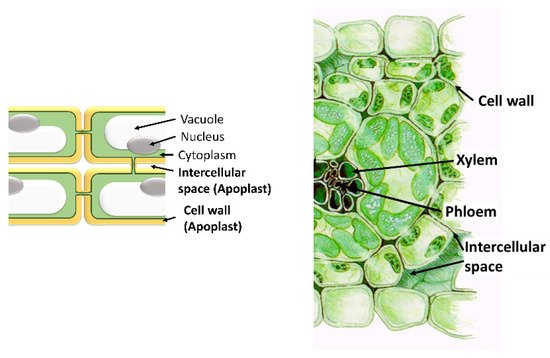
The apoplast comprises the intercellular space, the cell walls, and the xylem. Important functions for the plant, such as nutrient and water transport, cellulose synthesis, and the synthesis of molecules involved in plant defense against both biotic and abiotic stresses, take place in it. The most important molecules are ROS, antioxidants, proteins, and hormones. Even though only a small quantity of ROS is localized within the apoplast, apoplastic ROS have an important role in plant development and plant responses to various stress conditions. In the apoplast, like in the intracellular cell compartments, a specific set of antioxidants can be found that can detoxify the different types of ROS produced in it. These scavenging ROS components confer stress tolerance and avoid cellular damage. Moreover, the production and accumulation of proteins and peptides in the apoplast take place in response to various stresses. Hormones are also present in the apoplast where they perform important functions. In addition, the apoplast is also the space where microbe-associated molecular Patterns (MAMPs) are secreted by pathogens. In summary, the diversity of molecules found in the apoplast highlights its importance in the survival of plant cells.
1. Introduction
The term apoplast was coined by the German scientist E. Münch in 1930
[1]. He considered the apoplast as the intercellular space involved only in water and nutrient transport and separated the “dead” apoplast from the “living” symplast. Nowadays, the apoplast is defined as the intercellular space filled with gas and water, contained between cell membranes, the interfibrillar and intermicellar space of the cell walls, and the xylem extending to the rhizoplane and cuticle of the outer plant surface (). The apoplast is involved in a vast number of functions, since its dynamic nature allows for many reactions of great importance for the plant to take place in it. Among these functions, we can mention nutrient and water transport and the syntheses of components of the cell wall and other molecules. Among them, the synthesis of molecules involved in plant defense against both biotic (phytoalexins, PR, proteins, enzymes) and abiotic stresses are important, since it is in the apoplast where the environmental changes are detected ().
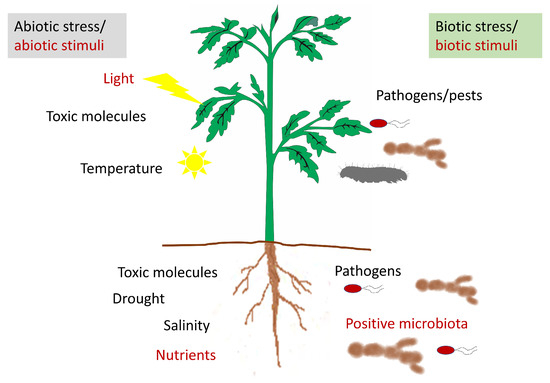
Figure 2. Representative diagram of the stresses and stimuli perceived by the plant that could modify the apoplastic content.
The synthesis of the cell wall takes place in the apoplast, after the liberation of the cell wall components through the plasma membrane. This produces the modification, degradation, and reorganization of cell wall polymers, depending on the state of cell development and differentiation. The main components of the cell wall are pectin, cellulose, and hemicellulose, but other components such as xyloglucan or phenolic compounds can be part of it. It is known that some of the steps involved in wall assembly are enzymatic, although there are other steps that seem to simply occur by chemical changes produced in the structure of its components that allow their interaction or deposition
[2][3][4][2,3,4]. The content of the cell wall is constantly changing and can be modified according to the stimuli that the plant cell is receiving through the apoplast. That way, both growth activation and the direction of growth or its arrest are regulated by the presence of certain molecules in this intercellular space
[5]. In addition to the general constituents of the cell wall, when the plant faces adverse stimuli such as different pathogens, the cell wall can be reinforced with the deposition of callose.
