; Alkhouri, R.; Baker, R.D.; Gill, S.R. Characterization of gut microbiomes in nonalcoholic steatohepatitis (NASH) patients: A connection between endogenous alcohol and NASH. Hepatology 2013, 57, 601–609.
- Schnabl, B.; Brenner, D.A. Interactions between the intestinal microbiome and liver diseases. Gastroenterology 2014, 146, 1513–1524.
- Ferreira, D.F.; Fiamoncini, J.; Prist, I.H.; Ariga, S.K.; de Souza, H.P.; de Lima, T.M. Novel role of TLR4 in NAFLD development: Modulation of metabolic enzymes expression. Biochim. Biophys. Acta 2015, 1851, 1353–1359.
- Cai, C.; Zhu, X.; Li, P.; Li, J.; Gong, J.; Shen, W.; He, K. NLRP3 Deletion Inhibits the Non-alcoholic Steatohepatitis Development and Inflammation in Kupffer Cells Induced by Palmitic Acid. Inflammation 2017, 40, 1875–1883.
- Kolodziejczyk, A.A.; Zheng, D.; Shibolet, O.; Elinav, E. The role of the microbiome in NAFLD and NASH. EMBO Mol. Med. 2019, 11.
- Leung, C.; Rivera, L.; Furness, J.B.; Angus, P.W. The role of the gut microbiota in NAFLD. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 412–425.
- Ivanov, I., II; Frutos Rde, L.; Manel, N.; Yoshinaga, K.; Rifkin, D.B.; Sartor, R.B.; Finlay, B.B.; Littman, D.R. Specific microbiota direct the differentiation of IL-17-producing T-helper cells in the mucosa of the small intestine. Cell Host Microbe 2008, 4, 337–349.
- Brahe, L.K.; Astrup, A.; Larsen, L.H. Is butyrate the link between diet, intestinal microbiota and obesity-related metabolic diseases? Obes. Rev. Off. J. Int. Assoc. Stud. Obes. 2013, 14, 950–959.
- Topping, D.L.; Clifton, P.M. Short-Chain Fatty Acids and Human Colonic Function: Roles of Resistant Starch and Nonstarch Polysaccharides. Physiol. Rev. 2001, 81, 1031–1064.
- Rivière, A.; Selak, M.; Lantin, D.; Leroy, F.; De Vuyst, L. Bifidobacteria and Butyrate-Producing Colon Bacteria: Importance and Strategies for Their Stimulation in the Human Gut. Front. Microbiol. 2016, 7, 979.
- Bluemel, S.; Williams, B.; Knight, R.; Schnabl, B. Precision medicine in alcoholic and nonalcoholic fatty liver disease via modulating the gut microbiota. Am. J. Physiol. Gastrointest. Liver Physiol. 2016, 311, G1018–G1036.
- Markowiak, P.; Slizewska, K. Effects of Probiotics, Prebiotics, and Synbiotics on Human Health. Nutrients 2017, 9, 1021.
- Paolella, G.; Mandato, C.; Pierri, L.; Poeta, M.; Di Stasi, M.; Vajro, P. Gut-liver axis and probiotics: Their role in non-alcoholic fatty liver disease. World J. Gastroenterol. 2014, 20, 15518–15531.