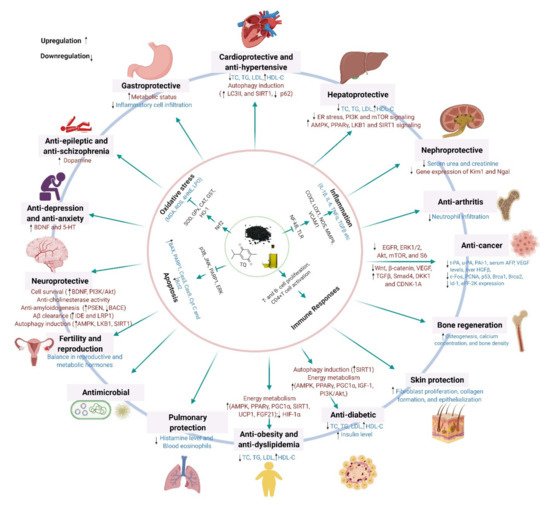
Black cumin (Nigella sativa L.), a highly valued nutraceutical herb with a wide array of health benefits, has attracted growing interest from health-conscious individuals, the scientific community, and pharmaceutical industries. The pleiotropic pharmacological effects of black cumin, and its main bioactive component thymoquinone (TQ), have been manifested by their ability to attenuate oxidative stress and inflammation, and to promote immunity, cell survival, and energy metabolism, which underlie diverse health benefits, including protection against metabolic, cardiovascular, digestive, hepatic, renal, respiratory, reproductive, and neurological disorders, cancer, and so on. Furthermore, black cumin acts as an antidote, mitigating various toxicities and drug-induced side effects.
- black seed
- thymoquinone
- nutraceutical
- essential oil
- molecular mechanism
1. Introduction
2. Phytochemical Profiles
3. Benefits of Black Cumin on Human Health and Disease Conditions
3.1. Antioxidant Effects
3.2. Anti-Inflammatory Effects
3.3 Protection against Neurological Disorders
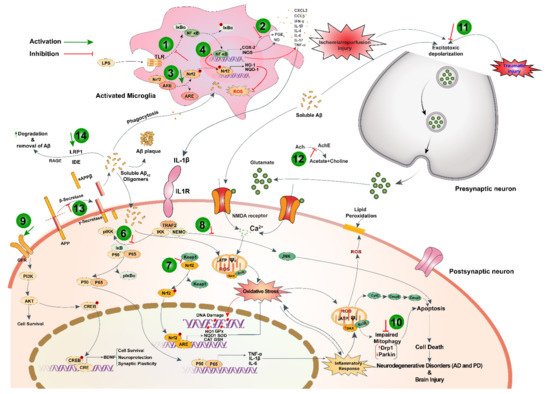
Black cumin and TQ have shown their therapeutic promises against a range of neurological conditions, including neurodegenerative disorders (Alzheimer’s disease (AD), and Parkinson’s disease (PD)), ischemic stroke and acute brain injury, anxiety and depression, epilepsy, and schizophrenia (Table 1). Moreover, black cumin and TQ were shown to protect against various chemical-induced neuronal injury in experimental conditions (Table 1). The neuroprotective potentials of black cumin and TQ mostly stem from antioxidative and anti-inflammatory properties [21][47] (Figure 1Figure 3).