Since the apoplast represents the first barrier between the cell and the surrounding environment, its role in cell nutrition is also of great importance. It has been shown that the apoplast is involved in the transport of glucose produced during photosynthesis and of amino acids to the phloem
[6][7][6,7]. In the same way, Sattelmacher
[8] reviewed the role of the apoplast in plant mineral nutrition, analyzing its role in the acquisition of nutrients in root cells, and important aspects such as Na
+ toxicity and Al
3+ tolerance. Moreover, he also defined its role in long-distance transport and maintenance of the microbiota through the xylem, and short-distance storage of nutrients in the leaves. The apoplastic content of the leaves is very rich in different molecules and may vary according to the state of the plant, which points to its fundamental role in plant response to external stimuli such as the presence of pathogens
[9][10][11][9,10,11].
Hoson
[12] attributed an important role to the apoplast in the communication of plants with the external environment, not only at the level of response but also at the level of stimuli perception and their transduction, enabling, that way, plant adaptation to stress. One of the abiotic stresses to which the plant may be subjected is the presence of toxic molecules in the environment. Related to this, it was observed that the apoplastic content plays an important role in overcoming aluminum toxicity, especially through modification of its components avoiding aluminum binding to the wall
[13][14][13,14]. That way, the changes in the composition prevent the inhibition of root elongation and confer resistance against this stress. It was also shown that other abiotic stresses such as drought or salinity can induce changes in the apoplastic content that could help the plant to overcome them. Moreover, Geilfus
[15] suggested that the alkalinization of the pH of the apoplast after exposure to these stresses could play an important role since the transient increase in the pH of the apoplast reduces the stomatal aperture or even may inhibit stomatal opening via effects related to abscisic acid (ABA).
The apoplast also serves as a niche for microbes
[7][16][7,16]. On the one hand, the endophytic organisms, which tend to be present at low levels, are beneficial for the plant. It has been demonstrated recently by different authors that this symbiosis with endophytic microorganisms play an important role in growth and production, and can help the plant to overcome different stresses
[17][18][19][17,18,19].
On the other hand, the content of the apoplastic fluid is rich in nutrients that allow for populations of pathogenic microorganisms to develop in it
[9]. However, it has been shown that changes in nutrient composition and metabolism within the apoplast itself may favor pathogen control
[11][20][11,20]. The apoplastic defense is related to the set of molecules that are either present in the apoplast or secreted in the presence of the pathogen that contributes to plant defense against it
[21]. These include reactive oxygen species, toxic compounds, and anti-pathogenic protein molecules such as “pathogenesis related” (PR), other proteins, peptides, and small molecules that are secreted into the apoplastic space. In the plant defense system, through the ETI (effectors triggered immunity) plants are able to recognize the effector molecules secreted by the pathogen through immunoreceptors that are nucleotide-binding leucine-rich repeat proteins (NLR), the synthesis of which leads to a strong apoplastic defense. The interaction between apoplastic defense and control in stomatal closure has been studied, showing that NLR function is cell autonomous in guard cells for stomatal closure defense and non-cell autonomous for apoplastic defense and in the latter case it could affect the defense of the entire plant
[22].
2. Antioxidants in the Apoplast
As mentioned above, ROS can be at the same time beneficial and deleterious for the plant, since they can act as secondary messengers in different physiological processes
[23][65]. However, they can also induce oxidative damage under several environmental stress conditions, such as salinity, drought, cold, heavy metals, and UV irradiation. In the latter case, ROS accumulation may cause many cellular damages that consist of degradation of several biomolecules such as pigments, proteins, lipids, carbohydrates, and DNA, which finally leads to PCD. It was revealed that a high concentration of ROS has a toxic role in plant cells, so the actions of ROS scavengers and antioxidant enzymes are required to avoid its toxicity
[23][65]. The components of the antioxidant machinery can be broadly divided into enzymatic and non-enzymatic. The relevant ROS-scavenging enzymes are superoxide dismutase (SOD), catalase (CAT), ascorbate peroxidase (APX), monodehydroascorbate reductase (MDHAR), dehydroascorbate reductase (DHAR), and glutathione reductase (GR) (). Moreover, the non-enzymatic antioxidants such as ascorbate, glutathione, proline, phenolic compounds, and polyamines are also required to avoid cell toxicity
[24][66] ().
