Age-related bone disorders such as osteoporosis or osteoarthritis are a major public health problem due to the functional disability for millions of people worldwide. Furthermore, fractures are associated with a higher degree of morbidity and mortality in the long term, which generates greater financial and health costs. As the world population becomes older, the incidence of this type of disease increases and this effect seems notably greater in those countries that present a more westernized lifestyle. Thus, increased efforts are directed toward reducing risks that need to focus not only on the prevention of bone diseases, but also on the treatment of persons already afflicted. Evidence is accumulating that dietary lipids play an important role in bone health which results relevant to develop effective interventions for prevent bone diseases or alterations, especially in the elderly segment of the population.
- dietary lipids
- hallmarks of aging
- oxidative stress
- saturated fat diet
Note: The following contents are extract from your paper. The entry will be online only after author check and submit it.
1. Introduction
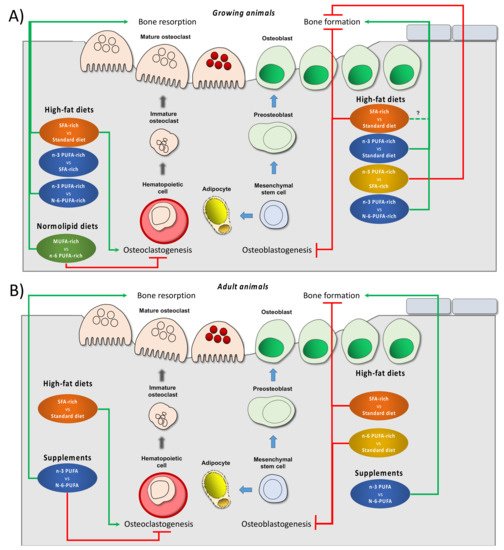
2. Dietary Lipids and Bone Health
| Model; Age | Experimental Diet vs. Control Diet;Duration | Main Changes vs. Control Diet | Ref. | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Growing animals | ||||||||||||
| Male Wistar rat; 3 wks | SFA-HFD | al | (40%E beef tallow) vs. SD | al | (AIN-93G with soybean oil asa fat source); 8 wks |
| [14] | [38] | ||||
| n-6 PUFA-rich HFD | al | (40%E soybean oil) vs. SFA-HFD | al | (40%E beef tallow); 8 wks |
| [14] | [38] | |||||
| n-6 PUFA-rich HFD | al | (40%E corn oil) vs. SFA-HFD | al | (40%E beef tallow); 8 wks |
| [14] | [38] | |||||
| n-3 PUFA- rich HFD | al | (40%E linseed oil) vs. SFA-HFD | al | (40%E beef tallow); 8 wks |
| [14] | [38] | |||||
| n-3 PUFA- rich HFD | al | (40%E linseed oil) vs. SD | al | (AIN-93G with soybean oil as fat source); 8 wks |
| [14] | [38] | |||||
| Male Wistar rats; Weanling | MUFA-rich SD | al | (AIN-93 diet with 9.5%E extra virgin olive oil) vs. n-6 PUFA-rich SD | al | (AIN-93 diet 9.5%E sunflower oil); 6 m |
| [22] | [46] | ||||
| MUFA-rich SD | al | (AIN-93 diet with 9.5%E extra virgin olive oil) vs. n-6 PUFA-rich SD | al | (AIN-93 diet 9.5%E sunflower oil); 6 m |
| [22] | [46] | |||||
| MUFA-rich SD | al | (AIN-93 diet with 9.5%E extra virgin olive oil) vs. n-6 PUFA-rich SD | al | (AIN-93 diet 9.5%E sunflower oil); 24 m |
| [23] | [47] | |||||
| n-3 PUFA-rich SD | al | (AIN-93 diet 9.5%E fish oil); vs. n-6 PUFA-rich SD | al | (AIN-93 diet 9.5%E sunflower oil); 24 m |
| [23] | [47] | |||||
