The South-Pyrenean Basin (northeastern Spain) has yielded a rich and diverse record of Upper Cretaceous (uppermost Campanian−uppermost Maastrichtian) vertebrate fossils, including the remains of some of the last European dinosaurs prior to the Cretaceous-Paleogene (K-Pg) extinction event. In this work, we update and characterize the vertebrate fossil record of the Arén Sandstone and Tremp formations in the Western Tremp Syncline, which is located in the Aragonese area of the Southern Pyrenees. The transitional and continental successions of these sedimentary units are dated to the late Maastrichtian, and exploration of their outcrops has led to the discovery of numerous fossil remains (bones, eggshells, and tracks) of dinosaurs, including hadrosauroids, sauropods, and theropods, along with other tetrapods such as crocodylomorphs, testudines, pterosaurs, squamates, and amphibians. In particular, this fossil record contains some of the youngest lambeosaurine hadrosaurids (Arenysaurus and Blasisaurus) and Mesozoic crocodylomorphs (Arenysuchus and Agaresuchus subjuniperus) in Europe, complementing the lower Maastrichtian fossil sites of the Eastern Tremp Syncline.
- Fossil
- The South-Pyrenean Basin
1. Background
The Cretaceous-Paleogene (K-Pg) extinction event is undoubtedly one of the most debated topics in the evolutionary history of life on the planet. Ever since a catastrophic meteorite impact at the end of the Maastrichtian was proposed as the major cause of the extinction [1], scientific debate on this event has been of ongoing significance. At the end of the Cretaceous, a set of destabilizing events occurred on Earth, including a marine regression [2], climate changes [3][4], the volcanic activity of the Deccan Volcanic Province (India) with the emission of a huge amount of gases and volcanic material into the atmosphere [5][6][7][8], and the impact of an asteroid in Chicxulub (Mexico) 66 Ma ago [1][9][10][11][12]. Although all these causes seem to have contributed to the extinction to a certain degree, the meteorite impact hypothesis shows the most solid arguments for having been the major disturbing mechanism [12][13][14][15][16].
Whatever the cause, the K-Pg extinction eradicated nearly 70% of the living species on Earth [17][18]. Among vertebrates, this event led to the disappearance of several groups, including non-avian dinosaurs, enantiornithine birds, pterosaurs, mosasaurs, plesiosaurs, and several lineages of crocodylomorphs, among others [19][20][21][22][23]. However, the mechanism by which they became extinct and how fast they did so remain difficult questions for researchers, as is the issue of how determinant the Chicxulub impact was on the stability of the ecosystems. Except for the Hell Creek Formation in North America, whose vertebrate faunas are well known [22][24][25] and their chronostratigraphic framework is well constrained [26][27][28][29], the main difficulty in assessing the end-Cretaceous extinction is the lack of well-studied sedimentary formations with vertebrate remains encompassing the K-Pg boundary. In Europe, a great effort has been made in recent decades to characterize the terrestrial uppermost Cretaceous-Paleocene formations, especially in Spain, France, and Romania (e.g., [30][31][32][33]). The best-known deposits are those from the so-called Ibero-Armorican island, which encompassed the current south of France and the northeast of Spain (Languedoc, Provence, and the Pyrenees), and other outcrops in the east, northwest, and center of Spain and part of Portugal. Of these regions, the South-Pyrenean Basin is the best-known area. Since the end of the 20th century, several research teams have worked on the uppermost Campanian–Danian outcrops in this area, improving our knowledge of the biodiversity of fossil vertebrates, the environments they inhabited, and the chronostratigraphic framework [31][33][34].
2. Fossil Record
The sedimentary succession of the Tremp Fm in the Western Tremp Syncline here under study encompasses sedimentary rocks belonging to the ‘Grey Garumnian’ and ‘Lower Red Garumnian’, and therefore, the late Maastrichtian between the upper part of chron C30n and chron C29r. The studied area is the part of the Tremp Syncline where the thickest succession of the upper Maastrichtian is preserved (more than 210 m). In these upper Maastrichtian sediments, there is a diverse and significant record of vertebrate fossils, including avian and non-avian dinosaurs, crocodylomorphs, testudines, squamates, amphibians, and fishes. More than 1200 fossil remains have been recovered from the hilly outcrops, with 57 different fossil localities identified. In order to facilitate our explanation of the Western Tremp Syncline fossil record, we have clustered the paleontological sites by their closeness to certain reference stratigraphic logs, which are called the Campo (I), Larra (II), Rin (III), Serraduy (IV), Beranuy (V), Isclés (VI), and Areny (VII) sections (Figure 1). All the fossils are housed at the Natural Science Museum of the University of Zaragoza (Spain) (MPZ) [35].

Figure 1. Upper Maastrichtian outcrops of the Western Tremp Syncline (Aragón, NE Spain). (a) Campo section (I); (b) Serraduy section (IV); (c) Isclés section (VI); (d) Arén/Areny section (VII).
2.1. Dinosauria
2.1.1. Hadrosauroidea
Hadrosauroid dinosaurs are the clade of Cretaceous ornithopods with the most abundant fossil record, especially in the Northern Hemisphere. In Europe, the best record of hadrosauroids has been recovered from France and Spain. In Spain, it is concentrated principally in the South-Pyrenean Basin (the provinces of Huesca and Lleida, NE Spain) [23][36][37][38][39][40][41][42][43].
In the Western Tremp Syncline, hadrosauroids are recorded in the upper Maastrichtian sediments of the Arén Sandstone Fm and the ‘Grey and Lower Red Garumnian’ of the Tremp Fm; these are among the youngest non-avian dinosaurs in the world [44]. The first hadrosauroid bones were found in the 1990s near the locality of Arén (Areny in Catalan) (Huesca, Aragón) by the geologists Lluís Ardèvol and Fabián López Olmedo during geological mapping work. Early work on several sites (Blasi 1 to 5 and Blasi 3,4) by a multidisciplinary team yielded fossil remains of indeterminate euhadrosaurids together with bones and eggshells of several dinosaurs and other terrestrial and aquatic vertebrates [45]. Later studies on specimens from the Blasi 1 and Blasi 3 sites resulted in the erection of two lambeosaurine hadrosaur species: Blasisaurus canudoi Cruzado-Caballero, Pereda-Suberbiola, and Ruiz-Omeñaca 2010a [46] (Figure 1 and Figure 2) and Arenysaurus ardevoli Pereda-Suberbiola, Canudo, Cruzado-Caballero, Barco, López-Martínez, Oms and Ruiz-Omeñaca [36][47] (Figure 1 and Figure 3). Both sites fall within the upper part of chron C30n [36]. These two species are recovered within Arenysaurini, which is a recently erected clade of lambeosaurines from Europe [48].