Superoxide dismutases (SOD) are enzymes that convert O
2•− into H
2O
2, which is less harmful to the plant. Therefore, they are considered as the first line of detoxification of ROS. SODs are classified according to the metal cofactor used by the enzyme, as manganese (Mn-SOD), iron (Fe-SOD), and copper–zinc (Cu/Zn-SOD) dependent, although Cu/ZnSOD was identified as the most active apoplastic isoform. Furthermore, many studies have reported that these enzymes confer resistance to abiotic stress. These studies highlight the fact that the tolerance level of plants is positively correlated with SOD activity and with the number of SOD isoforms. SOD is up-regulated in many plant species by various types of abiotic stress-inducing factors, such as drought, salt, and heavy metals, in a large number of crops, such as, tomato, wheat, barley, and citrus
[25][67]. Related to this, it was shown that apROS production in wheat root decreased upon a copper treatment, which was correlated with an increase in SOD activity
[26][68]. Moreover, Garcia et al.
[27][69] found a positive correlation between SOD activity and salt-tolerance when studying the response of the apoplastic antioxidant systems in root and leaf tissues from a salt-sensitive and a salt-resistant onion genotype in response to salinity. Furthermore, increased SOD activity was also observed in response to biotic stress. Vanacker et al.
[28][70] showed that the activities of several antioxidant enzymes such as SOD, CAT, APX, DHAR, MDHAR, and GR were induced in the apoplasts of barley and oat leaves 24 h after inoculation with the biotrophic fungal pathogen
Blumeria graminis [29][71]. Furthermore,
Trichoderma harzianum inoculated sunflower plants were more resistant to
Rhizoctonia solani. This resistance was accompanied by an increase in SOD activity in root apoplast seven days post-inoculation
[30][72].
Table 1. Schematic overview of the enzymatic antioxidants.
| ENZYMATIC ANTIOXIDANTS |
| Enzyme |
Chemical Reaction |
Involved in |
Cellular Location |
Ref. |
Schematic overview of the non-enzymatic antioxidants.
| NON-ENZYMATIC ANTIOXIDANTS |
| Enzyme |
Functions |
Location |
Ref. |
| Superoxide Dismutase (SOD) |
O2•−+O2•−+2H+→2H2O2+O2 |
Regulation of oxidative stress.
Stress resistance or tolerance mechanisms |
Apoplast, cytosol, mitochondria, chloroplast, peroxisomes |
| Ascorbic Acid (AsA) |
-Stress perception
-Redox homeostasis
-Regulation of oxidative stress
-Improvement of plant stress tolerance |
Apoplast, cytosol mitochondria, chloroplast, vacuoles, peroxisomes, nucleus |
[31][41][23,81] | [31][27][23,69] |
| Catalase (CAT) |
H2O2→H2O+(1/2) O2 |
Regulation of oxidative stress.
Stress resistance or tolerance mechanisms.
Plant metabolism |
Apoplast, cytosol, chloroplast, mitochondria, peroxisomes |
| Glutathione (GSH) |
-Protect membranes.
-Prevent protein oxidative denaturation under stress conditions
-Substrate for glutathione peroxidase and gluthatione S-transferase
-Metal chelator | [ | 31 | ][32][33] |
Apoplast, cytosol, chloroplast, mitochondria, vacuole, peroxisome, nucleus[23,27,73] |
| [ | 41 | ] | [81] |
Ascorbate Peroxidase (APX) |
H2O2+Asc→2H2O+DHA |
Regulation of oxidative stress.
