Lumpy skin disease, sheeppox, and goatpox are notifiable diseases of cattle, sheep, and goats, respectively, caused by viruses of the Capripoxvirus genus. They are responsible for both direct and indirect financial losses. These losses arise through animal mortality, morbidity cost of vaccinations, and constraints to animals and animal products’ trade. Control and eradication of capripoxviruses depend on early detection of outbreaks, vector control, strict animal movement, and vaccination which remains the most effective means of control. To date, live attenuated vaccines are widely used; however, conferred protection remains controversial. Many vaccines have been associated with adverse reactions and incomplete protection in sheep, goats, and cattle. Many combination- and recombinant-based vaccines have also been developed.
- Capripoxvirus Infections in Ruminants
1. Introduction
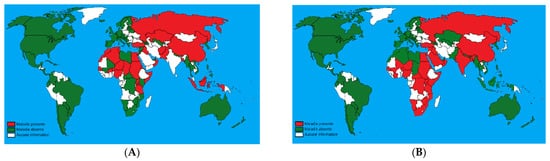
2. Clinical Signs
3. Immunity
4. Vaccination against Capripoxviruses
| Vaccine/Strain | Safety and Protection | Reference | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Sheep | Goat | Cattle | |||||||||
| GPV Gorgan | Safe, partially protective | Safe and protective | Safe and protective | [56,106,154] | [30][81][130] | ||||||
| SPV Romania | Sheep | Lamb kidney cells | 40 | [88 | |||||||
| GPV Mysore | - | ] | [ | 63] | |||||||
| Safe and protective | - | [ | 108 | ] | [83] | SPV Bakirkoy | Sheep | Calf kidney cells | 32 | [102] | [77] |
| GPV Uttarkashi | - | Safe and protective | - | [107] | [82] | SPV Rumania Fanar | Sheep | Lamb testis | 26 | [103] | [78] |
| GPV Kedong and isiolo | - | - | Safe and protective | [ | SPV Perego | Sheep | Lamb testis and calf kidney | 11 times in lamb testis and 10 times in calf kidney | [104] | [79] | |
| SPV Ranipet | Sheep | Sheep thyroid cells | 35 | [105] | [80] | ||||||
| GPV Gorgan | Goat | Kid kidney cortex cells | - | [106] | [81] | ||||||
| GPV Uttarkashi | Goat | Primary lamb testis cells and Vero cells | 34 on primary cells and 26 on Vero | [107] | [82] | ||||||
| GPV Mysore | Goats | Primary lamb testis cells | 25 | [108] | [83] | ||||||
| GPV Kedong | Sheep | Lamb testis cells | - | [109] | [84] | ||||||
| GPV Isiolo | Sheep | Lamb testis cells | - | [109] | [84] | ||||||
| LSD KSGP O 180 | Sheep | Bovine foetalmuscle cells | 18 | [110] | [85] | ||||||
| LSD KSGP O 240 | Sheep | Lamb testis cells | 13–27 | [111] | [86] | ||||||
| LSD Neethling | Cattle | Lamb kidney cells and chorioallantoic membranes (CAM) | 61 on primary cells and 20 on CAM | [99] | [74] | ||||||
| 155 | |||||||
| ] | |||||||
| [ | |||||||
| 131 | |||||||
| ] | |||||||
| SPV RM65 | Safe and protective | - | Partially protective | [21,156] | [21][132] | ||
| SPV Perego | Safe and protective | - | - | [103, | [78 | 157] | ][133] |
| SPV Rumania Fanar | Safe and protective | - | - | ||||
| SPV Romania | Safe and protective | Safe and protective | Partially protective | [57,158] | [31][134] | ||
| SPV Bakirkoy | Safe and protective | - | Partially protective | [102,115] | [77][90] | ||
| LSD Neethling | Partially protective | - | Causes Neethling disease, protective | [23,57] | [23][31] | ||
| LSD KSGP 0180 | Safe and protective | Safe and protective | Safe and protective | [110] | [85] | ||
| LSD KSGP 0240 | Safe and protective | Safe and protective | Residual virulence, partial protection | [20,112,126, | [20][87][102] | 127 |
References
- OIE. Sheep Pox and Goat Pox. Manual of Diagnostic Tests and Vaccines for Terrestrial Animals. Chapter 3.7.12. 2018. Available online: (accessed on 10 February 2021).
- Babiuk, S.; Bowden, T.R.; Boyle, D.B.; Wallace, D.B.; Kitching, R.P. Capripoxviruses: An Emerging Worldwide Threat to Sheep, Goats and Cattle. Transbound Emerg. Dis. 2008, 55, 263–272.
- Tuppurainen, E.S.M.; Venter, E.H.; Shisler, J.L.; Gari, G.; Mekonnen, G.A.; Juleff, N.; Lyons, N.A.; De Clercq, K.; Upton, C.; Bowden, T.R.; et al. Review: Capripoxvirus Diseases: Current Status and Opportunities for Control. Trans. Emerg. Dis. 2017, 64, 729–745.
- World Animal Health Information Database (WAHIS). 2020. Available online: (accessed on 26 October 2020).
- Sprygin, A.; Pestova, Y.; Wallace, D.B.; Tuppurainen, E.; Kononov, A.V. Transmission of Lumpy Skin Disease virus: A short review. Virus Res. 2019, 269, 197637.