Figure 2. Main tetrapod remains from the Western Tremp Syncline. (a) Cranial elements of Arenysaurus ardevoli (MPZ2008/17, MPZ2008/256, MPZ2008/258, MPZ2008/259, MPZ2011/01), in left lateral view (modified from Cruzado-Caballero et al., 2013); (b) cranial elements of Blasisaurus canudoi (MPZ 99/664, MPZ 99/665, MPZ99/666a, MPZ99/666b, MPZ99/667, MPZ 2009/348), in left lateral view; (c) mid-caudal vertebra of Arenysaurus (MPZ204/480), in left lateral view (modified from Cruzado-Caballero et al., 2013); (d) articulated mid-caudal vertebrae of the small hadrosaurid from Serraduy (MPZ 2013-371), in left lateral view; (e) femur (proximal end) of Titanosauria indet. from Serraduy (MPZ 99/143), in posterior view (modified from Puértolas-Pascual et al., 2018); (f–h) posterior caudal vertebra of Titanosauria indet. (MPZ2021/1), in anterior view (f); dorsal view (g); left lateral view (h); (i) cervical vertebra of Ornithuromorpha indet. (MPZ 2019/264), in left lateral view; (j) cf. Arcovenator tooth (MPZ 2017/804), in lingual view; (k) pedal ungual II of Dromaeosauridae indet. (MPZ 2019/196), in lateral view; (l) fragmentary bone of Pterosauria indet. (MPZ 2021/54) (note the thin cortex in the transverse section); (m) skull of Agaresuchus subjuniperus (MPZ 2012/288), in dorsal view; (n) skull of Arenysuchus gascabadiolorum (MP Z2011/184), in dorsal view.
In addition to this, other remains of indeterminate hadrosaurids and euhadrosaurids have been described from the Blasi sites [38][40][49][50][51]. The findings from these sites have also led to the first description of a pathological bone from a hadrosaurid in Spain [52] and the first paleo-neuroanatomical description of a European lambeosaurine, Arenysaurus ardevoli [53]. Recent studies on the paleohistology of the hadrosauroids from the Blasi sites reveal the presence of hadrosaurid individuals at different ontogenetic stages, including early and late juveniles, subadults, and mature adults [54][55]. New areas with hadrosaurid remains have been found in the vicinities of Serraduy (Isábena, Huesca, Aragón) and Beranuy (Huesca, Aragón) (Figure 2) [23][44][39][41][56][57][58]. The new sites are characterized by the presence of fossil remains of the smallest adult hadrosaurids (maybe affected by insular dwarfism) from Europe, which coexisted alongside larger hadrosaurids [41] (Figure 4d).
This rich osteological record of hadrosauroids in the Western Tremp Syncline is complemented by several track sites. These tracks appear in several levels from Arén to Campo (Huesca, Aragón), with large ornithopod footprints, many of which have been referred to the ichnogenus Hadrosauropodus [31][44][59][60] (Figure 1 and Figure3), spanning from the top of chron C30n into chron C29r. Recently, in the Blasi 2B site, eggshells attributable to hadrosaurid dinosaurs have been tentatively referred to Spheroolithus aff. europaeus Sellés, Vila, Galobart 2014 [61][62].

Figure 3. Tetrapod tracks from the Western Tremp Syncline. (a) Hadrosauropodus trackway from the Areny 1 site; (b) foot cast of a hadrosaurid dinosaur from the 172-i/04/a site; (c) crocodylomorph tracks from the Serraduy Norte site.
2.1.2. Sauropoda
The sauropod remains in the Western Tremp Syncline are very scarce compared to those in the eastern part, where titanosaur bones, eggshells, and tracks are moderately abundant [33][63][64]. A remarkable specimen is the proximal half of a femur (MPZ 99/143) that probably corresponds to a large and indeterminate titanosaur [63][65] (Figure 2). MPZ 99/143 was recovered northwest of the town of Serraduy, in the ‘Grey Garumnian’ unit (‘Femur’ site in Figure 2). Interestingly, the femur was originally correlated to the top of chron C30n, but the chronostratigraphical data indicate that this fossil lies within chron C29r [31][44]. Thus, this femur is one of the youngest records of titanosaurian sauropods in the Ibero-Armorican island, along with those recorded in fossil sites in the Catalonia region, including the ‘Molí del Baró-2’ femur [63], the vertebra from ‘El Portet’ site [66], and the skin impressions and footprints from the ‘Mirador de Vallcebre’ [67]. In addition, Cruzado-Caballero et al. (2012) [39] reported a caudal vertebral centrum that was found in the ’Lower Red Garumnian’ unit near Serraduy and Beranuy. The caudal vertebra (MPZ 2021/1) is from the ‘Barranco Serraduy 4’ site (Serraduy) (Figure 2), which is situated stratigraphically above the ‘Femur’ site, making it the youngest evidence of sauropods in the Western Tremp Syncline. MPZ 2021/1 is a slightly deformed centrum from a posterior caudal vertebra, which is elongated craniocaudally and compressed dorsoventrally. It is amphiplatyan, with both articular surfaces flat to slightly concave and with a rounded contour (Figure 2). In anteroposterior view, the centrum has a subcircular outline (Figure 2). Its ventral surface is slightly concave and lacks chevron facets (Figure 2). Together with its length and the absence of transverse processes, this indicates that it was situated distally in the caudal series [68]. The neural arch is not preserved, but its attachment facets can be observed in the anterior area of the centrum (Figure 2), which is a synapomorphy of Titanosauriformes [69]. The amphiplatyan condition in the middle and posterior caudal vertebrae is plesiomorphic within Titanosauria [69][70]. Some basal titanosaurs show this condition, as is the case of Andesaurus [71][72] or the distalmost vertebrae of Lirainosaurus [68]. Therefore, we tentatively refer it to Titanosauriformes indet., but for the reasons mentioned above, its ascription to Titanosauria cannot be ruled out.