Stress resistance or tolerance mechanisms |
| Proline (Pro) |
-Osmoprotectant activity
-Antioxidant capacity
-Metal chelator
-Signalling under abiotic and biotic stresses
|
Plant growth and physiology |
Apoplast, cytosol, mitochondria, peroxisomes, chloroplast |
-Plant growth and development[34 |
Apoplast, cytosol, mitochondria, chloroplast |
[42][43]][35][74,75] |
| [ | 82 | , | 83] |
Monodehydroascorbate Reductase (MDHAR) |
MDHA+NADPH→Asc+NADP+ |
| Phenolic Compounds |
-Antioxidant activity
-Metal chelator
-Protective and signalling functions against different stresses | Regulation of oxidative stress.
Stress resistance or tolerance mechanisms |
Apoplast, cytosol, mitochondria, chloroplast |
-Plant growth and development[ |
Ubiquitous |
[44][84]31][36][37][23,76,77] |
| Dehydroascorbate Reductase (DHAR) |
DHA+2GSH→Asc+GSSG |
| Polyamines |
-Antioxidant capacity
-Plant growth and development.
-Biotic and abiotic stress responses.
| Regulation of oxidative stress
Stress resistance or tolerance mechanisms
Plant growth and development |
Apoplast, cytoplasm, mitochondria, chloroplast, peroxisomes |
-Osmotic adjustment ability |
Ubiquitous |
[45][46][85,86][31][38][39][23,78,79] |
| Glutathione Reductase (GR) |
GSSG+NADPH→2GSH+NADP+ |
Regulation of oxidative stress.
Stress resistance or tolerance mechanisms |
Apoplast, cytoplasm, mitochondria, chloroplast |
[40][80] |
Catalases (CAT) are the only and the main scavenging enzymes that do not require any reductant for ROS decomposition and directly decompose H
2O
2 to water and O
2 [47][33]. Moreover, it seems that there is a link between CAT activity and ROS production by RBOH and POXsin the apoplast
[48][87]. In addition, CAT seems to have the highest enzyme activity in this intercellular space, reaching around 0.2% to 2% of the total
[28][29][49][70,71,88]. Several studies have shown that an increase of CAT is essential for plant defense against different stresses, since its correct functioning could avoid the damages produced by ROS accumulation. For example, the overexpression of a catalase gene isolated from
Cucumis sativum (
CsCAT3) in
E. coli could increase the tolerance to cold, heat, osmotic, or salinity stresses
[50][89]. Moreover, it was also suggested that SA binds this enzyme, reducing its activity such as it occurs in NO
3− treated citrus plants exposed to salinity compared to NH
4+ treated plants that showed an increase in CAT activity, allowing plants to tolerate better the salinity-induced oxidative damage
[51][90].
Within the antioxidant system in plants, the ascorbate–glutathione (Asc-GSH) pathway plays a major role in regulating ROS. Four enzymes are engaged in this pathway:
ascorbate peroxidase (APX), dehydroascorbate reductase (DHAR), monodehydroascorbate reductase (MDHAR), and glutathione reductase (GR). These enzymes catalyze the reduction of H
2O
2 to water using ascorbate (Asc) and glutathione (GSH) as electron donors. In the Asc-GSH pathway the reduction of H
2O
2 is ultimately linked to NAD(P)H oxidation. All the enzymes of the Asc-GSH pathway were found in the apoplasts of barley, oat, and pea leaves, but their activity was much lower when compared to their symplastic activity
[28][29][70,71]. However, in the apoplasts of other plants, not all Asc-GSH pathway enzymes were found to be active; therefore, only certain enzymatic reactions may take place
[52][91].
Ascorbate peroxidase (APX) is a key enzyme of the Asc-GSH pathway
[53][92]. APX catalyzes the reduction of H
2O
2 to water using Asc as an electron donor. Based on amino acid composition, five isoforms of APX have been identified in plants: cytosolic (cAPX), mitochondrial (mitAPX), chloroplastic (chlAPX: stromal-APX and thylakoidal-APX), and peroxisomal/glyoxysomal (mAPX)
[54][93]. In Arabidopsis only the cytosolic isoform was found to be secreted to the apoplast
[55][94]. It was observed that the expression of APX encoding genes takes place under abiotic stresses (salt, drought, heat, cold, UV radiations, oxidative stress, etc.) and under biotic stresses (pathogen attack, herbivory). The role of APX isoforms concerning different types of abiotic stresses is reviewed in detail by Pandey et al.