- Buller, R.M.; Arif, B.M.; Black, D.N.; Dumbell, K.R.; Esposito, J.J.; Lefkowitz, E.J.; McFad–den, G.; Moss, B.; Mercer, A.A.; Moyer, R.W.; et al. Family Poxviridae. In Virus Taxonomy: Classification and Nomenclature of Viruses. Eighth Report of the International Committee on Taxonomy of Viruses; Fauquet, C.M., Mayo, M.A., Maniloff, J., Desselberger, U., Ball, L.A., Eds.; Elsevier: Amsterdam, The Netherlands, 2005; pp. 117–133.
- Kitching, R.P.; Mellor, P.S. Insect transmission of capripox virus. Res. Vet. Sci. 1988, 40, 255–258.
- Bowden, T.R.; Babiuk, S.L.; Parkyn, G.R.; Copps, J.S.; Boyle, D.B. Capripoxvirus tissue tropism and shedding: A quantitative study in experimentally infected sheep and goats. Virology 2008, 371, 380–393.
- Chihota, C.M.; Rennie, L.F.; Kitching, R.P.; Mellor, P.S. Mechanical transmission of Lumpy Skin Disease virus by Aedes aegypti (Diptera: Culicidae). Epidemiol. Infect. 2001, 126, 317–321.
- Chihota, C.M.; Rennie, L.F.; Kitching, R.P.; Mellor, P.S. Attempted mechanical transmission of Lumpy Skin Disease virus by biting insects. Med. Vet. Entomol. 2003, 17, 294–300.
- Tuppurainen, E.S.M.; Stoltsz, W.H.; Troskie, M.; Wallace, D.B.; Oura, C.A.L.; Mellor, P.S.; Coetzer, J.A.W.; Venter, E.H. A potential role for ixodid (hard) tick vectors in the transmission of Lumpy Skin Disease virus in cattle. Transbound. Emerg. Dis. 2011, 58, 93–104.
- Tuppurainen, E.S.; Lubinga, J.C.; Wilhelm, J.C.; Stoltsz, H.; Troskie, M.; Carpenter, S.T.; Coetzer, J.A.W.; Venter, E.H.; Oura, A.L. Mechanical transmission of Lumpy Skin Disease virus by Rhipicephalus appendiculatus male ticks. Epidemiol. Infect. 2013, 141, 425–430.
- Lubinga, J.C.; Tuppurainen, E.S.; Coetzer, J.A.; Stoltsz, W.H.; Venter, E.H. Transovarial passage and transmission of LSDV by Amblyomma hebraeum, Rhipicephalus appendiculatus and Rhipicephalus decoloratus. Exp. Appl. Acarol. 2014, 62, 67–75.
- Sohier, C.; Haegeman, A.; Mostin, L.; De Leeuw, I.; Van Campe, W.; De Vleeschauwer, A.; Tuppurainen, E.; van den Berg, T.; De Regge, N.; De Clercq, K. Experimental evidence of mechanical Lumpy Skin Disease virus transmission by Stomoxys calcitrans biting flies and Haematopota spp. horseflies. Sci. Rep. 2019, 9, 1–10.
- Babiuk, S.; Bowden, T.R.; Parkyn, G.; Dalman, B.; Hoa, D.M.; Long, N.T.; Vu, P.P.; Bieu, D.X.; Copps, J.; Boyle, D.B. Yemen and Vietnam capripoxviruses demonstrate a distinct host preference for goats compared with sheep. J. Gen. Virol. 2009, 90, 105–114.
- Kreindel, S.; Masiulis, M.; Skrypnyk, A.; Zdravkova, A.; Escher, M.; Raizman, E. Emergence of lumpy skin disease in Asia and Europe. Empres Anim. Health 2016, 46, 360.
- Hurisa, T.T.; Jing, Z.; Jia, H.; Chen, G.; He, X.B. A Review on Sheeppox and Goatpox: Insight of Epidemiology, Diagnosis, Treatment and Control Measures in Ethiopia. J. Infect. Dis. Epidemiol. 2018, 4, 057.
- Tuppurainen, E.S.; Galon, N. Lumpy Skin Disease: Current situation in Europe and neighbouring regions and necessary control measures to halt the spread in South-East Europe. Oie Reg. Comm. 2016.
- Yeruham, I.; Nir, O.; Braverman, Y.; Davidson, M.; Grinstein, H.; Haymovitch, M.; Zamir, O. Spread of Lumpy Skin Disease in Israeli dairy herds. Vet. Rec 1995, 137, 91–93.
- Ayelet, G.; Abate, Y.; Sisay, T.; Nigussie, H.; Gelaye, E.; Jemberie, S.; Asmare, K. Lumpy Skin Disease: Preliminary vaccine efficacy assessment and overview on outbreak impact in dairy cattle at debre zeit, central Ethiopia. Antivir. Res. 2013, 98, 261–265.
- Ben-Gera, J.; Klement, E.; Khinich, E.; Stram, Y.; Shpigel, N.Y. Comparison of the efficacy of Neethling Lumpy Skin Disease virus and x10RM65 sheep-pox live attenuated vaccines for the prevention of Lumpy Skin Disease–The results of a randomized controlled field study. Vaccine 2015, 33, 4837–4842.
- Boumart, Z.; Daouam, S.; Belkourati, I.; Rafi, L.; Tuppurainen, E.; Tadlaoui, K.O.; El Harrak, M. Comparative innocuity and efficacy of live and inactivated sheeppox vaccines. BMC Vet. Res. 2016, 12, 133.
- Hamdi, J.; Boumart, Z.; Daouam, S.; El Arkam, A.; Bamouh, Z.; Jazouli, M.; Tadlaoui, O.K.; Fassi Fihri, O.; Gavrilov, B.; El Harrak, M. Development and Evaluation of an Inactivated Lumpy Skin Disease Vaccine for Cattle. Vet. Microbiol 2020, 245, 108689.
