Non-coding RNAs (ncRNAs) are involved in the regulation of cell metabolism and neoplastic transformation. Recent studies have tried to clarify the significance of these information carriers in the genesis and progression of various cancers and their use as biomarkers for the disease; possible targets for the inhibition of growth and invasion by the neoplastic cells have been suggested. The significance of ncRNAs in lung cancer, bladder cancer, kidney cancer, and melanoma has been amply investigated with important results. Recently, the role of long non-coding RNAs (lncRNAs) has also been included in cancer studies. Studies on the relation between endometrial cancer (EC) and ncRNAs, such as small ncRNAs or micro RNAs (miRNAs), transfer RNAs (tRNAs), ribosomal RNAs (rRNAs), antisense RNAs (asRNAs), small nuclear RNAs (snRNAs), Piwi-interacting RNAs (piRNAs), small nucleolar RNAs (snoRNAs), competing endogenous RNAs (ceRNAs), lncRNAs, and long intergenic ncRNAs (lincRNAs) have been published. The recent literature produced in the last three years was extracted from PubMed by two independent readers, which was then selected for the possible relation between ncRNAs, oncogenesis in general, and EC in particular.
- endometrial cancer
- epigenetics
- non-coding RNA (ncRNA)
- long non-coding RNA (lncRNA)
- small non-coding RNA (small ncRNA)
1. Introduction
The analysis of non-coding RNA (ncRNAs) expression in endometrial cancers (EC) might be
of importance in understanding the neoplastic transformation of the endometrium. These RNA
transcripts are possible biomarkers of early neoplastic transformation, but their significance might be
even greater.
Approximately only 2% of the genome in humans has protein encoding functions, so most of
the human genome is not translated into proteins. It has been called genomic dark matter, because no
specific function has been described for it [1]. However, many information-carrying genes have been
demonstrated to have significant regulatory functions, even if they remain untranslated. The ncRNAs
include transfer RNAs (tRNAs), ribosomal RNAs (rRNAs), antisense RNAs (asRNA), micro RNAs
(miRNAs), small nuclear RNAs (snRNAs), small nucleolar RNAs (snoRNAs), competing endogenous
RNAs (ceRNAs) and piwi-interacting RNAs (piRNAs).
Recently, an additional group of ncRNAs has been discovered: they are called long-ncRNAs
(lncRNAs), as they are over 200 nucleotides in length, in contrast to other ncRNAs, which are
usually shorter [2,3]. lncRNAs are less comparable between species; protein-coding genes tend
to be homologous between mammals [4]. Four main mechanisms are involved in their expression
and they are sometimes defined as archetypes: signaling, decoying, scaffolding and guidance [5,6].
lncRNAs seem to have an importance in physiological regulation, and their dysfunction has been
implicated in cardiovascular and neurodegenerative diseases as well as in cancers [7,8]. lncRNAs
seem to be important modulators in biological mechanisms and significant cofactors of tumorogenesis.
lncRNA gene polymorphisms have been linked to cancer risk [9]. Systematic reviews on the association
between lncRNA single-nucleotide polymorphisms (SNPs) and the overall risk of developing a cancer
are starting to appear in the literature. A total of 17 SNPs in four common lncRNA genes have
already been described by Zhi et al. in a meta-analysis [9]. However, the involvement of lncRNAs
in endometrioid endometrial adenocarcinoma (EEC) remains to be fully addressed. Indeed, RNAs
are expressed in different ways in the normal, hyperplastic and dysplastic endometrium [6]; hence,
interference in RNA expression could actually lead to the clinical control of neoplastic transformation
and disease progression. We offer an overview of the current knowledge of the relationship between
ncRNAs and endometrial adenocarcinoma (EC) that shows promise in the diagnosis and treatment of
this condition.
Long Non-Coding RNAs: General Introduction
lncRNAs cover a large part of the non-coding information of the human DNA, which represents
more than 90% of the whole genome. They constitute a wide and complex group of molecules with
more than 200 nucleotides, usually lack an open reading frame, and in various ways are involved in
the pathophysiology of cancer [2,3,5]. Their roles in the regulation of gene expression, imprinting,
transcription, and post-translational processing have been documented in several types of cancer.
Thousands of long intergenic ncRNAs (lincRNAs) have been identified in various mammals in genome
sequencing studies. Some of these RNAs have been conserved during the evolution of various species
and might have a fundamental importance in the regulation of cell mechanisms [2–5].
3. Competing Endogenous RNAs, miRNA, lncRNA Profiling Regulation and Cancer
The miRNAs are a class of small ncRNAs that have been associated with several diseases,
including cancer [21]. They bind to target mRNAs via sequence complementarity, leading to translation
inhibition and mRNA destabilization. Several lncRNAs are capable of binding to miRNAs interfering
with this mechanism, thus blocking their action on target mRNAs. These lncRNAs are called
ceRNAs, and several examples having a role in tumorigenesis and evolution have been published.
Many lncRNA–miRNA interactions have been reported or suggested recently [22]. For example,
the lncRNA named MEG3 was recently implicated as an influencer of the signal transducer and
activator of transcription 3 (STAT3) expression, by altering miR-21 expression in ovarian cancer; it
has been shown to function as a tumor suppressor in many cancer types [23]. Maternally expressed
gene 3 (MEG3) is a maternally imprinted lncRNA with a tumor suppressor role in various tumors.
Sun et al. [24] found significantly lower MEG3 expression in endometrial carcinoma tissues than in
normal endometrial tissues.
A cross downregulation of miRNA and lncRNAs, regulating the metastasis associated lung
adenocarcinoma transcript 1 (MALAT-1) was suggested by Li et al. [25]. According to the authors,
the miR-200 family members were present in a high concentration in EEC, melanoma, and some
ovarian cancers. They showed that miR-200c levels were higher in most EEC specimens than in healthy
tissues, while MALAT-1 levels were much lower. However, they also found that miR-200c was directly
bound to MALAT-1 and miR-200c, both of which showed cross-repression, while TGF- β increased
MALAT-1 expression, working as an inhibitor of miR-200c. The interruption of the repressive effect of
miR-200c over MALAT-1 decreased the invasive capacity of EEC cells; the epithelial to mesenchymal
cell transition (EMT) markers’ expression was altered, at least in vitro [25]. However, the abnormal
expression of the lncRNA MALAT-1 or miR-200 family members were shown to facilitate EMT in
various human cancers. The details of the regulatory mechanism of MALAT1 and miR-200 remain
largely unknown.
lncRNAs can be a precursor for miRNAs. For example, H19 is the gene precursor of a ubiquitous
lncRNAs that show a negative or limiting regulation regarding body weight and cell proliferation. H19
is the RNA precursor of miR-675 [26]. Other miRNAs have been associated with tumor progression:
Vennin et al. showed that miR-675 enhanced tumor initiation, progression, and metastasis of breast
cancer cells by downregulating the c-Cbl and Cbl-b gene coding for the CBL protein [26].
This protein is an E3 ubiquitin-protein ligase involved in cell signaling [26]. The expression
pattern and biological functions of the nuclear-enriched abundant transcript 1 (NEAT1) was also
related to EEC. The levels of NEAT1 were up-regulated in ECC tissues and cell lines [27]. Moreover,
the lncRNA cancer susceptibility candidate 2 (CASC2) was identified as a potential tumor suppressor
in EC [28].
