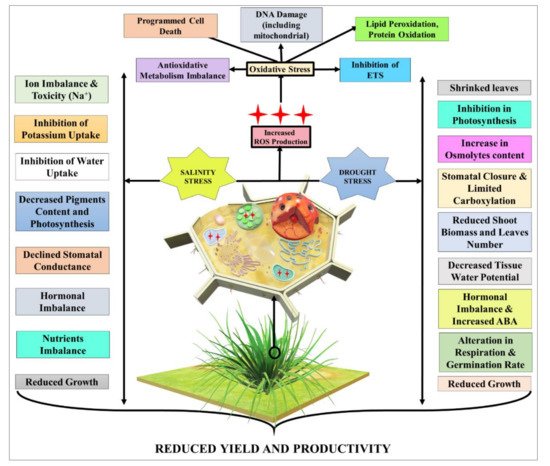
Abiotic stress in plants is a crucial issue worldwide, especially heavy-metal contaminants, salinity, and drought. These stresses may raise a lot of issues such as the generation of reactive oxygen species, membrane damage, loss of photosynthetic efficiency, etc. that could alter crop growth and developments by affecting biochemical, physiological, and molecular processes, causing a significant loss in productivity. To overcome the impact of these abiotic stressors, many strategies could be considered to support plant growth including the use of nanoparticles (NPs). However, the majority of studies have focused on understanding the toxicity of NPs on aquatic flora and fauna, and relatively less attention has been paid to the topic of the beneficial role of NPs in plants stress response, growth, and development. More scientific attention is required to understand the behavior of NPs on crops under these stress conditions.
- abiotic stresses
- environmental contaminants
- heavy metals
- nanoparticles
- soil
1. Introduction
2. Alleviation of Heavy Metal Toxicity in Plants Using Nanoparticles
| Nanoparticles | Plants | Germination and Morphological Responses | Physiological Responses | References | ||
|---|---|---|---|---|---|---|
| Si (10 μM) | Pisum sativum | L. | Presence of Si NPs improved the growth in presence of Cr | Si NPs minimized the Cr storage, enhanced the synthesis of defense enzymes and augmented nutrient uptake | [57] | [69] |
| ZnO (25 mg/L) | Leucaena leucocephala |
| Nanoparticles | Plants | Germination and Morphological Responses | Physiological Responses | References | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ag (0, 2, 5, and 10 mM) | Triticum aestivum | L. | Seed priming with Ag NPs significantly augmented the fresh and dry biomass of salinity stressed wheat plants at all doses compared to the control. | Ag NPs increased the activities of vital antioxidative enzymes whilst declined the contents of stress indicators, i.e., MDA and H | 2 | O | 2 | in wheat leaves as compared to salt stressed plants. | [83] | [85] | ||||||||||||
| Application of NPs induced seedling growth | ZnO NPs amendment improved pigments and soluble proteins, reduced peroxidation; there was rise in the antioxidant defense enzymes | [ | 58 | |||||||||||||||||||
| Zn-, B-, Si-, and Zeolite NPs | Solanum tuberosum | L., Diamont cultivar | Application of individual and binary treatment of NPs improved plant height, shoot dry weight, number of stems per plant, and tuber yield as compared to the control. | NP treatment increased leaf relative water content, leaf photosynthetic rate, leaf stomatal conductance, and chlorophyll content in comparison to the control; improved nutrients contents, leaf proline content, and leaf gibberellic acid level; and enhanced the contents of protein adn carbohydrates, and antioxidative enzymes’ activities. | ] | [70] | ||||||||||||||||
| [ | 85 | ] | [ | 87] | Fe | 3 | O | 4 | Triticum aestivum | L. | Fe | 3 | O | 4 | NP treatment minimized the inhibitory action of HMs | Fe | 3 | O | 4 | NPs supplementation improved the level of superoxide dismutase and peroxidase | [59] | [71] |
| Fe (0, 0.08, and 0.8 ppm), and potassium silicate (0, 1, and 2 mM) |
Vitis vinifera | — | Application of NPs significantly increased the total protein content, activities of antioxidative enzymes (POD, CAT, and SOD), and hydrogen peroxide, while reduced proline content. | [86] | [88] | Si (19, 48, and 202 nm) | Oryza sativa | L. | Si NPs enhanced the number of cultured cells and decreased proportionally with the rise in NP size; the treatment maintained the cellular integrity in the presence of metals | Si NPs amendment caused altered expression of genes responsible for reduced metal uptake | [60] | [72] | ||||||||||