- Wolff, J.; Moritz, T.; Schlottau, K.; Hoffmann, D.; Beer, M.; Hoffmann, B. Development of a Safe and Highly Efficient Inactivated Vaccine Candidate against Lumpy Skin Disease Virus. Vaccines 2021, 9, 4.
- Diallo, A.; Viljoen, G.J. Genus Capripoxvirus. In Poxviruses; Mercer, A.A., Schmidt, A., Weber, O., Eds.; Birkhäuser: Basel, Switzerland, 2007; pp. 167–181.
- Hamdi, J.; Bamouh, Z.; Jazouli, M.; Alhyane, M.; Safini, N.; Tadlaoui, K.O.; Fassi Fihri, O.; El Harrak, M. Experimental infection of indigenous North African goats with goatpox virus. Acta Vet. Scand 2021, 63, 9.
- CFSPH, Iowa State University. Sheep & Goat Pox. 2008. Available online: (accessed on 19 February 2021).
- Barnard, B.J.; Munz, E.; Dumbell, K.; Prozesky, L. Lumpy Skin Disease. In Infectious Diseases of Livestock with Special Reference to Southern Africa; Coetzer, J.A.W., Thomson, G.R., Tustin, R.C., Eds.; Oxford University Press: Oxford, UK, 1994.
- Wolff, J.; Abd El Rahman, S.; King, J.; El-Beskawy, M.; Pohlmann, M.; Beer, M.; Hoffmann, B. Establishment of a Challenge Model for Sheeppox Virus Infection. Microorganisms 2020, 8, 2001.
- Gari, G.; Abie, G.; Gizaw, D.; Wubete, A.; Kidane, M.; Asgedom, H.; Tuppurainen, E.S.M. Evaluation of the safety, immunoge- nicity and efficacy of three capripoxvirus vaccine strains against Lumpy Skin Disease virus. Vaccine 2015, 33, 3256–3261.
- Hamdi, J.; Bamouh, Z.; Jazouli, M.; Boumart, Z.; Omari Tadlaoui, K.; Fassi Fihri, O.; El Harrak, M. Experimental evaluation of the cross-protection between Sheepox and bovine Lumpy skin vaccines. Sci. Rep. 2020, 10, 8888.
- Tuppurainen, E.; Babiuk, S.; Klement, E. Lumpy Skin Disease; Springer International Publishing AG: Berlin/Heidelberg, Germany, 2018.
- Arsevska, E.; Bronner, A.; Calavas, D.; Cauchard, J.; Caufour, P.; Falala, S.; Hamon, M.; Hendrikx, P.l.; Lancelot, R.; Mercier, A.; et al. Dermatose nodulaire contagieuse des bovins: État des connaissances et situation épidémiologique dans les Balkans au 31 juillet 2016. Bull. Epid St. Anim Alim 2016, 75, 20–24.
- Omar, R. Comparison of the Two Lumpy Skin Disease Virus Vaccines, Neethling and Herbivac, and Construction of a Recombinant Herbivac-Rift Valley Fever Virus Vaccine. Master’s Thesis, University of Cape Town, Cape Town, South Africa, 2015.
- Smith, S.A.; Kotwal, G.J. Immune Response to Poxvirus Infections in Various Animals. Crit. Rev. Microbiol. 2002, 28, 149–185.
- Aristizábal, B.; González, A. Innate immune system. In Autoimmunity: From Bench to Bedside; Anaya, J.M., Shoenfeld, Y., Rojas-Villarraga, A., Eds.; El Rosario University Press: Bogota, Colombia, 2013.
- Moutaftsi, M.; Tscharke, D.C.; Vaughan, K.; Koelle, D.M.; Stern, L.; Calvo-Calle, M.; Ennis, F.; Terajima, M.; Sutter, G.; Crotty, S.; et al. Uncovering the interplay between CD8, CD4 and antibody responses to complex pathogens. Future Microbiol. 2010, 5, 221–239.
- Bonilla, F.A.; Oettgen, H.C. Adaptative immunity. J. Allergy Clin. Immunol. 2010, 125, 33–40.
- Marshall, J.S.; Warrington, R.; Watson, W.; Kim, H.L. An introduction to immunology and immunopathology. Allergy Asthma Clin. Immunol. 2018, 14, 49.
- Milovanović, M.; Dietze, K.; Milićević, V.; Radojičić, S.; Valčić, M.; Moritz, T.; Hoffman, B. Humoral immune response to repeated lumpy skin disease virus vaccination and performance of serological tests. BMC Vet. Res. 2019, 15, 80.
- Samojlović, M.; Polacek, V.; Gurjanov, V.; Lupulovic, D.; Lazic, G.; Petrovic, T.; Lazic, S. Detection of antibodies against Lumpy skin disease virus by Virus neutraliza–tion test and ELISA methods. Acta Vet. Beogr. 2019, 69, 47–60.
- Kitching, R.P.; Bhat, P.P.; Black, D.N. The characterization of African strains of capripoxvirus. Epidemiol. Infect. 1989, 102, 335–343.
- Carn, V.M. Control of capripoxvirus infections. Vaccine 1993, 11, 1275–1279.
- Seet, B.T.; Johnston, J.B.; Brunetti, C.R.; Barrett, J.W.; Everett, H.; Cameron, C.; Sypula, J.; Nazarian, S.H.; Lucas, A.; McFadden, G. Poxviruses and immune evasion. Annu. Rev. Immunol. 2003, 21, 377–423.