The ovarian adenocarcinoma amplified lncRNA (OVAL) is composed of three exons of
1489 nucleotides in length. OVAL was thought to encode various expressed sequence tags (ESTs) and
mRNAs, but recently an alternative first exon isoform has been identified, leading to the transcription
of the lncRNA OVAL [29]. The OVAL is over-expressed in Type I EC and in serous ovarian cancer
(OC). Type II EC, on the other hand, is four times less likely to show upregulation of the OVAL gene;
p53-regulated genes are upregulated in EC tissue samples with high expression of OVAL [29,30].
The carcinogenetic effect of estrogens in type I EC is well-documented [31]. Endometrial cancer
and breast cancer (BC) show a similar sensitivity to estrogens. According to Bhan et al., estradiol
induces lncRNA HOTAIR expression, as well as estrogen receptors and general transcription factors
of RNA polymerase II [32]. Thus, HOTAIR is not only abnormally expressed in BC, but in EC as
well [33]. According to He et al., HOTAIR upregulation was found in nearly three-quarters of EC
samples (63 out of 87), only 20% of control endometria tested positive for this lncRNA (4 out of 30).
Moreover, differentiated tumor samples (G3 grade) showed higher expression levels than Grade 1
samples. HOTAIR gene upregulation could be detected in about 50% of hyperplastic endometriums
(5 out of 12). HOTAIR is also associated with metastatic disease. High levels of HOTAIR expression
correlates with metastases and decreased patient survival in EC [34]. The lncRNA steroid receptor
RNA activator (SRA) regulates gene expression induced by steroid receptors [35]. SRA is upregulated
in BC and in other steroid-responsive tumor tissues, such as ovarian carcinoma [36]. In EC, high levels
of SRA expression are found with no relation to histological tumor grade. In reference tissue samples
of normal endometria, SRA expression is low and this may imply an early role in tumorigenesis [35].
Curiously, according to Lanz et al., SRA-transgenic mice did not develop any tumors, even with SRA
over-expression [37].
The expression of lncRNA and their role in defining EEC subgroups and their clinical
aggressiveness must be defined further. Xu et al. suggested that six lncRNAs may be the
main regulators of endometrial carcinogenesis [38]. Xu measured a total of 172 lncRNAs and
188 mRNAs and reported that they were expressed differently in type I EC and the normal control
samples [38]. Mathematical analysis with the gene ontology (GO) pathway analysis and the
lncRNA and mRNA co-expression networks indicated that six lncRNAs (KIAA0087, RP11-501O2,
FAM212B-AS1, LOC102723552, RP11-140I24 and RP11-600K151) could be the main regulators of
endometrial carcinogenesis [38].
Another study drew a parallel between EEC and liver cancer (LC) in relation to lncRNA expression.
Jiang et al. isolated 1931 expressed lncRNAs and gave them importance as to tumor initiation with
a mathematical integrative analysis [39]. A clustering of lncRNA expression helped define three groups
according to the authors: (i) basal-like, (ii) luminal-like and (iii) Catenin, β-1 (CTNNB1)-enriched
subgroups. The basal-like subgroup had a much higher number of tumors with higher pathological
grade (p < 0.0001), and TNM stage (p = 0.01); in the luminal-like subgroup, progesterone (PGR) and
estrogen receptor (ESR1) genes were not downregulated as in the EEC basal-like subgroup. The lncRNA
profile of the CTNNB1-enriched EEC subgroup was almost equal to that of the CTNNB1-enriched LC
subgroup [39].
Liu et al. showed that the lncRNA Taurine Upregulated Gene 1 (TUG1) was involved in EC
tumorigenesis due to the inhibition of miR-299 and miR-34a-5p [39]. The relationship between
lncRNA-TUG1 and EC was studied in 104 EC specimens, and corresponding control tissues.
lncRNA-TUG1 expression in cancer samples was higher than that in adjacent normal tissues.
The authors were able to show that lncRNA-TUG1 helps the evolution and progression of EC through
the inhibition of miR-299 and miR-34a-5p [40].
In a large study, 30,586 lncRNAs and 26,109 transcripts (fold change > 2.0) were found in 45
tested EC samples [40]. In particular, compared with normal tissues, 4010 lncRNA were upregulated,
and 3350 of them were downregulated. Among these lncRNAs, 3 were upregulated and 4 were
downregulated. Pathway analysis revealed that 24 pathways were correlated to the upregulated
transcripts [41]. Chen et al. showed that miR-93 was expressed at high levels in EC samples compared
to normal endometrial samples [42].
Zhai et al. looked into the expression profiles of lncRNAs and coding genes in three paired EC and
adjacent healthy tissues [43]. There was significant a difference in lncRNA and coding gene expression
between EC and their adjacent healthy non-tumor tissues; 53 lncRNAs (p value < 0.05) were differently
expressed in EC, compared with controls. ASLNC04080 was the most upregulated lncRNA in 22 out
of 24 samples and in HEC-1-B cell line [43]. Li et al. measured the levels of 26 lincRNAs in 176 pairs
of EC and adjacent healthy biopsy samples in two separate regional Chinese populations [44]. They
found that a lincRNA, LINC00672, was abnormally downregulated during the development of EC.
LINC00672 is a p53-targeting lincRNA, acting along with heterogeneous nuclear ribonucleoproteins as
a suppressive cofactor that reinforces p53-mediated suppression of the LIM and SH3 domain protein
(LASP)-1; this mechanism could possibly be associated with increased tumor aggressiveness [44].
Sun et al. looked into lncRNAs using support vector machine and random forest methods [45].
All expressed lncRNAs were ranked according to the standardized drop in probable prediction accuracy.
An odds analysis for predictive performance was performed, adding one lncRNA at a time starting
with the top two lncRNAs; of the ranked list, five lncRNAs were the optimal mix for diagnostic accuracy.
When choosing more than five lncRNAs, there was a downward trend in predictive performance.
The top five lncRNAs (FLJ27354, RP11-275I14.4, VIM-AS1, CTB-51J22.1 and RP11-229P13.20) were
selected as optimal predictive lncRNA biomarkers of uterine corpus EC (UCEC) progression. Only
one lncRNAs (FLJ27354) was clearly associated with disease progression, the other top four lncRNA
markers (RP11-275I14.4, VIM-AS1, CTB-51J22.1 and RP11-229P13.20) were silent. Using these top five
markers led to a high discriminatory performance in distinguishing advanced stages from early stages
with 78% prediction accuracy, 96.6% sensitivity and 76.6% specificity. These lncRNAs could have
a functional role in the progression of EC initiating, and in the promotion of important cancer-related
processes [45].
4. Regulation of Cell Growth by Long Non-Coding RNAs in Endometrioid Endometrial
Adenocarcinoma
lcnRNA H19 levels were significantly higher in EC tissues than in para-tumoral samples [45].
The expression pattern of H19 in EC tissues was addressed by quantitative polymerase chain reaction
(Q-PCR), and its function characterized in the EC cell line through the inhibition of expression with
small interfering RNAs (siRNAs) [45]. Lowering H19 levels did not affect the growth of HEC-1-B EC
cells, but it did suppress their capability to migrate and invade. Furthermore, H19 downregulation
decreased the transcription factor Snail, associated with E-cadherin downregulation, destabilization
of the adherent junction, and cellular polarization, and increased matrix metalloproteinase (MMP)
expression, with no effect on vimentin levels, indicating at least a partial reversion of EMT [46]. H19
was able to enhance EC aggressiveness by modulating the EMT process [47].