2.1.3. Theropoda
Theropod fossils are scarce in the Western Tremp Syncline, and these are mainly represented by teeth, eggshells, and some isolated bones. Torices et al. (2015) [73] describe several teeth from the Blasi sites of Arén/Areny (Figure 2). They identify one morphotype as Coelurosauria indet. (MPZ 98/79 to 82) and three morphotypes belonging to maniraptoran theropods, including Richardoestesia sp. (MPZ 98/72 to 74, MPZ 2004/7), cf. Paronychodon (MPZ 98/76 to 78), and Dromaeosauridae indet. (MPZ 2004/6). Finally, they describe two different morphotypes of large teeth whose assignation is problematic and that are referred to Theropoda indet. 1 (MPZ 98/67, MPZ 2004/3 to 5, 8) and Theropoda indet. 2 (MPZ 98/68), although a possible relation with neoceratosaurs is suggested. In fact, these two morphotypes were identified by Pérez-García et al. (2016) as cf. Arcovenator [74], which is an abelisaurid species from the Campanian of southern France. The Blasi sites 1, 2, and 3 are dated to within chron C30n [36] (Figure 2). Two more theropod teeth have been described from the fossil sites of 172-i/04/e (Serraduy) and Larra 4 (Valle de Lierp) [44]. The first tooth (MPZ 2017/804) (Figure 4j) is large and resembles the Theropoda indet. morphotype 1 (cf. Arcovenator) from Torices et al. (2015) [73], and the second one has been identified as Coelurosauria indet. Both sites are situated in outcrops of the ‘Lower Red Garumnian’ dated to within chron C29r [44].
Postcranial fossils of theropods are not very common and are usually fragmentary. A pedal ungual II (MPZ 2019/196) (Figure 4k) and the proximal part of an ulna (MPZ 2019/194) from a dromaeosaurid theropod were found at the Larra 4 site (Valle de Lierp, Huesca, Aragón, Spain) (Figure 2) [57]. Other sites in the Serraduy area have yielded fragmentary remains of indeterminate theropods (Figure 2). As regards avian theropods, a cervical vertebra from a large ornithuromorph bird has recently been described from the Tremp Fm outcrops between Serraduy and Biascas de Obarra (Beranuy, Huesca) (MPZ 2019/264) [75] (Figure 2 and Figure 4i). It has been dated as uppermost Maastrichtian (C29r) [44] and represents the youngest record of a Mesozoic bird in Europe.
Up to now, theropod eggshells have only been found in the site Blasi 2B (‘Grey Garumnian’ at Arén/Areny, C30n; Figure 2). This site has a diverse theropod eggshell assemblage, which was briefly described by López-Martínez et al. (1999) [76]. The authors recognized up to six different types of prismatoolithid eggshells, whereas more recent research [62][77] recognized at least four different types (Figure 6a,b), including two different morphotypes attributable to the oogenus Pseudogeckoolithus, which has recently been referred to maniraptoran theropods [78]. Further research is necessary to ascertain the exact number of theropod ootaxa present at Blasi 2B.

Figure 4. Tetrapod eggshells from the Western Tremp Syncline. (a) Pseudogeckoolithus sp. from the Blasi 2B site; (b) Prismatoolithidae indet. from Blasi 2B; (c) Krokolithes sp. from the Veracruz 1 site.
2.2. Pterosauria
The presence of pterosaurs in the upper Maastrichtian of the Tremp Syncline has only been reported from the site of Torrebilles-2 in the Eastern Tremp Syncline, within chron C29r [79]. In the Western Tremp Syncline, Puértolas-Pascual et al. (2018) [44] reported a possible mandible of a pterosaur from the upper part of the ‘Lower Red Garumnian’ near Serraduy (Isábena). This specimen has been reexamined, and although its identification as a dentary has been refuted, its affinity to a pterosaurian bone cannot be ruled out. However, until a future study identifies this bone more precisely, it cannot be assigned to Pterosauria. Nevertheless, we have identified a fragment of a long bone from the Blasi 5 site (Figure 2) that shows a very thin cortex (thinner than the pterosaur bones from Barranc de Torrebilles-2 [79]) and is hollow inside. This bone could be the first pterosaur fossil identified in the Western Tremp Syncline. Blasi 5 is situated in the upper part of the ‘Lower Red Garumnian’ and is dated to within chron C29r [36].
2.3. Crocodylomorpha
The crocodylomorph record in the Western Tremp Syncline is dominated by eusuchians. Two skulls belonging to two different genera have been identified. The first one is Arenysuchus gascabadiolorum Puértolas, Canudo, Cruzado-Caballero 2011 [80] (MPZ 2011/184) (Figure 2), from the Elias site near Arén/Areny. Phylogenetically, MPZ 2011/184 was initially placed within Crocodyloidea (crown-group Crocodylia) [80], but later cladistic studies have situated it as a more basal eusuchian within Allodaposuchidae [81][82][83][84][85]. The second species is the allodaposuchid Agaresuchus subjuniperus Puértolas-Pascual, Canudo, Moreno-Azanza 2014 [86] (MPZ 2012/288) (Figure 2). MPZ 2012/288 was initially identified as a member of the genus Allodaposuchus [86], but it was later reassigned to Agaresuchus [84]. This crocodylomorph comes from the Amor 3 site near the town of Serraduy, from one of the uppermost levels of the ‘Lower Red Garumnian’. As such, it could be one of the youngest crocodylomorphs on the Ibero-Armorican island before the K-Pg extinction. In addition, allodaposuchids are also represented by isolated teeth in several sites throughout the C30n−C29r interval [23][44][57][58][87]. All these teeth are conical with pointed crowns, showing the typical morphology of crocodylomorphs with a generalist diet. These dental morphologies have been observed in several allodaposuchid species from the Late Cretaceous of Europe (e.g., [87]). As the presence of other crocodylomorph clades with generalist dentition cannot be ruled out, these teeth were assigned to cf. Allodaposuchidae, since this is the most abundant clade in this region and time interval.
Gavialoidea is another clade of crocodylomorphs that may be present in the upper Maastrichtian of the Western Tremp Syncline. A few elongated conical teeth with basiapical ridges have been assigned to cf. Thoracosaurus. These are restricted to the transitional environments of the Arén Fm and the ‘Grey Garumnian’ unit of the Tremp Fm close to Arén/Areny, Beranuy and Serraduy (Figure 2) [23][44][87].
Hylaeochampsidae are represented by tribodont teeth from the Blasi 2B site, which were identified as cf. Acynodon (MPZ-2017/1137) [23][44][45][87]. The eusuchian record is augmented by teeth, osteoderms, and vertebrae from the Blasi and Serraduy sites, whose taxonomical position within Eusuchia is difficult to assign with precision. They are accordingly identified as Eusuchia indet. [23][44][45][87].
López-Martínez et al. (2001) [45] pointed out the presence of “trematochampsid”-like and alligatoroid teeth from the sites of Blasi 1, 2, and 3. However, although the authors did not provide pictures or specimen numbers, the morphotypes in question probably correspond to more recently erected taxa that had not been described at the time of publication of that paper. The “trematochampsid”-like teeth may correspond to non-eusuchian crocodylomorphs more typical of the Late Cretaceous of Europe, such as Sabresuchus or Doratodon, and the alligatorid teeth probably correspond to allodaposuchids. In addition, Blanco et al. (2020) [87] mentioned the presence of a conical tooth (MPZ 2010/948) with enamel striations and crenulated carinae assigned to Mesoeucrocodylia indet.