- Norian, R.; Ahangaran, N.A.; Varshovi, H.R.; Azadmehr, A. Evaluation of humoral and cell-mediated immunity of two capripoxvirus vaccine strains against Lumpy Skin Disease virus. Iran. J. Virol. 2016, 10, 1–11.
- Varshovi, H.R.; Norian, R.; Azadmehr, A.; Afzal Ahangaran, N. Immune response characteristics of Capri pox virus vaccines following emergency vaccination of cattle against Lumpy Skin Disease virus. IJVST 2017, 9, 33–40.
- Alberts, B. T Cells and MHC Proteins. In Molecular Biology of the Cell, 4th ed.; Garland Science: New York, NY, USA, 2002.
- Bray, M.; Wright, M.E. Progressive vaccinia. Clin Infect. Dis. 2003, 36, 766–774.
- Belyakov, I.M.; Earl, P. Shared modes of protection against poxvirus infection by attenuated and conventional smallpox vaccine viruses. Proc. Natl. Acad. Sci. USA 2003, 100, 9458–9463.
- Xu, R.; Johnson, A. Cellular and humoral immunity against vaccinia virus infection of mice. J. Immunol. 2004, 172, 6265.
- Fang, M.; Sigal, L. Antibodies and CD8+ T cells are complementary and essential for natural resistance to a highly lethal cytopathic virus. J. Immunol. 2005, 175, 6829.
- Edghill-Smith, Y.; Bray, M. Smallpox vaccine does not protect macaques with AIDS from a lethal monkeypox virus challenge. J. Infect. Dis. 2005, 191, 372–381.
- Fenner, F.; Anderson, D.A.; Arita, I.; Jezek, Z.; Ladnyi, I.D. Smallpox and Its Eradication; World Health Organization: Geneva, Switzerland, 1988.
- Redfield, R.R.; Wright, D.C. Disseminated vaccinia in a military recruit with human immunodeficiency virus (HIV) disease. N Engl. J. Med. 1987, 316, 673–676.
- Panchanathan, V.; Chaudhri, G.; Karupiah, G. Correlates of protective immunity in poxvirus infection: Where does antibody stand? Immunol. Cell Biol. 2008, 86, 80–86.
- Kennedy, J.S.; Frey, S.E.; Yan, L.; Rothman, A.L.; Cruz, J.; Newman, F.K.; Orphin, L.; Belshe, R.B.; Ennis, F.A. Induction of Human T Cell–Mediated Immune Responses after Primary and Secondary Smallpox Vaccination. JID 2004, 190, 1286–1294.
- Hammarlund, E.; Lewis, M.W.; Hansen, S.G. Duration of antiviral immunity after smallpox vaccination. Nat. Med. 2003, 9, 1131–1137.
- Hsieh, S.M.; Pan, S.C.; Chen, S.Y.; Huang, P.F.; Chang, S.C. Age distribution for T cell reactivity to vaccinia virus in a healthy population. Clin. Infect. Dis. 2004, 38, 86–89.
- Crotty, S.; Felgner, P.; Davies, H.; Glidewell, J.; Villarreal, L.; Ahmed, R. Cutting edge: Long-term B cell memory in humans after smallpox vaccination. J. Immunol. 2003, 171, 4969–4973.
- Putz, M.M.; Alberini, I.; Midgley, C.M.; Manini, I.; Montomoli, E.; Smith, G.L. Prevalence of antibodies to Vaccinia virus after smallpox vaccination in Italy. J. Gen. Virol. 2006, 86, 2955–2960.
- Agianniotaki, E.I.; Babiuk, S.; Katsoulos, P.D.; Chaintoutis, S.C.; Praxitelous, A.; Quizon, K.; Boscos, C.; Polizopoulou, Z.S.; Chondrokouki, E.D.; Dovas, C.I. Colostrum transfer of neutralizing antibodies against lumpy skin disease virus from vaccinated cows to their calves. Transbound. Emerg. Dis. 2018, 65, 2043–2048.
- Weiss, K.E. Lumpy Skin Disease. Virol. Monogr. 1968, 3, 111–131.
- Precausta, P.; Kota, F.; Vellut, G. A new freeze-dried living virus vaccine against sheep pox. Comp. Immunol. Microbiol. Infect. Dis. 1979, 1, 305–319.
- Gitao, C.G.; Mbindyo, C.; Omani, R.; Chemweno, V. Review of Sheep Pox Disease in Sheep. J. Vet. Med. Res. 2017, 4, 1068.
- Gulyaz, V. Investigation of immunity in lambs born to ewes vaccinated with sheep and goat pox vaccine. Pendik Veteriner Mikrobiyoloji Dergisi 1999, 30, 55–62.
- Rodriguez, J.F.; Janeczko, R.; Esteban, M. Isolation and characterization of neutralizing monoclonal antibodies to vaccinia virus. J. Virol. 1985, 56, 482–488.
- Gordon, J.; Mohandas, A.; Wilton, S.; Dales, S. A prominent antigenic surface polypeptide involved in the biogenesis and function of the vaccinia virus envelope. Virology 1991, 181, 671–686.
- Hsiao, J.C.; Chung, C.S.; Chang, W. Vaccinia virus envelope D8L protein binds to cell surface chondroitin sulfate and mediates the adsorption of intracellular mature virions to cells. J. Virol 1999, 73, 8750–8761.
- EFSA Panel on Animal Health and Welfare (AHAW). Scientific Opinion on sheep and goat pox. EFSA J. 2014, 12, 3885.