3.4 Anti-Cancer Effects
| Treatment with Doses | Experimental Model | Major Findings (Including Molecular Changes) |
References |
|---|---|---|---|
| Neuroinflammation | |||
| TQ (12.5 μM for 24 h) |
LPS/IFNγ or H2O2-activated BV-2 microglial cell | ↓H2O2; ↑GSH; ↑SOD and CAT | [14] |
| TQ (12.5 μM for 24 h) |
LPS/IFNγ or H2O2-activated BV-2 microglial cell | ↑Glutaredoxin-3, biliverdin reductase A, 3-mercaptopyruvate sulfurtransferase, and mitochondrial Lon protease; ↓IL-2, IL-4, IL-6, IL-10, and IL-17a, CFB, CXCL3 and CCL5 | [22] |
| TQ (2.5–10 μM) |
LPS-activated neuroinflammation in BV-2 microglial cell | ↓ROS; ↑LKB1 and AMPK; ↑nuclear accumulation of SIRT1 | [23] |
| Alzheimer’s disease | |||
| TQ (100 nM) |
Aβ1–42-induced neurotoxicity in hiPSC-derived cholinergic neurons | ↑GSH; ↓ROS; ↓synaptic toxicity, attenuate cell death and apoptosis | [24] |
| TQ fraction rich nanoemulsion of seeds (TQRFNE) (250 and 500 mg/kg BW) |
High fat/cholesterol diet-induced neurotoxicity in rats | ↓Aβ40 and Aβ42; ↑APP; ↓PSEN1 and PSEN2; ↓BACE1 and RAGE; ↑IDE and LRP1 | [25] |
| TQ fraction rich nanoemulsion of Nigella seeds (TQRFNE) (250 and 500 mg/kg BW) |
High fat/cholesterol diet-induced neurotoxicity in rats | ↓Memory impairment; ↓lipid peroxidation and soluble Aβ levels; ↑total antioxidant status and antioxidants genes expression | [26] |
| TQ (10, 20, and 40 mg/kg/day p.o. for 14 days) |
Combined AlCl3andD-Gal-induced AD in rats | Improved cognitive deficits; ↓Aβ formation and accumulation; ↓TNF-α and IL-1β; ↓TLRs pathway components; ↓NF-κB and IRF-3 mRNAs | [27] |
| TQ (intragastrically, 20 mg/kg/day once daily for 14 days) |
Combined AlCl3 and D-Gal induced neurotoxicity in rats | ↑ Memory performance; ↑ SOD; ↓TAC; ↓MDA; ↓NO; ↓TNF-α; ↓AChE activity; ↑BDNF and Bcl-2 | [28] |
| TQ (intragastrically, 20 mg/kg/day for 15 days) |
Aβ (1–42) infused rat model of AD | ↓Memory performance (Morris water maze test); ↓IFN-γ; ↑ DCX and MAP2 | [29] |
| Parkinson’s disease | |||
| TQ (100 nM) |
α-Synuclein-induced rat hippocampal and hiPSC-derived neurons | ↑Synaptophysin; ↓synaptic vesicle recycling; ↑spontaneous firing activity | [30] |
| TQ (10 mg/kg BW, 1 week prior to MPTP at 25 mg/kg BW) |
MPTP-induced mouse PD model | ↓MDA; ↑GSH; ↑SOD; ↑CAT; ↓IL-1β and IL-6; ↓TNF-α; ↓COX-2 and iNOS; ↓α-synuclein aggregation | [31] |
| TQ (7.5 and 15 mg/kg/day, p.o.) |
Rotenone-induced rat PD model | ↓Oxidative stress; ↑Parkin; ↓ Drp1; ↑dopamine; ↑TH levels | [32] |
| Ischemic stroke | |||
| Hydroalcoholic seed extract (20 mg/kg BW) |
Global ischemia in rats | ↓Brain edema and infarct volume; ↑VEGF, HIF and MMP9 | [33] |
| TQ | Stroke-prone spontaneously hypertensive rats | ↓Chemoattractant protein-1, Cox-2, IL-1β, and IL-6 | [34] |
| Traumatic brain injury | |||
| TQ (5 mg/kg/day for seven days) |
Feeney’s falling weight-induced moderate head trauma | ↑Neuron densities; ↓MDA | [35] |
| Anxiety and Depression | |||
| Ethanolic seed extract | Chronic stress-induced depression model | ↓NO | [36] |
| TQ-loaded solid lipid nanoparticle (20 mg/kg, p.o.) and TQ (20 mg/kg, p.o.) |
Chronic stress-induced depression model | ↓IL-6, TNFα; ↑BDNF; ↑5-HT; ↑IDO | [37] |
| NSO (0.2 mL/kg for 20 days) |
Stress-induced depression model | ↑Memory performance (FST) | [38] |
| Hydroalcoholic seed extract (200 and 400 mg/kg) |
Stress-induced depression and anxiety model | ↑Anxiolytic (Open field and elevated plus-maze test); ↓depression (FST) | [39] |
| Epilepsy | |||
| Ethanolic seed extract (400 mg/kg/day, p.o.) | PTZ-induced kindling mode | ↓Kindling development; ↑memory performance (Morris water maze test); ↓LTP | [40] |
| NSO (400 and 600 mg/kg BW) | Electroshock-induced seizures | ↑Anticonvulsant activity | [41] |