[54][93]. An increase in APX in the leaf apoplast of
Ctenanthe setosa in response to drought stress was observed
[56][95]. Related to biotic stress, in the Plum pox virus-resistant apricot cultivar Stark Early Orange, a rise in class I APX and strong increases in POX and SOD activities were noticed in the apoplastic compartment
[57][96].
3. Apoplastic Proteins and Peptides in Modulating Plant–Pathogen Interactions. Microbe-Associated Molecular Patterns (MAMPs) of Proteic Nature
The most abundant proteins related to defense are the pathogen-related proteins (PR-proteins) that represent 23–33% of the total apoplastic fluid proteins (AFPs)
[58][115]. The production and accumulation of PR-proteins are produced by various stresses, including biotic/biological stresses and abiotic/non-biological stresses
[59][116]. The plant PR-proteins are divided into 17 groups (). Moreover, based on the amino acid sequence and the biochemical activity identified during the last decade, they also include novel peptide families. The PR peptides include proteinase inhibitors (PR-6 family), plant defensins (PR-12 family), thionins (PR-13 family), and lipid transfer proteins (PR-14 family)
[60][61][117,118].
Table 3. Classification of pathogenesis-related proteins and peptides.
| Families |
Properties |
References |
| PR-1 |
Antifungal |
[62][119] |
| PR-2 |
β-1,3-glucanase |
[63][120] |
| PR-3 |
Chitinase type I, II, IV, V, VI, VII |
[64][121] |
| PR-4 |
Chitinase type I, II |
[65][122] |
| PR-5 |
Thaumatin- like |
[66][39] |
| PR-6 |
Proteinase- inhibitor |
[67][123] |
| PR-7 |
Endoproteinase |
[68][124] |
| PR-8 |
Chitinase type III |
[69][125] |
| PR-9 |
Peroxidase |
[70][126] |
| PR-10 |
Ribonuclease like |
[71][72][127,128] |
| PR-11 |
Chitinase, type I |
[73][129] |
| PR-12 |
Defensin |
[74][130] |
| PR-13 |
Thionin |
[75][131] |
| PR-14 |
Lipid- transfer protein |
[76][132] |
| PR-15 |
Oxalate oxidase |
[77][133] |
| PR-16 |
Oxalate oxidase-like |
[67][123] |
| PR-17 |
PRp27 Unknown |
[78][134] |
Besides the plant PR-peptides mentioned above, the regulation of signaling pathways by small peptides is a central theme in molecular biology. The apoplast is also the space where the microbe-associated molecular patterns (MAMPs), by plant cell surface-localized pattern recognition receptors (PRRS), are initially determined
[79][135].
3.1. Apoplastic Proteins Related to Plant Defense
Nowadays, PR-proteins, based on their protein sequences similarities and other biological features, show diverse functions, such as β-1,3-glucanase (PR-2), chitinases (PR-3), thaumatin-like (PR-5), and peroxidases (PR-9)
[61][80][118,136].
Despite their differential fungicidal activity, PR-1 proteins are often used as markers for salicylic acid (SA)-mediated disease resistance
[58][62][115,119]. PR-1 proteins have been identified in
Oryza sativa,
Nicotiana tabacum, and Arabidopsis, and often have a molecular weight of 14 to 17 kDa
[81][137]. Although PR-1 proteins can have different properties and differ substantially in biological activity, all of them have a similar structure and are classified in the same family based on sequence homology
[66][39].
Plant β-1,3-glucanases are grouped in the PR-2 family of PR proteins and play an imperative role in plant defense, and the usual processes of plant growth. These proteins have a size from 33 to 44 kDa, with both acidic and basic isoforms
[63][120]. Based on the amino acid sequencing, structural features, and cellular localizations, the glucanases have been classified into three major classes and two minor classes
[82][83][84][138,139,140]. Most of classes II, III, and IV of β-1,3-glucanases acidic proteins were found and secreted into the extracellular spaces. Moreover, several studies have shown that the synthesis of β-1,3-glucanases is stimulated by pathogen infections and it makes the plant resistant to fungal pathogens either alone or in association with other proteins, including chitinases, peroxidases, and thaumatin-like proteins
[85][86][87][141,142,143]. Furthermore, it has been suggested that β-1,3-glucanases act as the key enzymes in the lysis of phytopathogenic fungal cell walls during the antagonistic action by hydrolyzing the O-glycosidic linkages of β-glucan chains
[88][89][144,145]. Besides, the results shown by Floerl et al.