There are also crocodylomorph eggshells from the Blasi 2 site (upper part of C30n, Figure 2). These were first reported as megaloolithid eggshells [76], but they were later [88] described as having a crocodyloid morphotype and were identified as Krokolithes sp. Hirsch, 1985 [89], implying that these eggs were laid by crocodylomorphs. Something similar has occurred with the eggshells found at the Veracruz 1 site close to Biascas de Obarra,, which were first identified as hadrosaurid eggshells [58] (Figure 4), but after a more thorough study, their crocodylomorph affinities have been ascertained, and a description is in preparation.
Finally, the crocodylomorph record also includes several swimming and plantigrade tracks from the Serraduy, Beranuy, and Campo outcrops, all within chron C29r [23][90][91] (Figure 3). This is the youngest record of crocodylomorph tracks in Europe. These tracks represent digit scratch marks produced by the manus and pes of buoyant crocodylomorphs, and they have been assigned to the ichnogenus Characichnos. One pedal impression has been assigned to cf. Crocodylopodus, and although its assignation cannot be confirmed with certainty due to the scarce material, this is the youngest occurrence of this ichnotaxon [90].
2.4. Testudines
Testudines are represented mainly by disarticulated plates of the carapace or the plastron, which appear at most of the paleontological sites from the topmost part of the Arén Fm to the upper levels of the ‘Lower Red Garumnian’. Most of these remains show fine ornamentation comprising thin dichotomic grooves, which is a distinctive character of the bothremydids [92]. Among these remains, Murelaga and Canudo (2005) [93] describe several plates from the Blasi sites near Arén/Areny , including nuchal, pleural, and peripheral plates, a hyoplastron, a hypoplastron, and a xiphiplastron from bothremydid turtles. At the site of Rin 2, near the town of Serraduy (Isabena municipality), situated in the topmost part of the Arén Fm, Murelaga and Canudo (2005) [93] describe a xiphiplastron and a mesoplastron from a bothremydid. Pérez-Pueyo et al. (2019a, 2019b) [57][58] also describe indeterminate plates from this kind of turtle from the Larra 4 (Valle de Lierp) and Veracruz 1 (Biascas de Obarra, Beranuy) sites . Thus, the record of this group of pleurodiran turtles extends from the upper part of chron C30r to near the K-Pg boundary interval. It is also important to note that Murelaga and Canudo (2005) [93] identify a peripheral plate from a solemydid turtle from the Blasi 2 site . This shows its characteristic vermiculate ornamentation, although it is not well preserved.
2.5. Amphibia and Squamata
The Blasi 2 site has yielded a rich microvertebrate fossil assemblage, which includes the bones of small tetrapods, mainly amphibians and squamates [94] . Amphibian remains dominate, with at least one albanerpetontid (resembling the North American taxon Albanerpeton nexuosum) and two anurans, a discoglossid and a palaeobatrachid. The squamate remains comprise at least two undetermined lizards, one anguid lizard, and a snake. Blasi 2B is dated to the top of chron C30n in the ’Grey Garumnian’ and is the only well-studied microvertebrate site in the Western Tremp Syncline. However, it is noteworthy that the Larra 4 site (Valle de Lierp) (C29r) has yielded remains from discoglossid amphibians [44], making it the youngest microvertebrate site in the Western Tremp Syncline.
2.6. The Tetrapod Fossil Record from the Upper Maastrichtian of the Ibero-Armorican Island
The tetrapod fossil record of the Western Tremp Syncline adds several unique taxa to the late Maastrichtian assemblages of the Ibero-Armorican island, yielding the youngest record of some groups prior to the Paleocene. To date, the upper Maastrichtian record of the Ibero-Armorican island is limited to the South Pyrenean Basin in northeast Spain; the Sobrepeña Fm, Torme Fm, and equivalent outcrops in northwest Spain [95][96]; east Spain near Tous (Valencia) [97][98]; and the Haute-Garonne and Aude departments in southern France [33][99]. During the Maastrichtian, the dinosaur faunas underwent a change in dominant herbivores during the so-called “Maastrichtian Dinosaur Turnover” [33][34][100]. During the early Maastrichtian, ecosystems were inhabited by rhabdodontid ornithopods, titanosaurian sauropods and ankylosaurs, whereas in the late Maastrichtian, these communities were replaced by hadrosaurid ornithopods and new titanosaurian forms. However, nodosaurid ankylosaurians still persisted up to chron C30r, coexisting with these new assemblages for nearly 2 Myr [33].
Lambeosaurine hadrosaurids were present in the Ibero-Armorican island from the late early Maastrichtian, mostly recorded from the Pyrenees (Spanish and French). The lambeosaurine from Els Nerets (Vilamitjana, Catalonia, NE Spain) is the oldest evidence of hadrosaurids in Europe, which was dated to within chron C31r [43]. Lambeosaurines are also present within chron C29r, with some fossils falling very close to the K-Pg boundary. At present, there are five species of lambeosaurine hadrosaurids described from the region, comprising Adynomosaurus from the Costa de Les Solanes site (Basturs, Catalonia) [101], Arenysaurus [36], Blasisaurus [46] from Areny and Blasi sites (Ribagorza, Aragón, NE Spain), Pararhabdodon from the Sant Romà d’Abella site (Lleida, Catalonia, NE Spain) [42][102][103][104], and Canardia from the Lacarn and Tricouté sites (Haute-Garonne, southern France) [42]. Additional hadrosauroid remains include the aforementioned lambeosaurinae from Els Nerets [43] and other indeterminate lambeosaurines from Basturs Poble and Les Llaus (Lleida, Catalonia) [42][105][106][107]; a non-hadrosaurid hadrosauroid from Fontllonga-R (Fontllonga, Catalonia, NE Spain) [37]; an indeterminate euhadrosaurid from Blasi 3,4 [40]; and a small hadrosaurid from Serraduy [41]. Outside the Pyrenees, there is a dentary from La Solana (Tous, Valencia, E Spain) that has been identified as belonging to an indeterminate hadrosaurid [37][108]. Finally, there is a hadrosauroid femur from the Albaina site (Laño, Condado de Treviño, Burgos, NW Spain) [109]. With six to twelve taxa, hadrosauroids are the most speciose clade of dinosaurs in the Tremp Basin, five to eight of them being lambeosaurine hadrosaurids (Table 1, Figure 7 and Figure 8).