| TQ (10 mg/kg, i.p) |
Lithium chloride and pilocarpine-induced seizure | ↑Memory performance; ↑SOD; ↑Nrf2, HO-1 | [42] |
| TQ (10 mg/kg, i.p) |
Lithium chloride and pilocarpine-induced seizure | ↑Memory performance; ↓COX-2, TNF-α and NF-κB | [43] |
| Hydroalcoholic seed extract (200 and 400 mg/kg for 5 days) |
PTZ-induced seizure model | ↑Memory performance (Morris water maze and passive avoidance test); ↑ total thiol; ↓MDA | [44] |
| Schizophrenia | |||
| TQ (20 mg/kg, daily for 28 days, i.p.) |
Mice model of schizophrenia (haloperidol-induced catalepsy and apomorphine-induced climbing behavior) |
Anti-amnesic effect; ↓AChE activity; ↓ TBARS; ↑GSH and catalase; ↑dopamine level | [45] |
| Miscellaneous effects | |||
| Chemical-induced toxicity | |||
| TQ (5 mg/kg, i.p. for 11days) |
Acrylamide-induced neurotoxicity in rats | Improved gait abnormalities; ↑GSH; ↓MDA;↓caspases 3 and 9, and Bax/Bcl-2, pP38/P38 and pJNK/JNK; ↓pERK/ERK; restore BBB integrity | [46] |
| TQ (5 and 10 mg/kg, i.p., for 11 days) |
Acrylamide-Induced Peripheral Nervous System Toxicity in rats | Improved gait abnormalities; ↑GSH and ↓MDA;↓caspases 3 and 9, and Bax/Bcl-2, pP38/P38 and pJNK/JNK; ↓pERK/ERK | [47] |
| TQ (10 µM and 20 µM) |
Arsenic-induced cytotoxicity in SH-SY5Y cells | Promotes DNA repairing; ↓ROS, balanced transmembrane potential; ↓ Bax and PARP-1, and ↑Bcl-2 | [48] |
| TQ (5 mg/kg/day, for 3 days, p.o.) |
Arsenic-induced hippocampal toxicity in rats | Improve anxiety behavior (Open field test and elevated plus maze); ↑GSH and SOD; ↓DNA damage; ↓TNF-α and INF-γ | [49] |
| TQ (2.5 and 5 mg/kg BW, for 8 days, p.o.) |
Arsenic-induced hippocampal toxicity in Wistar rats | ↑Δψm | [50] |
| NSO (1 mL/kg BW for 7 days) |
Dichlorvos-induced oxidative and neuronal damage in rats | ↓Vacuolation in the frontal and cerebellar cortices;↑TAC and GSH↓ROS | [51] |
| Radiotoxicity | |||
| TQ | Radiation-induced oxidative stress in brain tissue | ↑Antioxidant enzymes | [52] |
3.4 Anti-Cancer Effects
Black cumin and its compounds are widely known for their potent anticancer actions. Accumulating evidence suggests that chemical constituents of black cumin seeds are chemopreventive and potent in inhibiting cell proliferation and provoking apoptosis (Table 2).
4. Molecular Mechanisms Underlying the Pharmacological Effects across Health and Disease Conditions
Black Icumin a recent study, administrand TQ were shown to exert diverse pharmacological and health effects through modulating multiple cellular signaling systems. The notable molecular pathways targeted by black cumin and TQ are Nrf2, NF-κB, TLR, SIRT1, AMPK-SIRT1-PGC-1α, PPAR, and PI3K/Akt signaling, which are shared across health/disease conditions (Figure 6).
Antioxidan t activity of black cumin seed ethanolic extractand TQ is amongst the pharmacological effects that underlie many of its health benefits and has been manifested by their capacity to enhance expression of enzymatic (such as SOD, GPx, CAT, and HO-1) and non-enzymatic (such as GSH) antioxidants, lowering various oxidative markers (such as ROS, MDA, LPO, and 4-HNE). The genetic expression of these antioxidants molecules is under the transcriptional regulation of Nrf2.
Activation of Nrf2 by either (cellular redox status or pharmacological intervention leads to the up-regulation of over 250 mg/genes encoding proteins that are involved in antioxidant defense systems, redox homeostasis, and xenobiotic detoxification [81]. Increased expression of antioxidant molecules and subsequent decline in oxidative markers by black cumin and TQ in various pharmacological effects indicate the involvement of Nrf2 activation [51,55,58,69,132,136].