- EFSA Panel on Animal Health and Welfare (AHAW). Scientific Opinion on Lumpy Skin Disease. EFSA J. 2015, 13, 3986.
- Munyanduki, H.M. Improvement of the Capripoxvirus, lumpy Skin Disease Virus for Use as a Vaccine Vector. Ph.D. Thesis, Faculty of Health Sciences Department of Pathology, University of Cape Town, Cape Town, South Africa, 2018. Available online: (accessed on 19 February 2021).
- Kalra, S.K.; Sharma, V.K. Adaptation of Jaipur strain of sheeppox virus in primary lamb testicular cell culture. Indian J. Exp. Biol. 1981, 19, 165–169.
- Das, S.K.; Pandey, A.K.; Mallick, B.B. A nore on rhe nat–ural goat pox outbreak in Garwal Hills of Uttar Pradesh. Indian Vet. J. 1978, 55, 671–673.
- Van Rooyen, P.J.; Munz, E.K.; Weiss, K.E. The optimal conditions for the multi–plication of Neethling-type Lumpy Skin Disease virus in embryonated eggs. Onderstepoort J. Vet. Res. 1969, 36, 165–174.
- Bhanuprakash, V.; Hosamani, M.; Juneja, S.; Kumar, N.; Singh, R.K. Detection of goat pox antibodies, comparative efficacy of indirect ELISA and ounterimmunoelectrophoresis. J. Appl. Anim. Res. 2006, 30, 177–180.
- Ramyar, H.; Hessami, M. Development of an attenuated live vaccine against sheep-pox. Arch. Inst. Razi 1968, 20, 77–80.
- Martin, W.B.; Ergm, H.; Koylu, A. Tests on sheep of attenuated sheep pox vaccines. Res. Vet. Sci. 1973, 14, 53.
- Penkova, V.M.; Jasslm, F.A.; Thompson, J.R.; AI-Doon, T.M. The propagation of an attenuated sheep pox virus and its use as a vaccine. Bull. Off. Int. Eptzoot. 1974, 81, 329–339.
- Mateva, V.; Stoichev, S. Adaptation and cultivation of sheep-pox virus (strain Perego) in tissue cultures and evaluation of its immunogenic properties. Vet. Nauk. 1975, 12, 18–23.
- Anandan, R.; Sundara Rajan, S.; Kannamani, G.; Jayaraman, M.S. Studies on live and inactivated sheep pox vaccines. Cherion 1972, 1, 42–55.
- Abbas, F.; Khan, F.A.; Hussain, A.; Ahmed, F.; Ahmed, M.; Ahmed, S.; Ahmed, M.Z.; Attique, M.A.; Wadood, A.; Taj, M.K. Production of goatpox virus vaccine from a live attenuated goatpox virus strain. J. Anim. Plant Sci. 2010, 20, 315–317.
- Hosamani, M.; Nandi, S.; Mondal, B.; Singh, R.K.; Rasool, T.J.; Bandyopadhyay, S.K. A vero cell-attenuated goat pox virus provides protection against virulent virus challenge. Acta Virol. 2004, 48, 15–21.
- Dubey, S.C.; Sawhney, A.M. Live and reactivated tissue culture vaccines against goat pox. Indian Vet. J. 1978, 55, 925.
- Capstick, P.B.; Prydle, J.; Coackley, W.; Burdm, M.L. Protection of cattle against the Neethhng type virus of lumpy skin disease. Vet. Rec. 1959, 71, 422–423.
- Davies, F.G.; Mbugwa, G. The alterations in pathogenicity and immunogenicity of a Kenya sheep and goat pox virus on serial passage in bovine foetal cell cultures. J. Comp. Pathol. 1985, 95, 565–572.
- Davies, F.G. Characteristics of a virus causing a pox disease in sheep and goats on Kenya, with observations on the epidemiology and control. J. Hyg. (Camb.) 1976, 76, 163–171.
- Kitching, R.P.; Smale, C. Comparison of the external dimensions of capripoxvirus isolates. Res. Vet. Sci. 1986, 41, 425–427.
- Chaudhary, S.S.; Pandey, K.D.; Singh, R.P.; Verma, P.C.; Gupta, P.K. A Vero cell derived combined vaccine against sheep pox and Peste des petits ruminants for sheep. Vaccine 2009, 27, 2548–2553.
- Yogisharadhya, R.; Bhanuprakash, V.; Hosamani, M.; Venkatesan, G.; Balamurugan, V.; Bora, D.P.; Bhanot, V.; Prabhu, M.; Singh, R.K. Comparative efficacy of live replicating sheepox vaccine strains in Ovines. Biologicals 2011, 39, 417–423.
- Sevik, M.; Dogan, M. Epidemiological and Molecular Studies on Lumpy Skin Disease Outbreaks in Turkey during 2014-2015. Transbound. Emerg. Dis. 2016, 64, 1268–1279.
- Kitching, R.P. Vaccines for Lumpy Skin Disease, Sheep Pox and Goat Pox. Vaccines for OIE List A and Emerging Animal Diseases. In Proceedings of the Symposium, Ames, IA, USA, 16–18 September 2003; pp. 161–167.
- Lamien, C.E.; Lelenta, M.; Goger, W.; Silber, R.; Tuppurainen, E.; Matijevic, M.; Luckins, A.G.; Diallo, A. Real time PCR method for simultaneous detection, quantitation and differentiation of Capripoxviruses. J. Virol. Methods 2011, 171, 134–140.