The lncRNA BANCR promotes EC cell replication and tumor invasion, by regulating Matrix
MetalloProteinase-1 (MMP1) and MMP2, and via extracellular signal-regulated kinase-1 (ERK)–MAPK
signaling pathways. BANCR was highly expressed in type 1 EC tissues, promoting EC cell proliferation,
migration, and invasion by activating the ERK–MAPK signaling pathway that regulates MMP2 and
MMP1 expressions [48]. For this reason, BANCR could become a prognostic marker and important
drug target in type 1 EC. MEG3 and two signaling molecules, Notch1 and Hes1, were detected in
both EC tissues and cell lines by RT-PCR and western blot analysis, all of them showing significant
expression. Lentiviral vectors with whole MEG3 transcripts or Short (or small) hairpin RNA (shRNA)
targeting MEG3 (shMEG3) was transinfected to evaluate cell proliferation. MEG3 dysregulation was
studied in xenograft models (established by subcutaneous implantation), and tumor growth was
compared. Important downregulation of MEG3 was observed in EC samples compared to controls,
while levels of Notch1 and Hes1 were found to be significantly upregulated. Cell proliferation was
greatly inhibited by MEG3 over-expression, while the opposite was observed in MEG3 knockout
cells. Interestingly enough, MEG3 changes could be reversed by Notch1 regulators. Moreover,
over-expression of MEG3 was strongly associated with repressed in vivo growth, along with Notch
signaling inhibition. Downregulated MEG3 was an important anti-proliferative factor in EC, by
repressing the Notch signaling pathway [49].
Cisplatin-resistant Ishikawa cells (a human EC cell line) showed high autophagy activity
compared with non-cisplatin-resistant parent Ishikawa cells. After lncRNA profiling, HOTAIR was
identified as the culprit: it was downregulated both in cisplatin-resistant Ishikawa cells and parental
Ishikawa cells treated with cisplatin. RNA interference on HOTAIR reduced the proliferation of
cisplatin-resistant Ishikawa cells and increased autophagy in cisplatin-resistant Ishikawa cells with or
without cisplatin treatment. Furthermore, beclin-1, multidrug resistance (MDR), and P-glycoprotein
(P-gp) expression was mediated by lncRNA HOTAIR. It was concluded that HOTAIR can regulate the
cisplatin resistance of human EC cells, regulating autophagy by influencing Beclin-1, MDR, and P-gp
expression [30].
The tumor suppressor candidate 7 (TUSC7) is an antisense lncRNA: if downregulated, it acts
as a possible tumor suppressor in several cancers. In the study by Shang et al., the low expression
of TUSC7 was associated with high pathological stages of EC, which revealed that TUSC7 might be
involved in the initiation and progression of EC. Moreover, the expression of TUSC7 in EC samples
and EC cell lines resistant to cisplatin CDDP, and Taxol was lower than that in sensitive EC tissues and
cell lines, which indicated that the TUSC7 expression level was positively related to the response of EC
patients to chemotherapy with CDDP and Taxol [50].
Li et al. showed that the expression and function of NEAT1 in EEC was elevated in ECC samples
and cell lines, with a high expression levels of NEAT1 being associated with enhanced cell growth,
colony formation ability, and invasive and migratory ability in pGCMV-NEAT1, while limiting the
expression of NEAT1 in HEC-59 cells by siNEAT1 transfection showed the opposite effects [27].
Guo et al. demonstrated that the growth arrest-specific 5 (GAS5) functions as a significant tumor
suppressing lncRNA in EC. Inhibiting the expression of miR-103, GAS5 enhances the expression of
the PTEN gene leading to cancer cell apoptosis. GAS5 downregulation could represent an important
factor in the pathogenesis of EC [51].
Moreover, the lncRNA Fer-1-like protein 4 (FER1L4) might suppress EC cell proliferation.
A plasmid containing FER1L4 was transfected into HEC-50 cells, showing low levels of FER1L
expression. Western blot analysis was used to determine PTEN expression and Akt phosphorylation.
FER1L4 was significantly downregulated in EC tissues compared to controls. This was positively
correlated with decreased PTEN expression. Therefore, FER1L4 could promote PTEN expression
and inhibit Akt phosphorylation. A significant decrease of cell proliferation was observed in FER1L4
overexpressing cells, with cell cycle arrest at G0/G1 phase and enhanced apoptosis [52].
5. Long Intergenic Non-Protein Coding RNA, Regulator of Reprogramming and Endometrioid
Endometrial Adenocarcinoma
Ample evidence indicates that linc- Regulator of Reprogramming (ROR) has a significant role in
tumor initiation and progression. In tumorigenesis, linc-ROR might act as an oncogene [53]. In general,
linc-ROR can be easily associated with cell proliferation, differentiation, apoptosis, invasion, and
metastasis in many human cancers. However, the molecules that act as mechanisms that mediate
linc-ROR’s effects must still be clearly identified before linc-ROR can be used in tumor treatment. Many
studies have shown the importance of linc-ROR as a tumor marker. Marked upregulation of linc-ROR
can be observed in various cancers, like BC [54,55], pancreatic cancer (PC) [56,57], hepatocellular
cancer (LC) [58,59], EC [60,61], and nasopharyngeal carcinoma [62]; furthermore, linc-ROR works as
a tumor suppressor in glioma [53]. Although linc-ROR is over-expressed in various cancers, the target
genes are variable and depend on the cell type.
In the paper by Chen et al., RNA was isolated from malignant and adjacent non-affected
endometrial tissue from six patients with low-grade, type I EC. Subsequently, Illumina paired-end RNA
sequencing was performed, to determine differential transcriptome expression patterns. Linc RNAs
were specifically analyzed. LINC00958 was upregulated, and four lincRNAs including LINC01480,
LINC00645, LINC00891 and LINC00702 demonstrated exquisite specificity for malignant endometrium
compared to normal endometrium, while also distinguishing EC from OC and Cervical Cancers
(CC) [62].
Zhou et al. studied a linc-RoR which was possibly implicated in the expression regulation
of core stem cell transcription factors (TFs) as ceRNA [60]. Adenovirus vectors carrying green
fluorescent protein (GFP) were transfected into E-Twenty-six (ETs) TFs to evaluate the effect of
down- or upregulation of miR-145, linc-RoR or Dicer. Expression of linc-RoR and core stem cell
TFs was associated with cell differentiation of the ET TF, whereas miR-145 expression increased
after ET differentiation. A higher expression of miR-145 successfully led to the downregulation of
linc-RoR and core TFs. The opposite was achieved by knocking down miR-145 expression. The effects
of miR-145 could be nullified by increasing the expression of linc-RoR in ETs or mutated targeted
sequences in linc-RoR. Reduced Dicer expression were able to enhance the expression of linc-RoR
and core TFs. The linc-RoR acted as a miR-145 inhibitor on the differentiation of ETs in endometrial
carcinogenesis [60].
6. Micro IRNA in Endometrioid Endometrial Adenocarcinoma
MicroRNA-93 is derived from a paralog (miR-106b-25) of the miR-17-92 cluster and has
been implicated in tumor initiation and progression of breast, colorectal, hepatic, lung, ovarian,
and pancreatic cancer.