In addition to its potentials on activating cellular antioxidant defense system, black cumin can directly scaveng;e free radicals, as demonstrated in several in vitro chemical assays like DPPH assay [35,292–294]. However, by virtue, TQ has a relatively poor cap.o. for 5 daysacity to quench free radicals because of its oxidized form [295]. This observation strengthens the idea that TQ can exert its antioxidant capacity by activating the Nrf2-dependent antioxidant defense system. However, thymohydroquinone, the reduced form of TQ, possesses a high radical-scavenging capacity [295]. It has been speculated that the conversion of TQ to thymohydroquinone can occur in cells and that the electron transport chain may have an important role in the antioxidant action of TQ.
While toll-like receptors (TLRs) signaling ensures protective immune response by recognizing invading pathogens and tissue-derived endogenous molecules, its overactivation perturbs the was reimmune homeostasis by sustained release of pro-inflammatory mediators and subsequently underlies the development of many inflammatory diseases [296]. TQ may improve inflammatory response in Alzheimer’s disease model by downregulating the expression of TLRs signaling components as well as their downstream effectors NF-κB and IRF-3 [54].
Modulatory role of TQ in autophagy-an evorted to attenuate diethylnitrosamine (Dlutionarily conserved cellular process that recycles defective and unwanted cell components and invading pathogens to retain cellular homeostasis has also been documented [297]. Protection against neuroinflammation by TQ in LPS-activated BV-2 microglia involved autophagy induction through activation and nuclear accumulation of SIRT1 [50]. Mitophagy, a form of autophagy that clears defective mitochondria, is regulated by parkin and Drp1 expression. An alteration of parkin and Drp1 expression may lead to impairment of mitophagy triggering apoptosis and neurodegeneration in the brain. Rotenone hindered parkin-mediated autophagy by upregulating Drp1 expression, which was ameliorated by TQ treatment [59].
The anticancer potentials of black cumin and TQ are vested on their capacity to regulate various cellular pathways that are implicated in proliferation, apoptosis, cell cycle regulation, carcinogenesis, angiogenesis, and metastasis [121]. Most of the anticancer actions of black cumin and TQ are reported to mediate by regulating cellular redox systems [117] through which both TQ and black cumin can inhibit cell proliferation, migration/invasion, and tumor growth by directly acting on growth factor signaling systems, such as ENGFR/ERK1/2, A)-induced likt/mTOR/S6, Wnt, β-catenin, and VEGF signaling [121,298,299]. TQ can prevent cancer development by its antioxidant function and can hinder cancer progression through its pro-oxidant function [121]. Besides, TQ enhanced chemosensitivity to chemotherapeutics and chemopreventive molecules by downregulating inflammatory signaling pathways and enhancing tumor-suppressing genes [116,118,120,300].
As a master upstream kinase, LKB1 phosphorylates and activater carcinogenesis and reduce serum s AMPK and many other kinases that play a fundamental role in the regulation of cell growth and metabolism [301]. The LKB1–AMPK pathway acts as a cell metabolic checkpoint, arresting cell growth under low intracellular ATP conditions, such as in nutrient-deficient states [301]. Energy overload may suppress LKB1–AMPK signaling, leading to increased cancer risk in patients with obesity or diabetes. Whereas, activation of LKB1–AMPK signaling might contribute to the suppression of cancer risk and, thus, pharmacological modulators, such as TQ, which was shown to activate LKB1–AMPK signaling [106], could have therapeutic promise in cancer prevention.