- Le Goff, C.; Fakhfakh, E.; Chadeyras, A.; Aba-Adulugba, E.; Libeau, G.; Hammami, S.; Diallo, A.; Albina, E. Host-range phylogenetic grouping of capripoxviruses: Genetic typing of CaPVs. In Applications of Gene-Based Technologies for Improving Animal Production and Health in Developing Countries; Makkar, H.P.S., Viljoen, G.J., Eds.; Springer: Berlin/Heidelberg, Germany, 2005; pp. 727–733.
- Tuppurainen, E.S.M.; Pearson, C.R.; Bachanek-Bankowska, K.; Knowles, N.J.; Amareen, S.; Frost, L.; Henstock, M.R.; Lamien, C.E.; Diallo, A.; Mertens, P.P.C. Characterization of sheep pox virus vaccine for cattle against Lumpy Skin Disease virus. Antivir. Res. 2014, 109, 1–6.
- Kara, P.D.; Afonso, C.L.; Wallace, D.B.; Kutish, G.F.; Abolnik, C.; Lu, Z.; Vreede, F.T.; Taljaard, L.C.F.; Zsak, A.; Viljoen, G.J.; et al. Comparative sequence analysis of the South African vaccine strain and two virulent field isolates of Lumpy Skin Disease virus. Arch. Virol. 2003, 148, 1335–1356.
- Mathijs, E.; Vandenbussche, F.; Haegeman, A.; King, A.; Nthangeni, B.; Potgieter, C.; Maartens, L.; Van Born, S.; De Clercq, K. Complete genome sequences of the Neethling-Like Lumpy Skin Disease virus strains obtained directly from three com–mercial live attenuated vaccines. Genome Announc. 2016, 4, e01255-16.
- Katsoulos, P.D.; Chaintoutis, S.C.; Dovas, C.I.; Polizopoulou, Z.S.; Brellou, G.D.; Agianniotaki, E.I.; Tasioudi, K.E.; Chondrokouki, E.; Papadopoulos, O.; Karatzias, H.; et al. Investigation on the incidence of adverse reactions, viraemia and haematological changes following field immunization of cattle using a live attenuated vaccine against Lumpy Skin Disease. Transbound. Emerg. Dis. 2018, 65, 174–185.
- European Food Safety Authority (EFSA). Lumpy Skin Disease: I. Data collection and analysis. EFSA J. 2017, 15, 4773.
- European Food Safety Authority (EFSA); Calistri, P.; DeClercq, K.; Gubbins, S.; Klement, E.; Stegeman, A.; Cortinas Abrahantes, J.; Antoniou, S.E.; Broglia, A.; Gogin, A. Scientific report on Lumpy Skin Disease: III. Data collection and analysis. EFSA J. 2019, 17, 5638.
- Agianniotaki, E.I.; Chaintoutis, S.C.; Haegeman, A.; Tasioudi, K.E.; De Leeuw, I.; Katsoulos, P.D.; Sachpatzidis, A.; De Clercq, K.; Alexandropoulos, T.; Polizopoulou, Z.S.; et al. Development and validation of a TaqMan probe-based real-time PCR method for the differentiation of wild type Lumpy Skin Disease virus from vaccine virus strains. J. Virol. Methods 2017, 249, 48–57.
- Bedekovic, T.; Simic, I.; Kresic, N.; Lojkic, I. Detection of Lumpy Skin Disease virus in skin lesions, blood, nasal swabs and milk following preventive vaccination. Transbound. Emerg. Dis. 2017, 65, 491–496.
- Yeruham, I.; Perl, S.; Nyska, A.; Abraham, A.; Davidson, M.; Haymovitch, M.; Zamir, O.; Grinstein, H. Adverse reactions in cattle to a capripox vaccine. Vet. Rec. 1994, 135, 330–332.
- Salib, F.A.; Osman, A.H. Incidence of Lumpy Skin Disease among Egyptian cattle in Giza Governorate, Egypt. Vet. World 2011, 4, 162–167.
- Kitching, R.P.; Hammond, J.M.; Taylor, W.P. A single vaccine for the control of capripox infection in sheep and goats. Res. Vet. Sci. 1987, 42, 53–60.
- Achour, H.A.; Bouguedour, R.; Bouhbal, A.; Guechtouli, A.; Aouissat, M. Étude comparative du pouvoir immunisant de quelques souches atténuées de virus de la clavelée et d’un vaccin sensibiliser. Rev. Sci. Tech. Oie 2000, 19, 773–783.
- Bhanuprakash, V.; Hosamani, M.; Singh, R.K. Prospects of control and eradication of capripox from the Indian subcontinent: A perspective. Antivir. Res. 2011, 91, 225–232.
- Awad, M.; Michael, A.; Soliman, S.M.; Samir, S.S.; Daoud, A.M. Trials for preparation of inactivated sheep pox vaccine using binary ethyleneimine. Egypt. J. Immunol. 2003, 10, 67–72.
- Kadymov, R.A. Combined immunization of sheep against anthrax, sheep pox and clostridial infections. Vet. Mosc. 1975, 2, 50–52.
- Selyaninov, Y.O.; Balyshev, V.M.; Kosyachenko, N.S.; Egorova, I.Y. Experimental assessment of the efficacy of associated vaccine against anthrax and sheep pox. Russ. Agric. Sci. 2003, 43–45.
- Hosamani, M.; Singh, S.K.; Mondal, B.; Sen, A.; Bhanuprakash, V.; Bandyopadhyay, S.K.; Yadav, M.P.; Singh, R.K. A bivalent vaccine against goat pox and peste des petits ruminants induces protective immune response in goats. Vaccine 2006, 24, 6058–6064.