| ZnO (0, 50, 75, and 100 mg/L) | Zea mays | |||||||||||||||||||||
| Rise in plant height and shoot dry weight | ||||||||||||||||||||||
| Increase in N and P content; reduced Cd transport, rise in the level of vital ions and antioxidant pool | ||||||||||||||||||||||
| [ | 66 | ] | [ | 58] | ||||||||||||||||||
| Cu (0, 25, 50, and 100 mg kg | −1 | of soil) | ||||||||||||||||||||
| Fe (0.0, 0.08, and 0.8 ppm) | Fragaria ananassa | Application of Fe NPs (at higher concentrations) increased root dry weight and dry weight of the explants. | Fe NPs improved the contents of photosynthetic pigments and total soluble carbohydrate, membrane stability index, and relative water content of salinity-stressed plants. | [87] | [89] | L. | Treatment caused rise in plant length, leaf number, and biomass | ZnO NPs application enhanced chlorophyll content, gas exchange characteristics, and antioxidant enzymes; addition led to reduced content of Cd in root and shoot | [61] | [73] | ||||||||||||
| T | . aestivum | |||||||||||||||||||||
| N–Na | 2 | SiO | ZnO (0, 25, 50, 75, and 100 mg/L) and Fe NPs (0, 5, 10, 15, and 20 mg/L) | T. aestivum | L. | Treatment induced plant growth, dry weight, and grains under Cd stress | Addition of NPs decreased the loss of electrolyte and activity of superoxide dismutase and peroxidase along with diminished Cd accumulation | [62] | [74] | |||||||||||||
| 3 | L. | Improved biomass and growth | Reduced Cr availability; increase in nutrient uptake; rise in antioxidant content | [67] | [57] | |||||||||||||||||
| (400 ppm) | S. tuberosum | L. | Foliar spraying of N–Na | 2 | SiO | 3 | restored the tuber number per plant and tuber yield along with improved water use efficiency and tuber dry matter percentage under salinity stress. | Application of N–Na | 2 | SiO | 3 | exerted positive impacts on the quantum yield of PS II, carotenoids content, and DPPH radical scavenging activity in salinity stressed plants. | [88] | [90] | ||||||||
| SiO | 2 | (0, 50, 100, and 150 mg/L) | Musa acuminata | All doses of SiO | 2 | NPs improved the number of shoots and shoot length of banana. | Application of SiO | 2 | NPs increased chlorophyll content, lowered electrolyte leakage, reduced MDA content, and altered the content of phenolic compounds | [89] | [91] | Si | Glycine max | L. | Si NPs minimized the growth inhibitory action of Hg | Incorporation of Si NPs improved the chlorophyll content and reduced the Hg content in root and shoot | [63 | |||||
| CNPs (0.3% and 90–110 nm) | Lactuca sativa | ] | ||||||||||||||||||||
| The salinity-induced deleterious effects on germination and associated parameters were alleviated by the exposure of C NPs, e.g., treatment of C NPs for 2 h significantly improved the germination rate in some varieties. | — | [ | 90 | ] | [92] | Mel-Au (200 μM) | O. sativa | L. | — | Application of Mel-Au NPs caused reduction of Cd level in root and shoot, improved chlorophyll content and raised the activity of antioxidant enzymes | [64] | |||||||||||
| ZnO (0, 1000, and 3000 ppm) | Trigonella | foenum-graecum | — | Interaction of NaCl and ZnO was recorded to reverse the salinity induced consequences (L-proline, protein, MDA, aldehydes, sugars, H | 2 | O | 2 | , and antioxidative enzymes) in both cultivars, but the results were more apparent in case cv. Ardestanian than cv. Mashhadian. | [91] | [93] | Fe (25 and 50 mg/L) | |||||||||||