The article by Chen et al. demonstrated miR-93 overexpression in EC tissue samples as compared
with normal endometrium. According to Chen et al., miR-93 overexpression promotes cell migration
and invasion, by downregulation of E-cadherin while N-cadherin expression in EC cells seems
significantly enhanced [42].
miRNA-205 is probably a key regulator of EC gene expression. Higher levels of miRNA-205
expression were measured in EC with invasion limited to less than half of the myometrium and
initial EC. A better survival rate was associated with higher levels of miRNA-205 using Kaplan–Maier
analysis (p = 0.034); miRNA-205 could thus be used as a prognostic marker [63]. Levels of miR-944
were analyzed in 68 cancerous and 20 normal endometrial samples: miR-944 was highly overexpressed
in EC tissues compared to normal controls. Furthermore, increased levels of miR-944 were observed
in EC cell lines. miR-944 expression was associated with the EC stages and pathology results of the
International Federation of Gynecology and Obstetrics (FIGO) [64].
Using small-RNA sequencing and microarrays, significant differences in Small Non-Coding
RNAs (sncRNA) expression patterns between normal, hyperplastic and neoplastic endometrium could
be easily identified. The authors propose a sncRNA signature (129 microRNAs, 2 of which were
previously unknown, 10 piRNAs and 3 snoRNAs) of neoplastic transformation. This new sncRNA
signature probably reflects events leading to EC development [65,66].
Micro RNA as a Marker and Promoter of Tumor Aggression in EEC
Can miRNA evaluation accurately predict the risk of lymph node extension in EC patients?
Eighty-six matched EC cases evenly distributed between lymph node-positive and lymph
node-negative cases, were analyzed in the study by Ahsen et al. [67]. A predictive matrix was
generated using genomic miRNA expression in EC node-positive patients. An independent set of 28
other tumor samples was similarly characterized and was used in a test cohort. A predictive signature
of miRNA expression was generated and used to predict the metastatic status of the independent
test cohort. Using 18 miRNAs, 100% accuracy was obtained with the control group. The EC group
showed 90% accuracy in the node-positive cases, and 80% in node-negative cases (false discovery rate,
FDR = 6.25%) [67]. The Dicer1 endoribonuclease plays a critical role in miRNA biogenesis. Bahubeshi
et al. showed that Dicer1 dysfunction leads to the enrichment of tumor stemness and tumor aggression
both in vitro and in vivo [68]. Loss of Dicer1-induced abnormal expression of the miRNA let-7 family,
which comprises well-known tumor suppressors, thus regulating stemness in EC cells [68].
Hu showed that silencing nc886, a ncRNA, leads to apoptosis of human EC cells in vitro.
In addition, tests results showed higher level of nc886 in the late phase of human EC tissue, slightly
more than in the early phase of EC but always much higher than in normal endometrial tissue. nc886
inhibition led to increased protein levels of phosphorylated PKR (p-PKR) and caspase-3, NF-kB and
vascular endothelial growth factor (VEGF) were on the opposite decreased. The rate of apoptosis
in the nc886 inhibition group was increased and cell proliferation was much slower as compared to
controls [69].
The inhibition of the enhancer of zeste homolog 2 (EZH2), (a histone-lysine N-methyltransferase
enzyme), and the reactivation of tumor suppressor miRNAs might lead to a functional cancer therapy
regime strategy. Ihira et al. observed that EZH2-suppressed miRNA let 7b and miR-361, two possible
tumor suppressors, with consequent EC cell proliferation and invasion, and negated their stem-cell-like
properties. In EC cells, EZH2 induced and worked together with the transcription factor Yin Yang
1 (YY1) to suppress miR-361, which in turn upregulated Twist, a target of miR-361. Treating EC
cells with GSK343, a targeted EZH2 inhibitor, had the same effect of a siRNA-mediated EZH2
knockdown, upregulating miR-361 and downregulating Twist expression. Combining GSK343 with
5-AZA-2’-deoxycytidine suppressed cell proliferation with an apparent synergic effect and blocked
invasion in vitro, lowering the tumor size and weight in EC cell grafted mice. Worse patient outcomes
could be predicted: a quantitative real-time PCR (Q-RT-PCR) analysis of 24 primary EC tissues showed
that lower let-7b and miR-361 levels were dire prognostic indicators. Results were confirmed on a large
patient data set from The Cancer Genome Atlas [70]. EZH2 might drive EC progression by regulating
miR-361–Twist signaling and indicate EZH2 inhibition as a possible anti-EC therapeutic strategy [68].
miRNA-93 overexpression increased cell migration and invasion, and downregulated E-cadherin
expression while increasing N-cadherin expression [42]. Chen et al. showed, by dual-luciferase reporter
assay, that miR-93 directly binds to t 30 untranslated region of Forkhead Box A1 (FOXA1), and that its
over-expression downregulated FOXA1 expression. On the other hand, miR-93 inhibitor transfection
upregulated FOXA1 expression at protein and mRNA levels. The conclusion was that miR-93 may
aid or initiate the process of EMT in EC cells by targeting FOXA1 [42]. An intriguing hypothesis by
Gong et al. was that the miRNA-194 downregulation (miRNA-194, is an inhibitor of EMT in several
cancers) is associated to a worse prognosis in human EC. Self-renewal factor Sox3 induces EMT at
gastrulation and is also involved in EMT in several cancers. Ectopic expression of microRNA-194 in EC
stem cells induced a MET by normalizing E-cadherin expression, decreasing vimentin expression, and
interfering with cell invasion in vitro. Moreover, overexpression of microRNA-194 inhibited EC stem
cell invasion or metastasis in vivo by the injection of adenovirus microRNA-194. This demonstrates
the new mechanism by which Sox3 contributes to EC stem cell invasion and suggests that repression
of Sox3 by miRNA-194 may have a therapeutic use in the suppression of EC metastasis. The cancer
stem cell marker CD133 might be the most significant surface marker of EC stem cell [69]. The effects
of miR-29b regulate via MAPK–ERK and PI3K–Akt signaling pathways the angiogenesis in EC by
targeting Vascular endothelial growth factor A (VEGFA). In the paper by Chen et al., EC tissues were
compared with the adjacent normal tissues and showed that miR-29b expression was downregulated,
while the mRNA and protein expression of VEGFA, ERK, Akt, mTOR and B-cell lymphoma 2 (Bcl-2)
were upregulated [71]. The left-right determination factor 2 (LEFTY2), considered a suppressor of
cell proliferation, tumor growth, and regulator of stem cell properties and embryonic differentiation,
seems to be a negative regulator of cancer cell programming.
LEFTY2 seems to downregulate MKi67 expression and focal adhesion kinase (FAK) activity, while
upregulating miR-200 and E-cadherin. It is thus a significant negative regulator of endometrial cell
growth and migration, probably under-expressed in EEC [72].
7. Conclusions
Abnormal expression of several sets of genes is probably the prime mover or a significant factor
in the progression of a cancer, and both mechanisms can be at work in sequence. Genetic alterations
include both mutational and epigenetic changes, such as methylation, acetylation, and phosphorylation
of nuclear chromatin [73–75]. Cellular genes are in charge of proliferation, apoptosis, and stem cell
differentiation; all of them can undergo epigenetic modifications in cancer [76,77]. The significance of
ncRNA is starting to be put under investigation, but more questions than answers are available for
now. Is ncRNA a possible marker of cancer risk? Can it be used as a biomarker for tumor diagnosis
and as an indicator of tumor progression or of a risk of tumor metastases? Can selective activation
or inhibition of RNA pathways help cure or control the aggressiveness of the disease? Are results
valid for all cancers? Do some of the promising preliminary results obtained in other conditions
apply to EC? Various information-carrying genes have shown important biological and regulatory
functions without expressing a translation of their information, such as classically the production of
proteins. In particular, lncRNAs show specific and altered expression patterns in EC compared to
normal endometrial tissue (Table 1, Figure 1). Some well-known factors associated with EEC show
an association with ncRNA expression. Levels of sex hormones are involved in the development of
EC. Hormones affect RNAs like lncRNAs SRA, H19 and HOTAIR, and their levels could play a role
in the carcinogenesis of EC. Estradiol is an important link between expression of these lncRNAs as it
correlates positively with upregulation of H19 and HOTAIR in EC [32,78]. Preliminary in vitro studies
showing the reduced growth of EEC cells indicate that a possible new therapeutic pathway for EEC is
possible. Study of the mechanisms of expression of RNA in EEC should be pursued, and their effect
on cancer growth clarified. lncRNAs could be useful clinical biomarkers of EEC for progression to
advanced stages. Hence, lncRNAs could be optimal biomarkers, useful to identify patients at high
risk for progression to advanced stage, and the profiling of lncRNAs could actually offer improved
diagnostic accuracy.