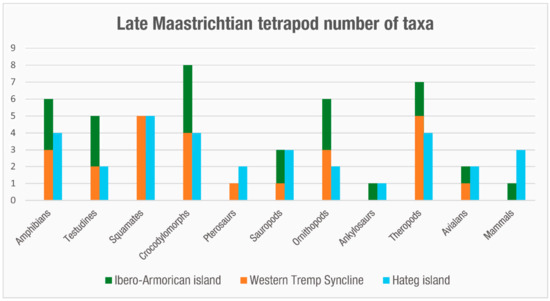
Figure 7. Bar chart with the minimum number of tetrapod taxa (genera) present in the Ibero-Armorican island and Haţeg island during the late Maastrichtian.

Figure 8. Dinosaur groups and species occurrences in the Ibero-Armorican island and Haţeg island during the late Maastrichtian. Haţeg island data are based on [32] with updated information. The yellow band marks the temporal interval used for comparison, whereas the green band marks the last ≈350 ka of the Maastrichtian, with a record only in the Ibero-Armorican island. Magnetochronostratigraphic scale based on [110] and calibration for the K-Pg boundary based on [28].
Table 1. Number of tetrapod taxa present on the Ibero-Armorican and Hațeg islands during the late Maastrichtian. Red numbers mark possible additional taxa.
| Taxon | Ibero-Armorican Island |
Western Tremp Syncline |
Hațeg Island |
|---|---|---|---|
| Amphibia | 6 | 3 | 4 |
| Anura | 4 | 2 | 3 |
| Albanerpetontidae | 1 | 1 | 1 |
| Salamandridae | 1 | − | − |
| Squamata | 6 + 2? | 3 + 2? | 5 |
| ‘Scincomorpha’ | 1 | 1? | 1 |
| Anguimorpha | 1 | 1 | 1 |
| Teiioidea | 1 | − | 1 |
| Borioteiioidea | − | − | 1 |
| Scleroglossa | 1 | 1 | − |
| Iguanidae | 1 | 1? | − |
| Amphisbaenia | 1? | − | − |
| Varanoidea | 1? | − | − |
| Alethinophidia | 1 | 1 | 1 |
| Testudines | 5 | 2 | 2 |
| Meiolaniformes | − | − | 1 |
| Pan-Pleurodira | 3 | 1 | 1 |
| Pan-Cryptodira | 2 | 1 | − |
| Crocodylomorpha | 8 | 4 | 4 |
| Notosuchia (Doratodon) | 1 | − | 1 |
| Neosuchia (‘Atoposauridae’) | 1 | - | 1 |
| Basal Eusuchia (Allodaposuchidae) | 4 | 2 | 1 |
| Basal Eusuchia (Hylaeochampsidae, cf. Acynodon) | 1 | 1 | 1 |
| Eusuchia (Gavialoidea) | 1 | 1 | − |
| Pterosauria | 1 | 1 | 2 |
| Azhdarchidae | 1 | 1 | 2 |
| Dinosauria | 18 + 1? (6) | 10 (1) | 12 + 1? (3) |
| Sauropoda | 3 | 1 | 3 |
| Titanosauria | 3 | 1 | 3 |
| Theropoda | 8 + 1? | 6 | 6 + 1? (3) |
| Alvarezsauridae | − | - | 1? |
| Abelisauroidea | 1 | 1 | − |
| Coelurosauria indet. | 1 | 1 | − |
| Maniraptora | 5 | 3 | 3 (2) |
| Paraves with uncertain affinities (Balaur, Elopteryx) | − | − | 1 (1) |
| Enantiornithes | 1? | − | 1 |
| Ornithuromorpha | 1 | 1 | 1 |
| Ornithopoda | 6 (6) | 3 (1) | 2 |
| Rhabdodontidae | − | − | 1 |
| Hadrosauroidea | 6 (6) | 3 (1) | 1 |
| Ankylosauria | 1 | - | 1 |
| Nodosauridae | 1 | − | 1 |
| Mammalia | 1? | 3 | |
| Multituberculata | − | − | 3 |
| Theria | 1? | − | − |
Ankylosaurs are represented during the late Maastrichtian by isolated and fragmentary material referred to nodosaurids from several sites in the Southern Pyrenees within the Lleida province (Catalonia, NE Spain), including Els Nerets [111], Fontllonga-6 [76], and Biscarri [112]. They are also present at the Lestaillats site, in the Petite Pyrénées (Haute-Garonne, southern France) [99]. Their last occurrence is documented at the Fontllonga-6 and Lestaillats sites, dated to within chrons C30r and C30n (Figure 8).
Titanosaurs from the upper Maastrichtian of the Ibero-Armorican island consist mainly of three undetermined but distinct taxa represented by three femur morphotypes [63] (Figure 7 and Figure 8). The femur from Serraduy corresponds to a large titanosaur, whereas the other two femora represent small-medium titanosaurs. Although not formally described, these titanosaurs represent different taxa from those of the early Maastrichtian assemblage [63]. This distinction is additionally supported by the distinct ootaxa association reported from the pre- and post-turnover assemblages, respectively [33][64].
Theropods from the late Maastrichtian of the Ibero-Armorican island are mainly abelisaurids and maniraptorans [44][57][73][113][114], and they have been found only in the South-Pyrenean Basin. Due to their fragmentary and incomplete nature, the number of taxa is difficult to determine. Based on tooth morphotypes from the Southern Pyrenees, at least one abelisaurid taxon inhabited the island during the late Maastrichtian (Theropoda indet. 1 and 2 or cf. Arcovenator; [73]) (Figure 7 and Figure 8). Maniraptorans are also represented mainly by teeth from the Southern Pyrenees, with at least three taxa identified in this way (Richardoestesia, Paronychodon, and Dromaeosauridae indet. from [73]), and by the troodontid Tamarro insperatus Sellés, Vila, Brusatte, Currie and Galobart 2021 [115], which were recently described on the basis of skeletal remains (Table 1, Figure 7 and Figure 8). However, the real abundance of theropods is hard to establish, since there are several fragmentary skeletal remains attributable to undetermined dromaeosaurids, and an oological record comprising several ootaxa of maniraptoran-like eggshells, including Prismatoolithus trempii Sellés, Vila, Galobart 2014 [116], and Pseudogeckoolithus Vianey-Liaud and López-Martínez 1997 [117] ([62][78][116]). Avialan dinosaurs are represented by the giant ornithuromorph bird from Beranuy [75] and a putative enantiornithine from southern France [118] (Table 1, Figure 7 and Figure 8).
The pterosaur record in the late Maastrichtian is scarce, with some isolated and fragmentary bones from the Pyrenees of France [119][120][121] and Spain [79] and from the upper Maastrichtian outcrops near Valencia (Spain) [122][123] (Figure 9). All of them have been identified as belonging to undetermined giant azhdarchids.