| ZnO (10, 50, and 100 mg/L) | O. sativa | L. | Treatment of Fe NPs improved plant length and dry weight | Fe NPs application caused rise in the level of proline, glutathione and phyto-chelatins; Fe NPs addition led to improved defense enzymes and glyoxalase machinery | Lycopersicon esculentum | [19] | ||||||||||||||||
| Foliar spraying of ZnO NPs increased shoot length and root length, biomass, and leaf area. | Increased chlorophyll content and photosynthetic attributes, protein content, and activities of antioxidative enzymes (POX, SOD, and CAT) in salinity-stressed tomato plants. | [ | 92 | ] | [94] | ZnO (10–100 mg/L) | O. sativa | L. | Amendment of ZnO increased the growth of seedlings | Treatment facilitated reduced accumulation of arsenic in root and shoot together with rise in phytochelatin level | [65] | |||||||||||
| TiO | 2 | , (40, 60, and 80 ppm) | [ | 75 | Zea mays | L. | ] | |||||||||||||||
| Seed priming with TiO | 2 | positively impacted the germination (germination percentage, germination energy, and seedling vigor index) and seedling growth (lengths of root and shoot, fresh, and dry weight) and reduced the mean emergence time. | Results showed the enhancement in potassium ion concentration, relative water content, contents of total phenolic and proline contents; increased SOD, CAT, and PAL activities; and decreased sodium ion concentration, membrane electrolyte leakage, and MDA content. | [53] | Cu (25, 50, and 100 mg kg | −1 | of soil) | T. aestivum | L. | Fe | 2 | O | 3 | (0, 25, 50, and 100 mg kg | −1 | soil) | O. sativa | L. | Improved fresh and dry biomass; increased height | Augmented detoxifying enzymes, photosynthetic potential, and nutrient uptake attributes; reduced formation of ROS, lowered expression of genes supporting the transport of Cd; restricted Cd mobilization in upper plant parts | [68] | [59] |
| Fe | 2 | O | 3 | (25, 50, and 100 mg kg | −1 | soil) | T. aestivum | L. | Rise in plant fresh and dry biomass; increase in plant length | Reduced Cd transport; enhanced N, P, and K content; increased antioxidants and pigment content | [69] | [60] | ||||||||||
| TiO | 2 | (0, 100, and 250 mg/L soil) | Z. mays | Foliar application improved shoot and root dry weight | Reduced accumulation of Cd; increased activities of antioxidant enzymes | [70] | [61] | |||||||||||||||
| SiO | 2 | (30 and 50 nm) | G. max | Improved seedling fresh weight | Improved chlorophyll content; lowered accumulation of Hg in root | [63] | ||||||||||||||||
| Au (200 μM) | O. sativa L. | — | Reduced level of Cd in root and leaves by 33 and 46.2%, respectively; improvement in antioxidant defense enzyme; restricted expression of genes associated with metal transport | [64] | ||||||||||||||||||
| Si (0, 25, 50, and 100 mg/kg soil) | T. aestivum L | . | Improved plant height | Improved chlorophyll; photosynthesis; diminished Cd content in tissues; | [64] | |||||||||||||||||
| ZnO (0, 50, and 100 mg L | −1 | ) | G. max | Improved root and shoot growth | Reduced arsenic concentration in root and shoot; improved photosynthesis, water loss, photochemical yield; raised antioxidative defense enzymes | [71] | [66] | |||||||||||||||
| Ti (0.1 to 0.25%) | Vigna radiata L | . | Augmented radicle length and biomass | Decline in the level of ROS and lipid peroxidation; upregulation of genes related with antioxidative enzymes | [72] | [67] | ||||||||||||||||
| Se and Si (5, 10, and 20 mg L | −1 | ) | O. sativa L. | — | Lowered accrual of Cd and Pb; improved yield | [73] | [68] |
3. Alleviation of Salinity Stress in Plant Using Nanoparticles