AFpart from the aforementioned mechanism, there still remain other (albeit not less significant) signaling systems that are targeted by black cumin and TQ, such as unfolded protein response (UP and VEGF levels and liver HGFβ R). Triggering of endoplasmic reticulum (ER) stress is a common phenomenon in several pathological conditions such as hypoxia/reoxygenation and oxidative stress. ER homeostasis is crucial for proteostasis and its disruption results in the buildup of unfolded and misfolded proteins in the ER lumen. Consequently, UPR is activated to resolve this protein-folding defect and thus to restore ER homeostasis. In the case of an insufficient UPR, pharmacological activation can play a therapeutic role in mitigating ER stress. Attenuation of ER stress by TQ suggests its protective role in maintaining proteostasis. Moreover, black cumin nanoemulsion promoted Aβ clearance, thus maintained protein in ratshomeostasis in the brain, by upregulating LRP1 [52], a type I transmembrane [54]glycoprotein expressed abundantly in neurons that facilitate trafficking and degradation of Aβ [302].
| Treatment with Doses | Experimental Model | Major Findings (Including Molecular Changes) |
References |
|---|---|---|---|
| Seeds incorporated silver nanoparticles (NS-AgNP) (25–200 µg/mL) |
Human breast cancer cell line (HCC-712) | Dose-dependent cytotoxicity; ↓cell density | [55] |
| Aqueous seed extract (11.5 µg/mL) |
Human breast cancer cell line (MCF-7) | Potent cytotoxic effect with IC50 11.5 µg/mL; ↑caspase-3,8 and 9, and Bax | [56] |
| NSO nanoemulsion (10–100 µL/mL) |
Human breast cancer cell line (MCF-7) | ↓Cell proliferation; ↑apoptosis and necrosis | [57] |
| TQ (25 µmol/L) |
Human breast cancer cell line (MCF-7) | Inhibit tumor cell growth; ↑p53; induce apoptosis | [58] |
| Seeds incorporated platinum nanoparticles (NS-PtNP) (25, 50, 100 and 150 µg/mL) |
HeLa cervical cancer and MDA-MB-231 breast cancer cell lines | Dose-dependent cytotoxic effect with IC50 value 36.86 µg/mL (MDA-MB-231) and 19.83 µg/mL (HeLa), respectively | [59] |
| TQ (0.78 µM) |
HeLa cervical cancer cell line | Dose-dependent antiproliferative effect | [60] |
| TQ (2, 4, 6 and 8 µM) |
Human colon cancer cell line (LoVo) | Inhibit metastasis; ↑JNK, p38; ↓P13K, ERK1/2, IKKα/β and NF-κB | [61] |
| TQ (20 µmol/L) |
Human colon cancer cell line (LoVo) | Reduce cell proliferation; ↓p-P13K, p-Akt, p-GSK3β, β-catenin and COX-2; ↓PGE2, LEF-1 and TCF-4 | [62] |
| TQ (10–120 µmol/L) |
Human bladder cancer cell lines (253J and T24) | Inhibit proliferation and metastasis; ↓MYC, Axin-2, MMP-7, MET and cyclin-D1; ↓Wnt/β-catenin signaling cascade | [63] |
| TQ (40, 60 and 80 µM) |
Human bladder cancer cell lines (253J and T24) | Significant cytotoxicity and reduction in cell proliferation; ↑caspase-3, cleaved PARP, Bax, cyt c and AIF; ↑ER-stress marker GRP78, IRE1, ATF6, ATF4 and CHOP; ↓Bcl-2 and Bcl-xl; induce apoptosis | [64] |
| TQ (10–50 µM) |
Pancreatic ductal adenocarcinoma cell lines (AsPC1 and MiaPaCa-2) | Inhibit cell viability; reduce tumor size; ↑p53, p21; ↓Bcl-2 and HDAC; induce apoptosis and G2 cell cycle arrest | [65] |
| TQ (0.5–20 µM) |
Human renal tubular epithelial cell line (HK2) and human renal cancer cell lines (769-P and 786-O) | Inhibit metastatic phenotype and epithelial-mesenchymal transition; ↑E-cadherin; ↓Snail, ZEB1 and vimentin; ↑LKB1/AMPK signaling | [66] |
| TQ (0–100 µmol/L) |
Human renal cancer cell lines (ACHN and 786-O) | Inhibition of metastasis; ↑LC3; ↑AMPK/mTOR signaling; induce autophagy | [67] |