2. Long Non-Coding RNAs: General Introduction
3. Competing Endogenous RNAs, miRNA, lncRNA Profiling Regulation and Cancer
4. Regulation of Cell Growth by Long Non-Coding RNAs in Endometrioid Endometrial Adenocarcinoma
5. Long Intergenic Non-Protein Coding RNA, Regulator of Reprogramming and Endometrioid Endometrial Adenocarcinoma
6. Micro IRNA in Endometrioid Endometrial Adenocarcinoma
Micro RNA as a Marker and Promoter of Tumor Aggression in EEC
7. Conclusions
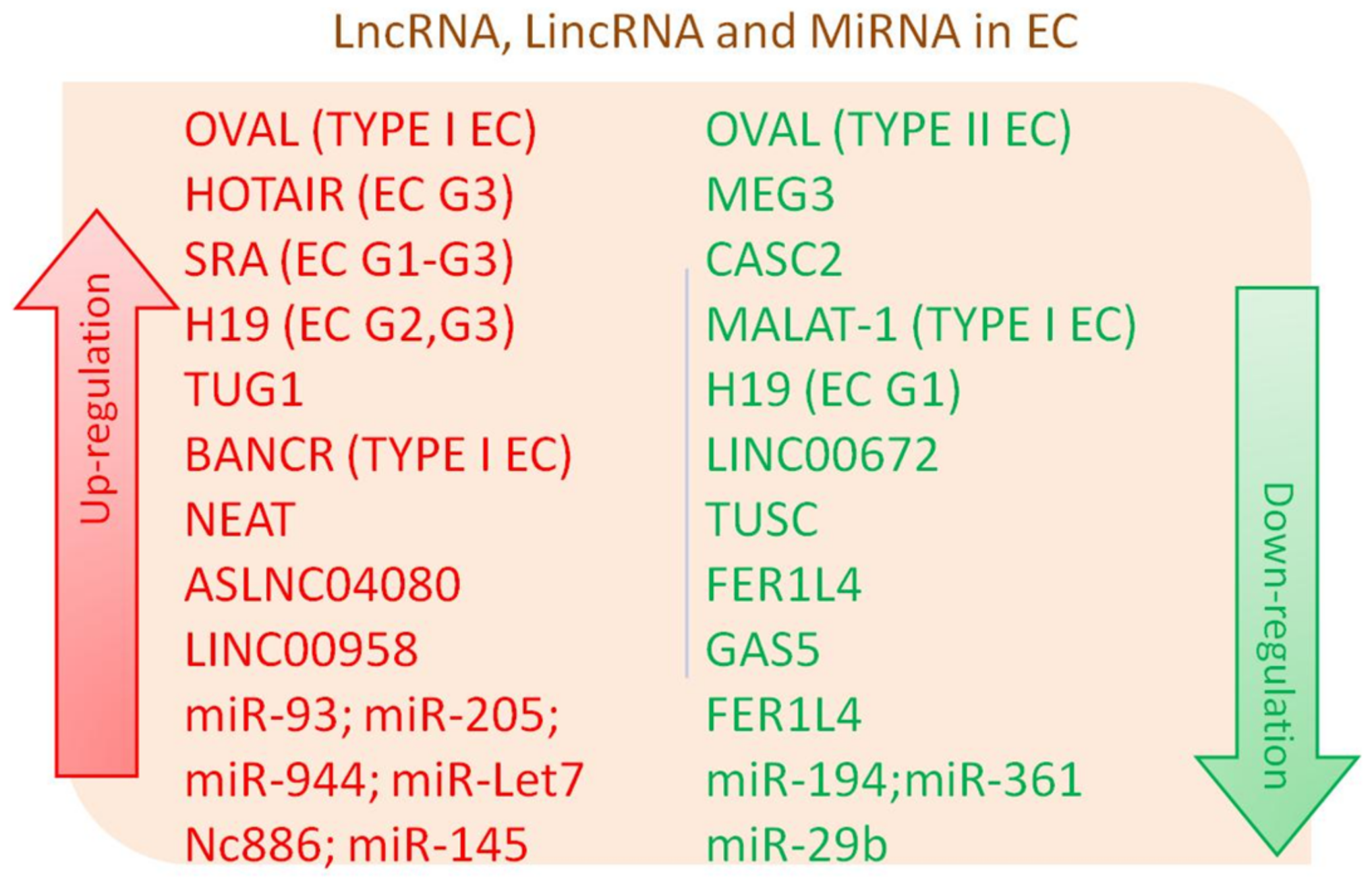 Figure 1. Differentially expressed long non-coding RNA (lncRNA), long intergenic non-coding RNAs (lincRNAs) lincRNA and micro RNAs (miRNA) in Endometrial Cancer (EC).