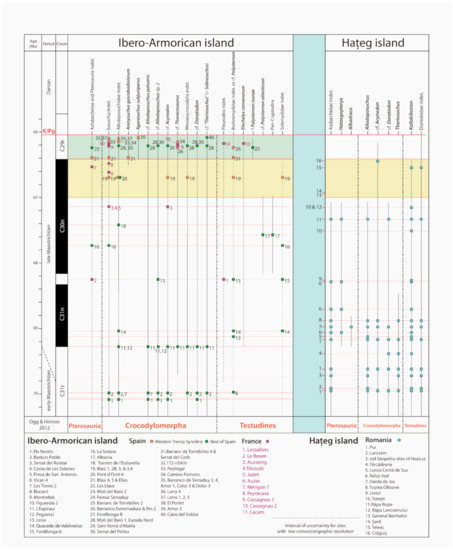
Figure 9. Pterosaur, crocodylomorph, and testudines groups and species occurrences in the Ibero-Armorican island and Haţeg island during the late Maastrichtian. Haţeg island data based on [32] with updated information. The yellow band marks the temporal interval used for comparison, whereas the green band marks the last ≈350 ka of the Maastrichtian, with a record only in the Ibero-Armorican island. Magnetochronostratigraphic scale based on [110] and calibration for the K-Pg boundary based on [28].
During the late Maastrichtian, the crocodylomorphs of the Ibero-Armorican island show great abundance, with a similar number of taxa to that during the early Maastrichtian [23]. The best-represented clade is the eusuchian Allodaposuchidae, with two taxa described from the Western Tremp Syncline (Agaresuchus subjuniperus and Arenysuchus), and probably Allodaposuchus palustris, whose characteristic teeth have been found up to chron C29r [87][114]. In addition, Blanco et al. (2020) [87] described Allodaposuchus sp. 2 on the basis of a dentary from the Fontllonga-6 site (C30r), which seems to be different from the allodaposuchids previously described and could represent a new taxon. Finally, there are plenty of isolated teeth of allodaposuchids [44][86][114][124] that due to their conical generalist shape are difficult to ascribe to specific taxa.
Gavialoidea is represented by a skull and other associated remains from the site of Cassagnau (Haute-Garonne, southern France). These have been ascribed to Thoracosaurus neocesariensis [125]. This assignment has been debated, as the remains could belong to a new taxon [126]. In addition, more teeth referred to cf. Thoracosaurus have been found in the Spanish Pyrenees, including the record of the Western Tremp Syncline [44][87].
The diversity of hylaeochampsids, “atoposaurids”, and notosuchians during the late Maastrichtian is difficult to determine, since most of their fossils are isolated teeth. There are several teeth referred to cf. Acynodon from France [127][128] and the Spanish Pyrenees, including the Western Tremp Syncline [87]. “Atoposaurids” are represented by teeth identified as cf. Theriosuchus; these are from the Spanish Pyrenees [87][114] but not the Western Tremp Syncline. “Atoposauridae” is here written in quotes, since Tennant et al. (2016) [129] have argued that some taxa assigned to this clade, such as “Theriosuchus” ibericus and “Theriosuchus” sympiestodon, belong to Paralligatoridae and have accordingly grouped these taxa under the new genus Sabresuchus. There are also some teeth from the Spanish Pyrenees identified as the notosuchian cf. Doratodon [87][114]. It should further be noted that plenty of undetermined eusuchian and crocodylomorph remains have been discovered in the French and Spanish Pyrenees (see [23] and references therein), as well as fossil tracks of crocodylomorphs [90], but due to their limited diagnostic value, it is difficult to ascertain their taxonomic status more precisely. There are also indeterminate eusuchian remains from La Solana (Valencia) [97] and Quecedo de Valdivielso (Burgos, NW Spain) [130][131]. Thus, Ibero-Armorican crocodylomorphs are represented during the late Maastrichtian by a minimum of eight taxa (Table 1, Figure 7 and Figure 9).
The record of testudines during the late Maastrichtian of the Ibero-Armorican island is poorer than during the early Maastrichtian. In the Pyrenees, pleurodiran turtles are represented by the bothremydid Elochelys convenarum Laurent, Tong, Claude [132] from southern France and another bothremydid turtle from Isona. This represents the species Polysternon isonae Marmi, Luján, Riera, Gaete, Oms, Galobart [133], although Pérez-García [134] considers this a nomen dubium, lacking enough diagnostic characters for a new species, and classifies the remains as Foxemydina indet. Isolated remains of indeterminate bothremydids are also present in other sites in the Pyrenees [93][135] and in the northwestern Spanish sites of Urria and Quecedo de Valdivielso (Burgos) [130][131]. In the fossil site of Albaina, there is a plate identified as cf. Polysternon atlanticum [109]. Pan-cryptodirans are represented by the remains of solemydid turtles from the Pyrenees, from the sites of Blasi and Fontllonga-6 [93][135], and from La Solana (Valencia) [97][98]. Pereda-Suberbiola et al. (2015) [109] describe a plate from a putative pan-cryptodiran that differs from solemydids. This makes a minimum of three pan-pleurodirans and two pan-cryptodirans in the Ibero-Armorican island during the late Maastrichtian (Table 1, Figure 7 and Figure 9).
Small-sized upper Maastrichtian tetrapods from the Ibero-Armorican island are represented only by amphibians and squamates from the Spanish and French Pyrenees [94][99][118][136] and from Valencia [137] (Figure 10). The first group consists of albanerpetontids, with at least one taxon present, identified in Blasi 2 as Albanerpeton aff. nexuosum [94], plus several albanerpetontid remains from the L’Espinau and Serrat del Rostiar 1 sites (Lleida, Catalonia, NE Spain) [136], Cassagnau 1 (Haute-Garonne, southern France) [99][118], and La Solana [137]. In La Solana, the presence of a salamandrid is also documented [137]. Anurans may be represented by at least four different groups with one discoglossid and one palaeobatrachid recognized at Blasi 2, L’Espinau, and Serrat del Rostiar [94][136], and an alytid and a putative pelobatid or gobiatid at L’Espinau [136]. It is noteworthy that there are remains of a palaeobatrachid from Valencia [137] that shows differences from the Blasi 2 taxon and could represent another taxon.

Figure 10. Squamate, amphibian and mammal groups and species occurrences in the Ibero-Armorican island and Haţeg island during the late Maastrichtian. Haţeg island data based on [32] with updated information. The yellow band marks the temporal interval used for comparison, whereas the green band marks the last ≈350 ka of the Maastrichtian, with a record only in the Ibero-Armorican island. Magnetochronostratigraphic scale based on [110] and calibration for the K-Pg boundary based on [28].