[10] demonstrated PR-2 apoplastic activity in susceptible primary leaves of
Arabidopsis thaliana 25 days after inoculation with
Verticillum longisporum. Additionally, in the related article they reported that β-1, 3-glucanases are increased in wheat plants (
Triticum aestivum) in response to
Zymoseptoria tritici infection
[65][122].
Chitinases are a huge and diverse group of enzymes that have been classified into two main categories—endochitinases and exochitinases, and based on their primary structures, plant chitinases have been divided into seven classes, class I through VII
[90][91][146,147]. Plant chitinases which represent a major group of PR-proteins are expressed in response to environmental stresses and the attack of phytopathogens and secreted in extracellular space
[92][93][94][95][96][148,149,150,151,152]. Moreover, several studies have shown that extracellular chitinases induce chitinase biosynthesis by detecting fungal elicitors and block the spreading hyphae invading intercellular spaces as well. For example, Petriccione, et al.
[97][153] reported that extracellular chitinase was induced in infected
Actinidia deliciosa leaves with
Pseudomonas syringae. Furthermore, Pechanova et al.
[98][154] showed that class IV chitinase and acidic class III chitinase were accumulated in leaves of poplar upon water stress. They demonstrated that three different type III chitinases were detected in leaf apoplasts, while the poplar stem apoplast contains only one single type III chitinase. In addition, apoplastic chitinases are involved in the destruction of the bacterial cell wall, and in the production of ROS by the production of chitin oligomers
[99][155]. Chitinase induction has been reported against bacterial (mainly Pseudomonas) and fungal pathogens in
Actinidia deliciosa and
Brassica napus subsp.
Napus, respectively
[97][100][153,156]. Additionally, leaf apoplastic chitinases and 1, 3-β-glucanases increased in response to
Cladosporium fulvum and
Septoria tritici diseases
[101][102][157,158]. Moreover, Han et al.
[103][159] described the apoplastic Cys-rich repeat protein 1 (CRR1), a protector of chitinase, helping the plant resistance to the pathogen releasing VdSSEP1 protease.
On the other hand, thaumatin-like proteins (TLPs) are classified as the PR-5 protein family
[66][39]. Several studies have shown that plant TLPs are secreted in the apoplast and are involved in plant protection against biotic and abiotic stresses
[104][105][106][107][160,161,162,163]. Furthermore, various studies have demonstrated that TLPs exhibit antifungal activity. For example, Shatters Jr. et al.
[108][164] revealed that PR-5 proteins can affect fungi, either by disrupting its membranes or by hydrolyzing β-1,3-glucans of the cell walls. Additionally, Wang et al.
[107][163] have shown that the expression of
TLP genes in plants infected by fungi and bacteria is significantly increased.
Germin–like proteins (GLPs) are a large gene family that belongs to the cupin superfamily
[109][165]. GLPs are involved not only in plant development but also in plant defense against biotic and abiotic stresses, including bacteria
[110][166], fungi
[111][112][167,168], salt pressures
[113][169], and drought stresses
[114][170]. Moreover, ADP–glucose pyrophosphatase or phosphodiesterase (AGPPas), oxalate oxidase (OxO), and superoxide dismutase (SOD) are three enzymes which are associated with GLPs
[115][171]. In general, the primary reactions detectable in plant–pathogen recognition are the opening of specific ion channels and the formation of reactive oxygen intermediates
[116][172]. It seems that SODs in association with GLPs protect the plant from the effect of oxidative stress by quickly converting O
2− and H
2O into H
2O
2 and O
2 [117][118][173,174]. Furthermore, GLPs also seem to be related to phytohormones, since the transcript levels of some GLPs are enhanced after application of phytohormones, including SA and ABA
[119][175]. Additionally, a GLP with serine protease inhibitory activity (GLP-serine protease inhibitor) has been detected in the wheat apoplast and suggested to be part of a defense system against insect and bacterial proteases
[101][157]. Moreover, GLPs are also expressed in abiotic stress. For example, expression of GLPs in tomato and
Nicotiana tabacum plants in response to salt stress has been reported by Amini et al.