- Fakri, F.; Ghzal, F.; Daouam, S.; Elarkam, A.; Douieb, L.; Zouheir, Y.; Tadlaoui, K.; Fassi-Fihri, O. Development and field application of a new combined vaccine against Peste des Petits Ruminants and Sheep Pox. Trials Vaccinol. 2015, 4, 33–37.
- Fakri, F.Z.; Embarki, T.; Baha, W.; Tadlaoui, K.O.; Fihri, O.F. Large mass vaccination of small ruminants against Peste des Petits Ruminants and Sheeppox using a combined live attenuated vaccine. J. Vet. Med. Res. 2020, 7, 1200.
- Wallace, D.; Ellis, C.; Espach, A.; Smith, S.; Greyling, R.; Viljoen, G. Protective immune responses induced by different recombinant vaccine regimes to Rift Valley Fever. Vaccine 2006, 24, 7181–7189.
- Wallace, D.B.; Mather, A.; Kara, P.D.; Naicker, L.; Mokoena, N.B.; Pretorius, A.; Nefefe, T.; Thema, N.; Babiuk, S. Protection of Cattle Elicited Using a Bivalent Lumpy Skin Disease Virus-Vectored Recombinant Rift Valley Fever Vaccine. Front. Vet. Sci. 2020, 7, 256.
- Diallo, A.; Minet, C.; Berhe, G.; Le Goff, C.; Black, D.N.; Fleming, M.; Barrett, C.; Libeau, G. Goat immune response to Capripox vaccine expressing the hemagglutinin protein of Pest des Petits Ruminants. Ann. N. Y. Acad. Sci. 2002, 969, 88–91.
- Chen, W.; Hu, S.; Qu, L.; Hu, Q.; Zhang, Q.; Zhi, H.; Huang, K.; Bu, Z. A goat poxvirus-vectored peste-des-petits-ruminants vaccine induces long-lasting neutralization antibody to high levels in goats and sheep. Vaccine 2010, 28, 4742–4750.
- Fakri, F.; Bamouh, Z.; Ghzal, F.; Baha, W.; Tadlaoui, K.; Fihri, O.F.; Elharrak, M. Comparative evaluation of three capripoxvirus- vectored peste des petits ruminants vaccines. Virology 2018, 514, 211–215.
- Zhang, M.; Sun, Y.; Chen, W.; Bu, Z. The 135 gene of goatpox virus encodes an inhibitor of NF-kappaB and apoptosis and may serve as an improved insertion site to generate vectored live vaccine. J. Virol. 2018, 92.
- Romero, C.H.; Barrett, T.; Evans, S.A.; Kitching, R.P.; Gershon, P.D.; Bostock, C.; Black, D.N. Single Capripoxvirus recombinant vaccine for the protection of cattle against Rinderpest and Lumpy Skin Disease. Vaccine 1993, 11, 737–742.
- Wade-Evans, A.M.; Romero, C.H.; Mellor, P.; Takamatsu, H.; Anderson, J.; Thevasagayam, J.; Fleming, M.J.; Mertens, P.P.; Black, D.N. Expression of the major core structural protein (VP7) of bluetongue virus, by a recombinant capripox virus, provides partial protection of sheep against a virulent heterotypic bluetongue virus challenge. Virology 1996, 220, 227–231.
- Perrin, A.; Albina, E.; Bréard, E.; Sailleau, C.; Promé, S.; Grillet, C.; Kwiatek, O.; Russo, P.; Thiéry, R.; Zientara, S.; et al. Recombinant capripoxviruses expressing proteins of bluetongue virus: Evaluation of immune responses and protection in small ruminants. Vaccine 2007, 25, 6774–6783.
- Ma, W.; Wei, J.; Wei, Y.; Guo, H.; Jin, Y.; Xue, Y.; Wang, Y.; Yi, Z.; Liu, L.; Huang, J.; et al. Immunogenicity of the capsid precursor and a nine-amino-acid site-directed mutant of the 3C protease of foot-and-mouth disease virus coexpressed by a recombinant goatpox virus. Arch. Virol. 2014, 159, 1715–1722.
- Liu, F.; Fan, X.; Li, L.; Ren, W.; Han, X.; Wu, X.; Wang, Z. Development of recombinant goatpox virus expressing Echinococcus granulosus EG95 vaccine antigen. J. Virol. Methods 2018, 261, 28–33.
- Sun, Z.; Liu, L.; Zhang, H.; Li, Y.; Wei, F.; Li, Z.; Wang, P.; Fu, Q.; Ren, Y.; Zhang, Y.; et al. Expression and functional analysis of Brucella outer membrane protein 25 in recombinant goat pox virus. Mol. Med. Rep. 2019, 19, 2323–2329.
- Boshra, H.; Truong, T.; Nfon, C.; Bowden, T.R.; Gerdts, V.; Tikoo, S.; Babiuk, L.A.; Kara, P.; Mather, A.; Wallace, D.B.; et al. A lumpy skin disease virus deficient of an IL-10 gene homologue provides protective immunity against virulent capri–poxvirus challenge in sheep and goats. Antivir. Res. 2015, 123, 39–49.
- Molla, W.; Frankena, K.; Gari, G.; Kidane, M.; Shegu, D.; de Jong, M.C.M. Seroprevalence and risk factors of lumpy skin disease in Ethiopia. Prev. Vet. Med. 2018, 160, 99–104.
- Haegeman, A.; De Leeuw, I.; Mostin, L.; Van Campe, W.; Aerts, L.; Vastag, M.; De Clercq, K. An Immunoperoxidase Monolayer Assay (IPMA) for the detection of lumpy skin disease antibodies. J. Virol. Methods 2019, 113800.