| TQ (40 and 50 µM) |
Human kidney cancer cell lines (A498 and Caki-1) | Anti-proliferative effects with GI50 value 40.07 µM (A498) and 51.04 µM (Caki-1), respectively; ↑Bax; ↓Bcl-2 and p-Akt; induce apoptosis | [68] |
| Hexanic seed extract (0–150 µg/mL) |
Human ovary cancer cell line (A2780) | Strong cytotoxic activity of SF2 with IC50 10.89 µg/mL; ↑caspase-3 and 9; ↓MMP; induce apoptosis | [69] |
| Seed extract and NSO with OM-90(0.5 and 2.4 mg/mL) | AGS human gastric adenocarcinoma cell line | Activates mitochondrial pathways; induce apoptosis | [70] |
| TQ (0.1–30 µM) |
Human prostate cancer cell lines (PC3 and DU145) | Inhibit metastatic phenotype and epithelial-mesenchymal transition; ↓TGF-β, Smad2 and Smad3 | [71] |
| TQ (0–80 µM) |
Head and neck squamous cells carcinoma cell lines (SCC25 and CAL27) | Dose-dependent cytotoxicity with IC50 value 12.12 µM (CAL27) and 24.62 µM (SCC25), respectively; induce apoptosis | [72] |
| TQ + Resveratrol (46 µM) |
Hepatocellular carcinoma cell line (HepG2) | Significant cell inhibition; ↑caspase-3; ↓GSH and MDA; induce apoptosis | [73] |
| NSO (50–250 µg/mL) |
Human liver cancer (HepG2), human breast cancer (MCF-7), human lung cancer (A-549) and normal human embryonic kidney (HEK293) cell lines | High cytotoxic effect in HepG2 cells with IC50 48µg/mL; ↑ROS and LPO; ↓GSH and MMP; ↑p53, caspase-3 and 9, Bax; ↓Bcl-2; induce apoptosis | [74] |
| TQ (In vitro: 1–50 µMIn vivo: 20 and 100 mg/kg for 3 days; i.v.) |
TNBC cells and orthotopic TNBC xenograft mice model | Inhibit cell proliferation, migration and invasion; ↓tumor growth; ↓eEF-2K, Src/FAK and Akt | [75] |
| TQ + Paclitaxel (In vitro: 0–100 µM In vivo: 2.4 mg/kg/day for 12 days; i.p) |
Mouse breast cancer cell line (4T1) and EAC cells-induced female Balb/c mice model | Dose-dependent cytotoxicity; ↑caspase-3,7 and 12, PARP; ↓p65, p53 and Akt1; ↓JAK-STAT signaling | [76] |
| Ethanolic seed extract (250 mg/kg/day for 5 days, p.o.) |
Diethyl nitrosamine-induced hepatocarcinogenesis in Wistar rat model | Antiangiogenic effect; ↓serum VEGF and AFP levels, and liver HGFβ level | [54] |
| Ethanolic seed extract and TQ (150, 250 and 300 mg/kg (extract) 6 days/week and 20 mg/kg (TQ) for 3 days/week, p.o.) |
Diethyl nitrosamine-induced hepatocellular carcinoma in albino-Wistar rat model | Reduction in cell proliferation; ↑Antioxidant activity; ↓PCNA, c-fos, Bcl-2; ↓EGFR/ERK1/2 signaling | [77] |
| TQ + 5-fluorouracil (35 mg/kg/day for 3 days/week for 9 weeks; p.o.) |
Azoxymethane-induced colon cancer in Wistar rat model | Subdues tumor growth; ↑TGF-β1, TGF-β/RII, Smad4, DKK-1, CDNK-1A and GPx; ↓Wnt, β-catenin, NF-κB, VEGF, COX2, iNOS and TBRAS | [78] |
| TQ + Piperine (10 mg/kg/day for 14 days; i.p) |
EMT6/P cells- inoculated Balb/c mice | Inhibit angiogenesis; ↓Tumor size; ↑serum INF-ᵧ level; ↓VEGF; induce apoptosis | [79] |
| TQ + Resveratrol (50 mg/kg/day for 14 days; i.p) |
EMT6/P cells- inoculated Balb/c mice | Inhibit angiogenesis; ↓Tumor size; ↑serum INF-ᵧ level; ↓VEGF; induce apoptosis | [80] |
4. Molecular Mechanisms Underlying the Pharmacological Effects across Health and Disease Conditions
Black cumin and TQ were shown to exert diverse pharmacological and health effects through modulating multiple cellular signaling systems. The notable molecular pathways targeted by black cumin and TQ are Nrf2, NF-κB, TLR, SIRT1, AMPK-SIRT1-PGC-1α, PPAR, and PI3K/Akt signaling, which are shared across health/disease conditions (Figure 2).