Figure 1. Differentially expressed long non-coding RNA (lncRNA), long intergenic non-coding RNAs (lincRNAs) lincRNA and micro RNAs (miRNA) in Endometrial Cancer (EC).| NcRNA | Type of ncRNA Regulation/Interaction | Type of EC Regulation/Pathway Interaction | In Vivo or In Vitro Assay or Humans | Reference |
|---|---|---|---|---|
| LncRNA OVAL | down and upregulation | downregulation in type II EC and upregulation in type I EC | In vitro assay from EC tissue | [6] |
| LncRNA MALAT-1 | downregulation/interaction with miR-200 | downregulation in EC through miR-200 | In vitro assay from EC tissue, xenograft tumor model | [14] |
| LncRNA TUG1 | interaction with miR-299/miR-34a-5p | promotion of EC via miR-299 and miR-34a-5p inhibition | In vivo mouse assay, in vitro assay from EC tissue and HEC-1-A cell lines | [29] |
| LncRNAs HOTAIR | upregulation | downregulation in Cisplatin-Resistant Ishikawa Cells (CRIC) and parental IC treated with cisplatin; up-regulation with EC tumor grade increase | In vitro assay CRIC cells; in vitro assay from EC tissue | [19][23] |
| LncRNAs H19 | upregulation | upregulation with EC tumor grade increase and other features associated with poor prognosis | In vitro assay from EC tissues and EC cell line | [35] |
| LncRNAs SRA | upregulation | upregulation regardless of histological tumor grade | In vitro assay from EC tissues | [25] |
| LncRNA BANCR | upregulation | promotion of EE cell proliferation and invasion by MMP2 and MMP1 regulation via ERK–MAPK signaling pathway in type 1 EC | In vitro assay from EC tissues | [37] |
| LncRNAs panel (e.g., KIAA0087, RP11-501O2, FAM212B-AS1, LOC102723552, RP11-140I24 and RP11-600K151) | differential regulation/interaction with 188 mRNAs | differential expression in type I EC | In vitro assay from EC tissues | [27] |
| LncRNA CASC2 | downregulation | potential role as a tumor suppressor | In vitro assay from EC tissues | [18] |
| LncRNA ASLNC04080 | upregulation | upregulation in 22–24 EC tissues and HEC-1-B cell line | In vitro assay from EC tissues and EC cell line | [32] |
| LncRNAs panel (e.g., FLJ27354, RP11-275I14.4, VIM-AS1, CTB-51J22.1 and RP11-229P13.20) | up and downregulation | potential biomarkers of Uterine Corpus (UCEC) | In vitro assay from EC tissues | [34] |
| LncRNA TUSC7 | downregulation | related to EC tumorigenesis and progression | In vitro assay from EC tissues | [39] |
| LncRNA FER1L4 | downregulation | decrease of cell proliferation in FER1L4-overexpressing cells | In vitro assay from EC cell line | [41] |
| LncRNA NEAT | upregulation | increase of Nuclear Enriched Abundant Transcript 1 (NEAT1) in EC | In vitro assay from EC tissues and cell lines | [16] |
| LncRNA MEG3 | downregulation | anti-proliferative role in EC by repressing Notch signaling pathway | In vitro assay from EC tissues and cell lines | [38] |
| LncRNA GAS5/miR-103 | downregulation / inhibition of miRNA-103 expression | tumor suppressor | In vitro assay from EC tissues and cell lines | [40] |
| LincRNA LINC00958 | upregulation | increase of LINC00958 in EC | In vitro assay from EC tissues | [51] |
| LincRNA LINC00672 | downregulation | downregulation during EC development | In vitro assay from EC tissues | [33] |
| NcRNA Nc886 | upregulation | upregulation of nc886 in EC late phases, compared to early stages and Normal Endometrial Tissues (NET) | In vitro assay from EC tissues | [57] |
| miRNA-93 | upregulation | over-expression associated to cell migration and invasion | In vitro assay from EC tissues and cell lines | [31] |
| miRNA-205 | upregulation | prognostic marker associated with better overall survival | In vitro assay from EC tissues | [52] |
| miRNA-944 | upregulation | over-expression in EC tissues compared to NET | In vitro assay from EC tissues | [54] |
| miRNA-29b | miRNA-29b/MAPK/ERK and PI3K/Akt signaling pathways | inhibition of angiogenesis by targeting VEGFA through MAPK/ERK and PI3K/Akt signaling | In vitro assay from EC tissues and cell lines | [62] |
| miRNA-145 | upregulation | upregulation of miR-145 lead to downregulation of linc-RoR7Dicer | In vitro assay from EC tissues | [49] |
| miRNA-361 | miR-361/twist signaling and miR-361/let-7b downregulation | lower let-7b, twist signaling and miR-361 are associated with worse patient outcome | In vitro assay from EC tissues | [60] |
| miRNA-194 | downregulation | downregulation associated to EC poor prognosis | In vitro assay from EC tissues and cell lines | [62] |
References
- Kapranov, P.; Cheng, J.; Dike, S.; Nix, D.A.; Duttagupta, R.; Willingham, A.T.; Stadler, P.F.; Hertel, J.; Hackermüller, J.; Hofacker, I.L.; et al. RNA maps reveal new RNA classes and a possible function for pervasive transcription. Science 2007, 316, 1484–1488.Vallone, C., Rigon, G., Gulia, C., Baffa, A., Votino, R., Morosetti, G., Zaami, S., Briganti, V., Catania, F., Gaffi, M., Nucciotti, R., Costantini, F. M., Piergentili, R., Putignani, L., Signore, F.; Non-Coding RNAs anNon-Coding RNAs and Endometrial Cancerd Endometrial Cancer. Genes 2018, 9, 187, 10.3390/genes9040187.
- Ma, L.; Bajic, V.B.; Zhang, Z. On the classification of long non-coding RNAs. RNA Biol. 2013, 10, 925–933.
- Ling, H.; Vincent, K.; Pichler, M.; Fodde, R.; Berindan-Neagoe, I.; Slack, F.J.; Calin, G.A. Junk DNA and the long non-coding RNA twist in cancer genetics. Oncogene 2015, 34, 5003–5011.
- Ponjavic, J.; Ponting, C.P.; Lunter, G. Functionality or transcriptional noise? Evidence for selection within long noncoding RNAs. Genome Res. 2007, 17, 556–565.
- Wang, K.C.; Chang, H.Y. Molecular mechanisms of long noncoding RNAs. Mol. Cell 2011, 43, 904–914.
- Smolle, M.A.; Bullock, M.D.; Ling, H.; Pichler, M.; Haybaeck, J. Long Non-Coding RNAs in Endometrial Carcinoma. Int. J. Mol. Sci. 2015, 16, 26463–26472.
- Mattick, J.S.; Amaral, P.P.; Dinger, M.E.; Mercer, T.R.; Mehler, M.F. RNA regulation of epigenetic processes. Bioessays 2009, 31, 51–59.
- Cheetham, S.W.; Gruhl, F.; Mattick, J.S.; Dinger, M.E. Long noncoding RNAs and the genetics of cancer. Br. J. Cancer 2013, 108, 2419–2425.
- Lv, Z.; Xu, Q.; Yuan, Y. A systematic review and meta-analysis of the association between long non-coding RNA polymorphisms and cancer risk. Rev. Mutat. Res. 2017, 771, 1–14.
- Pereira, D.M.; Rodrigues, P.M.; Borralho, P.M.; Rodrigues, C.M. Delivering the promise of miRNA cancer therapeutics. Drug Discov. Today 2013, 18, 282–289.
- McCaskill, J.; Praihirunkit, P.; Sharp, P.M.; Buck, A.H. RNA-mediated degradation of microRNAs: A widespread viral strategy? RNA Biol. 2015, 12, 579–585.
- Sheng, X.; Li, J.; Yang, L.; Chen, Z.; Zhao, Q.; Tan, L.; Zhou, Y.; Li, J. Promoter hypermethylation influences the suppressive role of maternally expressed 3, a long non-coding RNA, in the development of epithelial ovarian cancer. Oncol. Rep. 2014, 32, 277–285.
- Sun, K.X.; Wu, D.D.; Chen, S.; Zhao, Y.; Zong, Z.H. LncRNA MEG3 inhibit endometrial carcinoma tumorigenesis and progression through PI3K pathway. Apoptosis 2017, 22, 1543–1552.
- Li, Q.; Zhang, C.; Chen, R.; Xiong, H.; Qiu, F.; Liu, S.; Zhang, M.; Wang, F.; Wang, Y.; Zhou, X.; et al. Disrupting MALAT1/miR-200c sponge decreases invasion and migration in endometrioid endometrial carcinoma. Cancer Lett. 2016, 383, 28–40.
- Vennin, C.; Spruyt, N.; Dahmani, F.; Julien, S.; Bertucci, F.; Finetti, P.; Chassat, T.; Bourette, R.P.; Le Bourhis, X.; Adriaenssens, E. H19 non coding RNA-derived miR-675 enhances tumorigenesis and metastasis of breast cancer cells by downregulating c-Cbl and Cbl-b. Oncotarget 2015, 6, 29209–29223.