[120][176], and Dani et al.
[113][169], respectively.
The regulation of secreted proteases and protease inhibitors (PIs) plays an important role in apoplastic immunity
[121][177]. It seems that proteases increase after pathogen infection systemically and locally
[122][178]. Specifically, subtilisin-like serine proteases or subtilases play an essential role in responses to environmental stress
[123][179]. For example, the apoplastic PR protease (P69B) is required for the induction of tomato immunity against the pathogen
Phytophthora infestans [124][180]. Moreover, it has been also described that the Arabidopsis SBT3.3 gene, encoding an extracellular subtilase homologous to the tomato P69C, also plays a role in immune priming by induction SA-responsive gene
[125][181].
3.2. Apoplastic Peptides Related to Plant Defense
PR gene families PR-6, PR- 12, PR-13, and PR-14, which contain protease inhibitors, defensins, thionins, and lipid transfer proteins respectively, are the so-called antimicrobial peptides (AMPs). These AMPs are usually cysteine-rich molecules that possess potential and a broad range of antimicrobial activities
[61][76][118,132]. Specifically, the peptides belonging to the PR-6 family have shown effective antimicrobial activity against fungal pathogens
[126][182]. Apoplastic papain-like cysteine proteases (PLCP) play an important role in apoplastic immunity by releasing small peptides that serve as damage-associated molecular patterns (DAMPs)
[127][183]. In addition, it is worth noting that PLCPs and serine hydrolases (SHs) protease families are up-regulated upon pathogen infection or targeted by pathogen-derived inhibitors
[128][184]. For example, Ziemann et al.
[129][185] pointed out that Zip1 peptide, which is released in the presence of PLCP, leads to SA accumulation in maize leaves. Moreover, Schulze Hüynck et al.
[130][186] identified a specific PLCP named CP1C in maize root. Surprisingly, CP1C which shares sequence homology to the Arabidopsis RD21 subfamily did not show higher activity after SA treatment of maize roots. Moreover, RCR3 is an apoplastic PLCP that acts as PR protein against fungal pathogens and nematodes
[131][132][187,188].
Plant defensins and thionins are small, cysteine-rich peptides (around 5 kDa) which are grouped into plant-specific antimicrobial peptides
[133][134][189,190]. There is strong evidence that some defensins are also induced upon pathogen attack, environmental stimuli, and jasmonate treatment
[74][135][130,191]. For example, the defensin gene
MtDef4.2 of
Medicago truncatula plants provides strong resistance to the fungus
Puccinia triticina in transgenic wheat
[136][192]. Additionally, the
Arabidopsis thaliana apoplastic defensin type 1.1 (
AtPdF1.1) acts to mediate defense against
Pectobacterium carotovorum subsp.
Carotovorum by activation of ethylene pathway in result of iron-deficiency
[137][193]. Moreover, an increasing level of Tad1 which is a plant defensin-like has been detected in wheat in response to pathogen attack and cold temperatures. Interestingly, the
Tad1 expression is an independent defense signal and it is not regulated by the methyl jasmonate acid pathway
[138][194].
Lipid transfer proteins (LTPs) are small, cationic, cysteine-rich peptides, which are recognized based on their role and ability to transport lipids between cell membranes. They usually have below 10 kDa molecular weights
[139][140][195,196]. Extracellular LTPs have been found in various plant species, such as tobacco, cowpea (
Vigna unguiculata),
Medicago, and sunflower
[141][142][143][197,198,199]. Researchers have been interested in LTPs for three main reasons: for their ability of transferring and binding to lipids, because they are one of the components of plant innate immunity, and for their clinical features
[144][200]. Apoplastic LTPs were shown to be involved as major players in cutin formation
[145][201]. However, much evidence showed that LTPs play a key role in plant defense mechanisms
[146][147][202,203]. For example, Dani et al.