| MUFA-rich SD | al | (AIN-93 diet with 9.5%E extra virgin olive oil) vs. n-3 PUFA-rich SD | al | (AIN-93 diet 9.5%E fish oil); 24 m |
| [23] | [47] | |||||
| Male Sprague-Dawley rats; Weanling | HFD | al | (AIN93G with added 70 g/kg of safflower oil + menhaden oil) with n-6/n-3 PUFA ratio = 23.8, 9.8 2.6 or 1.2; 42 d |
| [24] | [48] | ||||||
| Female Sprague Dawley rats; 4 wks | HFD | al | (12wt% tuna oil) vs. HFD | al | (12wt% corn oil); 8 wks |
| [15] | [39] | ||||
| HFD | al | (12wt% flaxseed oil) vs. HFD | al | (12wt% corn oil); 8 wks |
| [15] | [39] | |||||
| HFD | al | (12wt% menhaden oil) vs. HFD | al | (12wt% corn oil); 8 wks |
| [15] | [39] | |||||
| Male C57BL/6J mice; 5 wks | SFA-HFD | al | (60%E lard) vs. SD | al | (D12450B chow with 10%E fat); 12 wks |
| [16] | [40] | ||||
| Male C57BL/6J mice; 6 wks | SFA-HFD | al | (60%E lard) vs. SD | al | (LabDiet 5LOD with 13.5% E lard); 20 wks |
| [13] | [16] | ||||
| Male C57BL/6J mice; 6 wks | SFA-HFD | al | (60%E lard) vs. SD | al | (12%E fat); 12 wks |
| [17] | [41] | ||||
| Male C57BL/6 mice; 4 wks | SFA-HFD | al | (21.2%E lard) vs. SD | al | (4.8%E fat); 13 wks |
| [25] | [49] | ||||
| Male BALB/cByJ mice; 7 wks | SFA-HFD | al | (45%E lard) vs. SD | al | (13.5%E fat); 15 wks |
| [18] | [42] | ||||
| Male Wistar rats; 9 wks | SFA-HFD | al | (24% fat with 100 g/kg of bw per day of ground nut and 50 g/kg of bw per day dried coconut) vs. SD | al | ; 38 wks |
| [26] | [50] | ||||
| Male Sprague-Dawley rat; 40 d | SFA-HFD | al | (20wt% coconut oil) vs. n-3 PUFA-HFD | al | (20wt% flaxseed oil) or n-6 PUFA-HFD | al | (20wt% safflower oil); 65 d |
| [27] | [51] | ||
| Newly hatched chicks; 4 d | n-3-rich PUFA diet | al | (menhaden oil + safflower oil at 90 g/kg) vs. n-6-rich PUFA diet | al | (soybean oil + safflower oil at 90 g/kg); 17 d |
| [28] | [52] | ||||
| Adult animals | ||||||||||||
| Male Sprague-Dawley rat; 200 g | Cholesterol-enriched HFD | al | (10.0 g cholesterol, 20.0 g sodium-cholate, and 112.0 g crude fat %per kg dry matter) vs. SD | al | (50.83 g crude fat %per kg dry matter); 114 d |
| [10] | [35] | ||||
| Male F344 × BNF1 rats; 12 m | n-3 PUFA-rich diet (167 g safflower oil + 33 g menhaden oil) vs. n-6 PUFA-rich diet (200 g safflower oil) or n-3 + n-6 PUFA-rich diet (190 menhaden oil + 10 g corn oil); 20 wks |
| [29] | [53] | ||||||||
| Male F344 × BNF1 rats; 12 m | n-3 PUFA-rich diet (167 g safflower oil + 33 g menhaden oil) vs. n-6 PUFA-rich diet (200 g safflower oil) or n-3 + n-6 PUFA-rich diet (190 menhaden oil + 10 g corn oil); 20 wks |
| [20] | [44] | ||||||||
| Male C57BL/6J mice; 8 wks | SFA-HFD | al | (35wt% lard) vs. SD | al | (6wt% fat); 20 wks |
| [30] | [54] | ||||