- Li, Z.; Wei, D.; Yang, C.; Sun, H.; Lu, T.; Kuang, D. Overexpression of long noncoding RNA NEAT1 promotes cell proliferation, invasion and migration in endometrial endometrioid adenocarcinoma. Biomed. Pharmacother. 2016, 84, 244–251.
- Palmieri, G.; Paliogiannis, P.; Sini, M.C.; Manca, A.; Palomba, G.; Doneddu, V.; Tanda, F.; Pascale, M.R.; Cossu, A. Long non-coding RNA CASC2 in human cancer. Crit. Rev. Oncol. Hematol. 2017, 111, 31–38.
- Cabili, M.N.; Trapnell, C.; Goff, L.; Koziol, M.; Tazon-Vega, B.; Regev, A.; Rinn, J.L. Integrative annotation of human large intergenic noncoding RNAs reveals global properties and specific subclasses. Genes Dev. 2011, 25, 1915–1927.
- Sun, M.Y.; Zhu, J.Y.; Zhang, C.Y.; Zhang, M.; Song, Y.N.; Rahman, K.; Zhang, L.J.; Zhang, H. Autophagy regulated by lncRNA HOTAIR contributes to the cisplatin-induced resistance in endometrial cancer cells. Biotechnol. Lett. 2017, 39, 1477–1484.
- Akhmedkhanov, A.; Zeleniuch-Jacquotte, A.; Toniolo, P. Role of exogenous and endogenous hormones in endometrial cancer: Review of the evidence and research perspectives. Ann. N. Y. Acad. Sci. 2001, 943, 296–315.
- Bhan, A.; Hussain, I.; Ansari, K.I.; Kasiri, S.; Bashyal, A.; Mandal, S.S. Antisense transcript long noncoding RNA (lncRNA) HOTAIR is transcriptionally induced by estradiol. J. Mol. Biol. 2013, 425, 3707–3722.
- Gupta, R.A.; Shah, N.; Wang, K.C.; Kim, J.; Horlings, H.M.; Wong, D.J.; Tsai, M.C.; Hung, T.; Argani, P.; Rinn, J.L.; et al. Long non-coding RNA HOTAIR reprograms chromatin state to promote cancer metastasis. Nature 2010, 464, 1071–1076.
- He, X.; Bao, W.; Li, X.; Chen, Z.; Che, Q.; Wang, H.; Wan, X.P. The long non-coding RNA HOTAIR is upregulated in endometrial carcinoma and correlates with poor prognosis. Int. J. Mol. Med. 2014, 33, 325–332.
- Mangelsdorf, D.J.; Thummel, C.; Beato, M.; Herrlich, P.; Schütz, G.; Umesono, K.; Blumberg, B.; Kastner, P.; Mark, M.; Chambon, P.; et al. The nuclear receptor superfamily: The second decade. Cell 1995, 83, 835–839.
- Lanz, R.B.; Chua, S.S.; Barron, N.; Söder, B.M.; DeMayo, F.; O’Malley, B.W. Steroid receptor RNA activator stimulates proliferation as well as apoptosis in vivo. Mol. Cell. Biol. 2003, 23, 7163–7176.
- Li, L.; Gu, M.; Bo, Y.; Shi, S.; Shan, Y.; Bao, L.; Yiwen, Y. Long non-coding RNA ROR promotes proliferation, migration and chemoresistance of nasopharyngeal carcinoma. Cancer Sci. 2016, 107, 1215–1222.
- Xu, J.; Qian, Y.; Ye, M.; Fu, Z.; Jia, X.; Li, W.; Xu, P.; Lv, M.; Huang, L.; Wang, L.; et al. Distinct expression profile of lncRNA in endometrial carcinoma. Oncol. Rep. 2016, 36, 3405–3412.
- Jiang, Y.; Malouf, G.G.; Zhang, J.; Zheng, X.; Chen, Y.; Thompson, E.J.; Weinstein, J.N.; Yuan, Y.; Spano, J.P.; Broaddus, R.; et al. Long non-coding RNA profiling links subgroup classification of endometrioid endometrial carcinomas with trithorax and polycomb complex aberrations. Oncotarget 2015, 6, 39865–39876.
- Liu, L.; Chen, X.; Zhang, Y.; Hu, Y.; Shen, X.; Zhu, W. Long non-coding RNA TUG1 promotes endometrial cancer development via inhibiting miR-299 and miR-34a-5p. Oncotarget 2017, 9, 31386–31394.
- Yang, L.; Zhang, J.; Jiang, A.; Liu, Q.; Li, C.; Yang, C.; Xiu, J. Expression profile of long non-coding RNAs is altered in endometrial cancer. Int. J. Clin. Exp. Med. 2015, 8, 5010–5021.
- Chen, S.; Chen, X.; Sun, K.X.; Xiu, Y.L.; Liu, B.L.; Feng, M.X.; Sang, X.B.; Zhao, Y. MicroRNA-93 promotes epithelial-mesenchymal transition of endometrial carcinoma cells. PLoS ONE 2016, 11, e0165776.
- Zhai, W.; Li, X.; Wu, S.; Zhang, Y.; Pang, H.; Chen, W. Microarray expression profile of lncRNAs and the upregulated ASLNC04080 lncRNA in human endometrial carcinoma. Int. J. Oncol. 2015, 46, 2125–2137.
- Li, W.; Li, H.; Zhang, L.; Hu, M.; Li, F.; Deng, J.; An, M.; Wu, S.; Ma, R.; Lu, J.; et al. Long non-coding RNA LINC00672 contributes to p53 protein-mediated gene suppression and promotes endometrial cancer chemosensitivity. J. Biol. Chem. 2017, 292, 5801–5813.
- Sun, Y.; Zou, X.; He, J.; Mao, Y. Identification of long non-coding RNAs biomarkers associated with progression of endometrial carcinoma and patient outcomes. Oncotarget 2017, 8, 52604–52613.
- Zhao, L.; Li, Z.; Chen, W.; Zhai, W.; Pan, J.; Pang, H.; Li, X. H19 promotes endometrial cancer progression by modulating epithelial-mesenchymal transition. Oncol. Lett. 2017, 13, 363–369.
- Kaufhold, S.; Bonavida, B. Central role of Snail1 in the regulation of EMT and resistance in cancer: A target for therapeutic intervention. J. Exp. Clin. Cancer Res. 2014, 33, 62.
- Wang, D.; Wang, D.; Wang, N.; Long, Z.; Ren, X. Long Non-Coding RNA BANCR promotes endometrial cancer cell proliferation and invasion by regulating MMP2 and MMP1 via ERK/MAPK signaling pathway. Cell. Physiol. Biochem. 2016, 40, 644–656.
- Guo, Q.; Qian, Z.; Yan, D.; Li, L.; Huang, L. LncRNA-MEG3 inhibits cell proliferation of endometrial carcinoma by repressing Notch signaling. Biomed. Pharmacother. 2016, 82, 589–594.
- Shang, C.; Lang, B.; Ao, C.N.; Meng, L. Long non-coding RNA tumor suppressor candidate 7 advances chemotherapy sensitivity of endometrial carcinoma through targeted silencing of miR-23b. Tumour Biol. 2017, 39.
- Guo, C.; Song, W.Q.; Sun, P.; Jin, L.; Dai, H.Y. LncRNA-GAS5 induces PTEN expression through inhibiting miR-103 in endometrial cancer cells. J. Biomed. Sci. 2015, 22, 100.
- Qiao, Q.; Li, H. LncRNA FER1L4 suppresses cancer cell proliferation and cycle by regulating PTEN expression in endometrial carcinoma. Biochem. Biophys. Res. Commun. 2016, 478, 507–512.