[113][169] detected two LTPs in the leaf apoplasts of tobacco in response to salt stress. It seems that the induction of LTPs during this, stress causes deposition of cuticular material decreasing, and that way, leaf transpiration. Moreover, defective-mutants in the induced resistance gene
DIR1, which encodes apoplastic LTPs, can still promote the SA-mediated pathway, but they have disabled the induction of systemic acquired resistance (SAR), which is one of the resistance mechanisms in plants
[147][148][203,204].
3.3. Microbe-Associated Molecular Patterns of Proteic Nature
The recognition of microbe-associated molecular patterns (MAMPs) triggers several cellular response processes, such as alteration of ionic current between cell membranes which alkalizes intercellular space, increasing the cytoplasmic calcium ion concentration
[128][184], and the biosynthesis of ethylene stress hormone which is activated by 1-aminocyclopropane-1-carboxylic acid-synthase (ACC-synthase). Other primary responses include activation of mitogen-activated protein kinases (MAPKs), which in turn activate transcription factors
[149][205]. This is followed by transcription of protein-coding genes such as defensins and other antimicrobial metabolites
[150][206].
Demonstrating the action of bacterial flagellin (a type of protein subunit of the bacterial flagellum) in the plant was the turning point in understanding innate immunity
[151][207]. Previous studies have shown that flagellin, or more precisely flg22, inhibits the growth of Arabidopsis and leads to callose deposition in leaf tissue
[152][208]. Flg22 led to alkalization of the tobacco cell culture suspension, and stimulated the production of ROS in the tobacco leaves. The receptors responsible for flagellin perception by Arabidopsis are the leucine-rich repeat (LRR) transmembrane receptor kinase flagellin sensitive 2 (FLS2) and the co-receptor brassinosteroid insensitive 1-associated kinase 1 (BAK1)
[153][154][209,210]. It was determined that the message sent by FLS2 was through a cascade of MAPK, including AtMPK3 and AtMPK6, and led to an induction of the expression of
WRKY22 and
WRKY29 genes
[149][205]. Moreover, when plants were treated with flg22 it protected them against a subsequent pathogen challenge, providing direct evidence that it drives an effective immune response in the plant. In addition, it seems that the flagellin perception on leaf surface may be important in stimulating the active defense response because the
Arabidopsis fls2 mutants, which had lost the flagellin receptor, were more susceptible to
Pseudomonas syringae pv DC3000 infection
[155][211].
The elongation factor thermal unstable, abbreviated as Ef-Tu, is another widely described MAMP protein which plays an unavoidable role in protein biosynthesis, so its presence is inevitable for bacteria. The protein was isolated and identified from
E. coli, which had been shown to have severe MAMP activity even by disrupting its flagellin gene
[156][212]. The receptor of this protein, called elongation factor receptor (EFR), like FLS2 also belongs to the LRR family
[157][213].
Moreover, many plant species of the Solanaceae family detect the highly conserved nucleic acid binding motif RNP-1 of bacterial cold-shock proteins (CSPs), represented by the peptide csp22, as a MAMP. The recipient of this MAMP, which is a kind of plant receptor-like kinase (RLK), was identified in tomato and named CORE. Transfer and expression of cold shock protein receptor to
Arabidopsis thaliana can increase plant resistance to
P. syringae pv. tomato
[158][214].
Unlike flagellin, EF-Tu, and CSP, the fungal protein ethylene-inducing xylanase (EIX) activity cannot be attributed to a peptide. Fungal xylanase—by releasing cell wall fragment, and therefore, the production of DAMPs—can activate various defense responses such as alkalization of the extracellular space, ethylene biosynthesis, and the production of ROS in tomato and tobacco
[159][160][161][215,216,217]. EIX is recognized by the tomato leucine-rich repeat receptor-like proteins with a signal for receptor-mediated endocytosis, LeEIX1, and LeEIX2, of which only the latter mediates a hypersensitive response
[162][218].
Besides proteic MAMPs, structural components of bacterial cell wall such as lipopolysaccharides (LPS) and peptidoglycan (PGN) can also act as MAMPs in the plant
[163][164][165][219,220,221].