Upper Maastrichtian squamates are represented by the previously described fossils from Blasi 2 [94], with two undetermined lizards, one anguid lizard, and an alethinophid snake. Additionally, in the Pyrenees, the site of Serrat del Rostiar 1 (Lleida, Catalonia) has yielded several squamate remains [136] including geckos, anguid, and “scincomorph” lizards, and an indeterminate iguanid. An indeterminate iguanid can also be found at L’Espinau. The Serrat del Rostiar 1 site is dated to within chron C31r in the early Maastrichtian, but due to its stratigraphic position, it lies very close to the boundary with the late Maastrichtian, so we have extended its faunal assemblage to the lower part of the late Maastrichtian (Figure 10). In the French Pyrenees, there is also evidence of a large varanoid, “scincomorph” lizards, and other indeterminate squamates [99][118]. Outside the Pyrenees, there are also undetermined squamate remains at the La Solana site [97].
It is interesting to note that during the late Maastrichtian, there is almost no evidence of mammals in the Spanish record of the Ibero-Armorican island, despite the fact that their presence is documented during the early Maastrichtian [138][139][140] and the earliest Paleocene [141][142]. The only evidence of mammals during the late Maastrichtian is some tribosphenic teeth from the Peyrecave site, in the Petites Pyrénées (Haute-Garonne, southern France). These would have belonged to a therian mammal [118][143].
References
- Alvarez, L.W.; Alvarez, W.; Asaro, F.; Michel, H.V. Extraterrestrial cause for the cretaceous-tertiary extinction. Science 1980, 208, 1095–1108.
- Miller, K.G.; Kominz, M.A.; Browning, J.V.; Wright, J.D.; Mountain, G.S.; Katz, M.E.; Sugarman, P.J.; Cramer, B.S.; Christie-Blick, N.; Peka, S.F. The Phanerozoic Record of Global Sea-Level Change. Science 2005, 310, 1293–1298.
- Li, L.; Keller, G. Maastrichtian climate, productivity and faunal turnovers in planktic foraminifera in South Atlantic DSDP sites 525A and 21. Mar. Micropaleontol. 1998, 33, 55–86.
- Barnet, J.S.K.; Littler, K.; Kroon, D.; Leng, M.J.; Westerhold, T.; Röhl, U.; Zachos, J.C. A new high-resolution chronology for the late Maastrichtian warming event: Establishing robust temporal links with the onset of Deccan volcanism. Geology 2018, 46, 147–150.
- Courtillot, V.; Féraud, G.; Maluski, H.; Vandamme, D.; Moreau, M.G.; Besse, J. Deccan flood basalts and the Cretaceous/Tertiary boundary. Nature 1988, 333, 843–846.
- Courtillot, V.E.; Renne, P.R. On the ages of flood basalt events. Comptes Rendus Geosci. 2003, 335, 113–140.
- Tobin, T.S.; Ward, P.D.; Steig, E.J.; Olivero, E.B.; Hilburn, I.A.; Mitchell, R.N.; Diamond, M.R.; Raub, T.D.; Kirschvink, J.L. Extinction patterns, δ18 O trends, and magnetostratigraphy from a southern high-latitude Cretaceous–Paleogene section: Links with Deccan volcanism. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2012, 350–352, 180–188.
- Schoene, B.; Eddy, M.P.; Samperton, K.M.; Keller, C.B.; Keller, G.; Adatte, T.; Khadri, S.F.R. U-Pb constraints on pulsed eruption of the Deccan Traps across the end-Cretaceous mass extinction. Science 2019, 363, 862–866.
- Hildebrand, A.R.; Penfield, G.T.; Kring, D.A.; Pilkington, M.; Camargo, Z.A.; Jacobsen, S.B.; Boynton, W.V. Chicxulub Crater: A possible Cretaceous/Tertiary boundary impact crater on the Yucatán Peninsula, Mexico. Geology 1991, 19, 867.
- Renne, P.R.; Arenillas, I.; Arz, J.A.; Vajda, V.; Gilabert, V.; Bermúdez, H.D. Multi-proxy record of the Chicxulub impact at the Cretaceous-Paleogene boundary from Gorgonilla Island, Colombia. Geology 2018, 46, 547–550.
- Maruoka, T. Mass Extinction at the cretaceous–paleogene (K–Pg) boundary. In Astrobiology; Springer: Singapore, 2019; pp. 303–320.
- Lyons, S.L.; Karp, A.T.; Bralower, T.J.; Grice, K.; Schaefer, B.; Gulick, S.P.S.; Morgan, J.V.; Freeman, K.H. Organic matter from the Chicxulub crater exacerbated the K–Pg impact winter. Proc. Natl. Acad. Sci. USA 2020, 117, 25327–25334.
- Schulte, P.; Alegret, L.; Arenillas, I.; Arz, J.A.; Barton, P.J.; Bown, P.R.; Bralower, T.J.; Christeson, G.L.; Claeys, P.; Cockell, C.S.; et al. The chicxulub asteroid impact and mass extinction at the cretaceous-paleogene boundary. Science 2010, 327, 1214–1218.
- Witts, J.D.; Whittle, R.J.; Wignall, P.B.; Crame, J.A.; Francis, J.E.; Newton, R.J.; Bowman, V.C. Macrofossil evidence for a rapid and severe Cretaceous–Paleogene mass extinction in Antarctica. Nat. Commun. 2016, 7, 11738.
- Chiarenza, A.A.; Farnsworth, A.; Mannion, P.D.; Lunt, D.J.; Valdes, P.J.; Morgan, J.V.; Allison, P.A. Asteroid impact, not volcanism, caused the end-Cretaceous dinosaur extinction. Proc. Natl. Acad. Sci. USA 2020, 117, 17084–17093.
- Dzombak, R.M.; Sheldon, N.D.; Mohabey, D.M.; Samant, B. Stable climate in India during Deccan volcanism suggests limited influence on K–Pg extinction. Gondwana Res. 2020, 85, 19–31.
- Raup, D.M.; Sepkoski, J.J. Mass extinctions in the marine fossil record. Science 1982, 215, 1501–1503.
- Jablonski, D. Extinctions in the fossil record. Philos. Trans. R. Soc. London Ser. B Biol. Sci. 1994, 344, 11–17.
- Bardet, N. Extinction events among Mesozoic marine reptiles. Hist. Biol. 1994, 7, 313–324.
- Longrich, N.R.; Tokaryk, T.; Field, D.J. Mass extinction of birds at the Cretaceous-Paleogene (K-Pg) boundary. Proc. Natl. Acad. Sci. USA 2011, 108, 15253–15257.