| Male C57BL/6 mice; 3 m | HFD | al | (45%E) vs. SD | al | (12%E fat); 11 wks |
| [31] | [55] | ||||
| Male C57BL/6J; 4 m | HFD | al | (45%E) vs. SD | al | (11%E fat); 8 wks |
| [32] | [56] | ||||
| Female C57BL/6J mice; 8 m | SFA-HFD | al | (45%E lard) vs. SD | al | (10%E fat); 8 wks |
| [8] | [33] | ||||
| MUFA-rich HFD | al | (45%E olive oil) vs. SFA-HFD | al | (45%E lard); 8 wks |
| [8] | [33] | |||||
| Female C57BL/6J mice; 13 m | n-6 PUFA-rich HFD | al | (19.5%E corn oil) al vs. SD | al | (9.5%E fat); 26 wks |
| [33] | [57] | ||||
| n-3 PUFA-rich HFD | al | (19.5%E fish oil) vs. n-6 PUFA-rich HFD | al | (19.5%E corn oil); 26 wks |
| [33] | [57] | |||||
| Roosters; 40 wks | HFD | al | (8% palmitic acid) vs. LFD | al | (8% cellulose); 20 wks |
| [21] | [45] | ||||
| C57BL/6 mice | Cholesterol-enriched HFD | al | (15.8wt% fat + 1.25wt% cholesterol) vs. SD | al | (6wt% fat), 7 m |
| [11] | [36] | ||||
| C3H/HeJ mice | Cholesterol-enriched HFD | al | (15.8wt% fat + 1.25wt% cholesterol) vs. SD | al | (6wt% fat), 7 m |
| [11] | [36] | ||||
| Aged animals | ||||||||||||
| OVX female Sprague Dawley; 3 m | DHA-rich diets (HP5 and LP5) High-PUFA diet vs. low-PUFA diet with a ratio of n-6/n-3 PUFAs of 5:1 or 10:1 (110.4 g/kg of fat from safflower oil (110.4 g/kg of high-oleate safflower oil blended with n-3 PUFAs); 12 wks | Fatty acid analyses confirmed that the dietary ratio of 5:1 significantly elevated the amount of DHA in the periosteum, marrow and cortical and trabecular bones of the femur. |
|
Serum osteocalcin was lowest in the HP groups. Regardless of the dietary PUFA content, DHA in the 5:1 diets (HP5 and LP5) preserved rat femur BMC in the absence of estrogen |
[34] | [58] | ||||||
| OVX female C57BL/6J mice; 8 wks | AIN-93 diet | al | (10%E virgin olive oil) vs. AIN-93 diet | al | (10%E refined olive oil); 4 wks |
| [35] | [59] | ||||
| Marrow stromal cells from C57BL/6 mice fed a high fat, atherogenic diet failed to undergo osteogenic differentiation in vitro | [12] | [37] | ||||||||||
| Female Wistar rats; 56 m | HFD | al | (31%E peanut + canola seed oil); 19 wks |
| [35] | [59] | ||||||
| Population; Age | Intervention vs. Control Diet/Placebo; Duration | Main Changes vs. Control Diet or Placebo | Ref. | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 42 women and 23 men; 51.3 ± 7.1 y | HFD (61%E fat) vs. SD (30%E fat); 12 m |
| [52] | [78] | ||||||
| 208 women and 99 men; 45.5 + 9.7 y | HFD (>45%E fat) vs. SD (30%E fat); 24 m |
| [53] | [79] | ||||||
| 242 women and 182 men; 51.8 ± 8.9 y | HFD (40%E fat) vs. LFD (20%E fat); 24 m |
| [54] | [80] | ||||||
| 236 women; 44–50 y | SD (32%E fat) vs. LFD (24%E fat); 18 m |
| [68] | [94] | ||||||