- Haegeman, A.; De Vleeschauwer, A.; De Leeuw, I.; Vidanović, D.; Šekler, M.; Petrović, T.; Demarez, C.; Lefebvre, D.; De Clercq, K. Overview of diagnostic tools for Capripox virus infections. Prev. Vet. Med. 2019.
- Kitching, R.P. Progress towards sheep and goat pox vaccines. Vaccine 1983, 1, 4–9.
- Haqiqi, S.; Hedayati, Z.; Varshuei, H.R.; Aqa Ebrahimiyan, M.; Khedmati, K.A.D. Study of Safety and Immunogenicity of Goat Pox Vaccine against Sheep Pox in Susceptible Sheep; Agricultural Scientific Information and Documentation Centre, Agricultural Research and Education Organization: Tehran, Iran, 2008.
- Capstick, P.B.; Cocackley, W. Protection of Cattle against Lumpy Skin Disease. I. Trials with a vaccine against Neethling type infection. Cab Rev. Perspect. Agric. Vet. Sci. Nutr. Nat. Resour. 1961, 2, 362–368.
- Brenner, J.; Bellaiche, M.; Gross, E.; Elad, D.; Oved, Z.; Haimo–vitz, M.; Wasserman, A.; Friedgut, O.; Stram, Y.; Bumbarov, V.; et al. Appearance of skin lesions in cattle populations vaccinated against Lumpy Skin Disease: Statutory challenge. Vaccine 2009, 27, 1500–1503.
- Solyom, F.; Perenlei, L.; Roith, J. A live a~enuated virus vaccine against sheep pox. Acta Vet. Acad. Scient Hung. 1980, 28, 389.
- Abd-Elfatah, E.B.; El-Mekkawi, M.F.; Aboul-Soud, E.A.; Fawzi, E.M.; El-Soally, S.A. Immunological response of a new trivalent capripoxvirus vaccine in pregnant ewes and does. Slov. Vet. Res. 2019, 56 (Suppl. 22), 445–455.
- Prasad, I.J.; Datt, N.S. Observation on the use of live and inactivated vaccines ag inst goat pox. Indian Vet. J. 1973, 50, 1–10.
- Abu-Elzein, E. Observations on natural and experimental infection of sheep and goats with a virulent field Capripoxvirus with high affinity to goats. Vet. Arshiv 2003, 73, 119–131.
- Abutarbush, S.M.; Hananeh, W.M.; Ramadan, W.; Al Sheyab, O.M.; Alnajjar, A.R.; Al Zoubi, I.G.; Knowles, N.J.; Bachanek-Bankowsa, K.; Tuppurainen, E.S.M. Adverse reactions to field vaccination against Lumpy Skin Disease in Jordan. Transbound. Emerg. Dis. 2016, 63, 213–219.
- Ali, A.A.; Esmat, M.; Attia, H.; Selim, A.; Abdel-Hamid, Y.M. Clinical and patho–logical studies on Lumpy Skin Disease in Egypt. Vet. Rec. 1990, 127, 549–550.
- Abdallah, F.M.; El Damaty, H.M.; Kotb, G.F. Sporadic cases of Lumpy Skin Disease among cattle in Sharkia province, Egypt: Genetic characterization of Lumpy Skin Disease virus isolates and pathological findings. Vet. World 2018, 11, 1150–1158.
- Zeedan, G.S.G.; Mahmoud, A.H.; Abdalhamed, A.B.; Abd El-Razik, K.A.E.; Khafagi, M.H.; Abou Zeina, H.A.A. Detection of Lumpy Skin Disease virus in cattle using real-time polymerase chain reaction and serological diagnostic assays in different governorates in Egypt in 2017. Vet. World 2019, 12, 1093–1100.
- Norian, R.; Afzal Ahagran, N.; Varshovi, H.R.; Azadmehr, A. Comparative efficacy of two heterologous capripox vaccines to control Lumpy Skin Disease in cattle. Bulg. J. Vet. Med. 2017, 22, 171–179.
- Mikhael, C.A.; Ibrahim, M.M.; Saad, M.A. Efficacy of Alternative Vaccination with Attenuated Sheep Pox and Inactivated Lumpy Skin Disease Vaccines against Lumpy Skin Disease. SCVMJ 2016, 21, 125–142.
- Mikhael, C.A.; Nakhla, O.E.; Mohamed, N.A. Study on the capability of a dual capripox vaccine in protection of cattle against LSD infection. J. Vet. Med. Res. 2017, 24, 224–233.
- Aboul-Soud, E.A.; Kafafy, M.H.; El-Suall, S.A.; Zaghloul, M.A.; Mikhael, C.A. Preparation of trivalent vaccine against Lumpy Skin Disease using different capripox viral strain. Int. J. Vet. Sci. Med. 2018, in press.
- Zhugunissov, K.; Bulatov, Y.; Orynbayev, M.; Kutumbetov, L.; Abduraimov, Y.; Shayakhmetov, Y.; Taranov, D.; Amanova, Z.; Mambetaliyev, M.; Absatova, Z.; et al. Goatpox virus (G20-LKV) vaccine strain elicits a protective response in cattle against Lumpy Skin Disease at challenge with Lumpy Skin Disease virulent field strain in a comparative study. Vet. Microbiol. 2020, 245, 108695.
- Tuppurainen, E.S.M.; Oura, C.A.L. Review: Lumpy Skin Disease: An Emerging Threat to Europe, the Middle East and Asia. Trans. Emerg. Dis. 2012, 9, 40–48.