- Feng, S.; Yao, J.; Chen, Y.; Geng, P.; Zhang, H.; Ma, X.; Zhao, J.; Yu, X. Expression and functional role of reprogramming-related long noncoding RNA (lincRNA-ROR) in Glioma. J. Mol. Neurosci. 2015, 56, 623–630.
- Eades, G.; Wolfson, B.; Zhang, Y.; Li, Q.; Yao, Y.; Zhou, Q. lincRNA-RoR and miR-145 regulate invasion in triple-negative breast cancer via targeting ARF6. Mol. Cancer Res. 2015, 13, 330–338.
- Hou, P.; Zhao, Y.; Li, Z.; Yao, R.; Ma, M.; Gao, Y.; Zhao, L.; Zhang, Y.; Huang, B.; Lu, J. LincRNA-ROR induces epithelial-to-mesenchymal transition and contributes to breast cancer tumorigenesis and metastasis. Cell Death Dis. 2014, 5, e1287.
- Gao, S.; Wang, P.; Hua, Y.; Xi, H.; Meng, Z.; Liu, T.; Chen, Z.; Liu, L.M. ROR functions as a ceRNA to regulate Nanog expression by sponging miR-145 and predicts poor prognosis in pancreatic cancer. Oncotarget 2016, 7, 1608–1618.
- Zhan, H.X.; Wang, Y.; Li, C.; Xu, J.W.; Zhou, B.; Zhu, J.K.; Han, H.F.; Wang, L.; Wang, Y.S.; Hu, S.Y. LincRNA-ROR promotes invasion, metastasis and tumor growth in pancreatic cancer through activating ZEB1 pathway. Cancer Lett. 2016, 374, 261–271.
- Takahashi, K.; Yan, I.K.; Haga, H.; Patel, T. Modulation of hypoxia-signaling pathways by extracellular linc-RoR. J. Cell Sci. 2014, 127, 1585–1594.
- Takahashi, K.; Yan, I.K.; Kogure, T.; Haga, H.; Patel, T. Extracellular vesicle-mediated transfer of long non-coding RNA ROR modulates chemosensitivity in human hepatocellular cancer. FEBS Open Bio 2014, 4, 458–467.
- Zhou, X.; Gao, Q.; Wang, J.; Zhang, X.; Liu, K.; Duan, Z. Linc-RNA-RoR acts as a “sponge” against mediation of the differentiation of endometrial cancer stem cells by microRNA-145. Gynecol. Oncol. 2014, 133, 333–339.
- Rezaei, M.; Emadi-Baygi, M.; Hoffmann, M.J.; Schulz, W.A.; Nikpour, P. Altered expression of LINC-ROR in cancer cell lines and tissues. Tumour Biol. 2016, 37, 1763–1769.
- Chen, B.J.; Byrne, F.L.; Takenaka, K.; Modesitt, S.C.; Olzomer, E.M.; Mills, J.D.; Farrell, R.; Hoehn, K.L.; Janitz, M. Transcriptome landscape of long intergenic non-coding RNAs in endometrial cancer. Gynecol. Oncol. 2017, 147, 654–662.
- Wilczynski, M.; Danielska, J.; Dzieniecka, M.; Szymanska, B.; Wojciechowski, M.; Malinowski, A. Prognostic and clinical significance of miRNA-205 in endometrioid endometrial cancer. PLoS ONE 2016, 11, e0164687.
- Jayaraman, M.; Radhakrishnan, R.; Mathews, C.A.; Yan, M.; Husain, S.; Moxley, K.M.; Song, Y.S.; Dhanasekaran, D.N. Identification of novel diagnostic and prognostic miRNA signatures in endometrial cancer. Genes Cancer 2017, 8, 566–576.
- He, Z.; Xu, H.; Meng, Y.; Kuang, Y. miR-944 acts as a prognostic marker and promotes the tumor progression in endometrial cancer. Biomed. Pharmacother. 2017, 88, 902–910.
- Ravo, M.; Cordella, A.; Rinaldi, A.; Bruno, G.; Alexandrova, E.; Saggese, P.; Nassa, G.; Giurato, G.; Tarallo, R.; Marchese, G.; et al. Small non-coding RNA deregulation in endometrial carcinogenesis. Oncotarget 2015, 6, 4677–4691.
- Ahsen, M.E.; Boren, T.P.; Singh, N.K.; Misganaw, B.; Mutch, D.G.; Moore, K.N.; Backes, F.J.; McCourt, C.K.; Lea, J.S.; Miller, D.S.; et al. Sparse feature selection for classification and prediction of metastasis in endometrial cancer. BMC Genom. 2017, 27, 233.
- Bahubeshi, A.; Tischkowitz, M.; Foulkes, W.D. miRNA processing and human cancer: DICER1 cuts the mustard. Sci. Transl. Med. 2011, 3.
- Hu, Z.; Zhang, H.; Tang, L.; Lou, M.; Geng, Y. Silencing nc886, a non-coding RNA, induces apoptosis of human endometrial cancer cells-1A in vitro. Med. Sci. Monit. 2017, 16, 1317–1324.
- The Cancer Genome Atlas; National Cancer Institute, National Human Genome Research Institute. Available online: (accessed on 27 March 2018).
- Ihira, K.; Dong, P.; Xiong, Y.; Watari, H.; Konno, Y.; Hanley, S.J.; Noguchi, M.; Hirata, N.; Suizu, F.; Yamada, T.; et al. EZH2 inhibition suppresses endometrial cancer progression via miR-361/Twist axis. Oncotarget 2017, 8, 13509–13520.
- Gong, B.; Yue, Y.; Wang, R.; Zhang, Y.; Jin, Q.; Zhou, X. Overexpression of microRNA-194 suppresses the epithelial-mesenchymal transition in targeting stem cell transcription factor Sox3 in endometrial carcinoma stem cells. Tumor Biol. 2017, 39.
- Chen, H.X.; Xu, X.X.; Tan, B.Z.; Zhang, Z.; Zhou, X.D. MicroRNA-29b inhibits angiogenesis by targeting VEGFA through the MAPK/ERK and PI3K/Akt signaling pathways in endometrial carcinoma. Cell. Physiol. Biochem. 2017, 41, 933–946.
- Alowayed, N.; Salker, M.S.; Zeng, N.; Singh, Y.; Lang, F. LEFTY2 controls migration of human endometrial cancer cells via focal adhesion kinase activity (FAK) and miRNA-200a. Cell. Physiol. Biochem. 2016, 39, 815–826.
- Sharma, S.; Kelly, T.K.; Jones, P.A. Epigenetics in cancer. Carcinogenesis 2010, 31, 27–36.
- Esteller, M. Epigenetics in cancer. N. Engl. J. Med. 2008, 358, 1148–1159.
- Hosseini, E.S.; Meryet-Figuiere, M.; Sabzalipoor, H.; Kashani, H.H.; Nikzad, H.; Asemi, Z. Dysregulated expression of long noncoding RNAs in gynecologic cancers. Mol. Cancer 2017, 16, 107.
- Ivanga, M.; Labrie, Y.; Calvo, E.; Belleau, P.; Martel, C.; Luu-The, V.; Morissette, J.; Labrie, F.; Durocher, F. Temporal analysis of E2 transcriptional induction of PTP and MKP and downregulation of IGF-I pathway key components in the mouse uterus. Physiol. Genom. 2007, 29, 13–23.