| 230 women; 27.3 + 1.1 y | LFD | dc | (<28%E fat) + vs. SD | dc | (30%E fat); 108 m |
| [55] | [81] | ||
| n-3 PUFA supplementation | ||||||||||
| 3 women and 20 men; 49.3 + 1.6 y | n-3 PUFA rich HFD (37.6%E fat; 6.5%E ALA; n-6/n-3 ratio: 1.6/1) vs. HFD (34.5%E fat; 0.8%E ALA; n-6/n-3 ratio: 9.5/1); 6 wks |
| [57] | [83] | ||||||
| n-3 PUFA rich HFD (37.6%E fat; 6.5%E ALA; n-6/n-3 ratio: 1.6/1) vs. n-6 PUFA rich HFD (37.1%E fat; 3.6%E ALA; n-6/n-3 ratio: 3.5/1); 6 wks |
| [57] | [83] | |||||||
| 43 women and 2 men with RA; 57.9 ± 10.8 | NCD + n-3 PUFA supplement (2.4 g of n-3 PUFA/d; 1.1 g ALA+ 0.7 g EPA + 0.1 g DPA + 0.4 g DHA) vs. NCD + dairy supplement (2.4 g of SFA/d); 3 m |
| [61] | [87] | ||||||
| 87 woman and 26 men; 18–67 y | NCD + n-3 PUFA supplement (1.48 g EPA + DHA/d) vs. NCD + placebo (NA g olive oil); 12 wks |
| [59] | [85] | ||||||
| 75 women and 6 men with RA; 49.24 ± 10.46 y | NCD + n-3 PUFA supplement (2.090 g of EPA and 1.165 g of DHA/d) vs. NCD + placebo (NA g of high-oleic-acid sunflower oil); 16 wks |
| [58] | [84] | ||||||
| 126 women; 75 ± 7 y | NCD + n-3 PUFA supplement (1.2 g EPA + DHA/d) vs. NCD + placebo (NA g olive oil); 6 m |
| [60] | [86] | ||||||
| 60 women and 15 men; 35–65 y | NCD + n-3 PUFA/MUFA enriched dairy supplement (23.7 g saturated fat + 5.17 g oleic acid + 0.14 g DHA + 0.20 g EPA/d) vs. NCD + semiskimmed milk (70g saturated fat + 2.05 g oleic acid); 12 m |
| [62] | [88] | ||||||
| n-6 PUFA supplementation | ||||||||||
| 3 women and 20 men; 49.3 + 1.6 y | n-6 PUFA rich HFD (37.1%E fat; 12.6%E LA; n-6/n-3 ratio: 3.5/1) vs. HFD (34.5%E fat; 7.7%E LA; n-6/n-3 ratio: 9.5/1); 6 wks |
| [57] | [83] | ||||||
| n-6 PUFA rich HFD (37.1%E fat; 12.6%E LA; n-6/n-3 ratio: 3.5/1) n-3 PUFA rich HFD (37.6%E fat; 10.5%E LA; n-6/n-3 ratio: 1.6/1); 6 wks |
| [57] | [83] | |||||||
| 38 women and 6 men with RA; 46.2 ± 13.1 y | NCD + n-6 PUFA supplement (2.5 g CLA/d) vs. NCD + placebo (2.5 g of high-oleic-acid sunflower oil); 3 m |
| [64] | [90] | ||||||
| 60 men; 49.1+ 6.2 y | NCD + n-6 PUFA supplement (3 g CLA/d) vs. NCD + placebo (NA g palm and bean oil blend/d); 8 wks |
| [63] | [89] | ||||||
| MUFA supplementation | ||||||||||
| 127 men; 67.9 ± 6.9 | CD + EVOO supplementation (>50 mL EVOO/d) vs. CD + nuts supplementation (30 g of mixed walnuts, almonds and hazelnuts); 2 y |
| [67] | [93] | ||||||
| 98 women and 104 men; 67.8 ± 6.5 | CD + EVOO supplementation (15L EVOO/3 m) vs. CD + nuts supplementation (1350 g of mixed walnuts, almonds and hazelnuts/3 m); 1 y |
| [65] | [91] | ||||||
| 104 women and 7 men severely obese; 18–40 y | CD + EVOO supplementation (52 mL EVOO/d) vs. CD; 12 wks |
| [66] | [92] | ||||||