- Longrich, N.R.; Martill, D.M.; Andres, B. Late Maastrichtian pterosaurs from North Africa and mass extinction of Pterosauria at the Cretaceous-Paleogene boundary. PLOS Biol. 2018, 16, e2001663.
- Brusatte, S.L.; Butler, R.J.; Barrett, P.M.; Carrano, M.T.; Evans, D.C.; Lloyd, G.T.; Mannion, P.D.; Norell, M.A.; Peppe, D.J.; Upchurch, P.; et al. The extinction of the dinosaurs. Biol. Rev. 2015, 90, 628–642.
- Puértolas-Pascual, E.; Blanco, A.; Brochu, C.A.; Canudo, J.I. Review of the late cretaceous-early paleogene crocodylomorphs of Europe: Extinction patterns across the K-PG boundary. Cretac. Res. 2016, 57, 565–590.
- Pearson, D.A.; Schaefer, T.; Johnson, K.R.; Nichols, D.J.; Hunter, J.P. Vertebrate biostratigraphy of the Hell Creek Formation in southwestern North Dakota and northwestern South Dakota. Spec. Pap. Geol. Soc. Am. 2002, 361, 145–167.
- Lyson, T.R.; Longrich, N.R. Spatial niche partitioning in dinosaurs from the latest Cretaceous (Maastrichtian) of North America. Proc. R. Soc. B Biol. Sci. 2011, 278, 1158–1164.
- Johnson, K.R.; Nichols, D.J.; Hartman, J.H. Hell Creek Formation: A 2001 synthesis. Geol. Soc. Am. Spec. Pap. 2002, 361, 503–510.
- LeCain, R.; Clyde, W.C.; Wilson, G.P.; Riedel, J. Magnetostratigraphy of the Hell Creek and lower Fort Union Formations in northeastern Montana. In Through the End of the Cretaceous in the Type Locality of the Hell Creek Formation in Montana and Adjacent Areas; Geological Society of America: Boulder, CO, USA, 2014.
- Sprain, C.J.; Renne, P.R.; Clemens, W.A.; Wilson, G.P. Calibration of chron C29r: New high-precision geochronologic and paleomagnetic constraints from the Hell Creek region, Montana. GSA Bull. 2018, 130, 1615–1644.
- Fowler, D. The Hell Creek Formation, Montana: A stratigraphic review and revision based on a sequence stratigraphic approach. Geosciences 2020, 10, 435.
- Csiki-Sava, Z.; Buffetaut, E.; Ősi, A.; Pereda-Suberbiola, X.; Brusatte, S.L. Island life in the Cretaceous—Faunal composition, biogeography, evolution, and extinction of land-living vertebrates on the Late Cretaceous European archipelago. Zookeys 2015, 469, 1–161.
- Canudo, J.I.; Oms, O.; Vila, B.; Galobart, À.; Fondevilla, V.; Puértolas-Pascual, E.; Sellés, A.G.; Cruzado-Caballero, P.; Dinarès-Turell, J.; Vicens, E.; et al. The upper Maastrichtian dinosaur fossil record from the southern Pyrenees and its contribution to the topic of the Cretaceous-Palaeogene mass extinction event. Cretac. Res. 2016, 57, 540–551.
- Csiki-Sava, Z.; Vremir, M.; Vasile, Ş.; Brusatte, S.L.; Dyke, G.; Naish, D.; Norell, M.A.; Totoianu, R. The east side story—The Transylvanian latest Cretaceous continental vertebrate record and its implications for understanding Cretaceous–Paleogene boundary events. Cretac. Res. 2016, 57, 662–698.
- Fondevilla, V.; Riera, V.; Vila, B.; Sellés, A.G.; Dinarès-Turell, J.; Vicens, E.; Gaete, R.; Oms, O. Galobart Chronostratigraphic synthesis of the latest Cretaceous dinosaur turnover in south-western Europe. Earth Sci. Rev. 2019, 191, 168–189.
- Vila, B.; Sellés, A.G.; Brusatte, S.L. Diversity and faunal changes in the latest Cretaceous dinosaur communities of southwestern Europe. Cretac. Res. 2016, 57, 552–564.
- Canudo, J.I. The Collection of Type Fossils of the Natural Science Museum of the University of Zaragoza (Spain). Geoheritage 2018, 10, 385–392.
- Pereda-Suberbiola, X.; Canudo, J.I.; Cruzado-Caballero, P.; Barco, J.L.; López-Martínez, N.; Oms, O.; Ruiz-Omeñaca, J.I. The last hadrosaurid dinosaurs of Europe: A new lambeosaurine from the Uppermost Cretaceous of Aren (Huesca, Spain). Comptes Rendus Palevol 2009, 8, 559–572.
- Pereda-Suberbiola, X.; Canudo, J.I.; Company, J.; Cruzado-Caballero, P.; Ruiz-Omeñaca, J.I. Hadrosauroid dinosaurs from the latest Cretaceous of the Iberian Peninsula. J. Vertebr. Paleontol. 2009, 29, 946–951.
- Cruzado-Caballero, P.; Ruiz-Omeñaca, J.I.; Canudo, J.I. Review of the fossill record of spanish hadrosaur remains. In Proceedings of the VIII Encuentro de Jóvenes Investigadores en Paleontología, Enciso, Spain, 21–22 April 2010; Volume 30, pp. 99–105.
- Cruzado-Caballero, P.; Puértolas-Pascual, E.; Canudo, J.I.; Castanera, D.; Gasca, J.M.; Moreno-Azanza, M. New hadrosaur remains from the Late Maastrichtian of Huesca (NE Spain). In Proceedings of the 10th Annual Meeting of the European Association of Vertebrate Palaeontologists, Teruel, Spain, 19–24 June 2012; pp. 45–48.
- Cruzado-Caballero, P.; Ruiz-Omeñaca, J.I.; Gaete, R.; Riera, V.; Oms, O.; Canudo, J.I. A new hadrosaurid dentary from the latest Maastrichtian of the Pyrenees (north Spain) and the high diversity of the duck-billed dinosaurs of the Ibero-Armorican Realm at the very end of the Cretaceous. Hist. Biol. 2014, 26, 619–630.
- Company, J.; Cruzado-Caballero, P.; Canudo, J.I. Presence of diminutive hadrosaurids (Dinosauria: Ornithopoda) in the Maastrichtian of the south-central Pyrenees (Spain). J. Iber. Geol. 2015, 41, 71–81.
- Prieto-Márquez, A.; Dalla Vecchia, F.M.; Gaete, R.; Galobart, À. Diversity, relationships, and biogeography of the lambeosaurine dinosaurs from the European archipelago, with description of the new aralosaurin Canardia garonnensis. PLoS ONE 2013, 8, e69835.
- Conti, S.; Vila, B.; Sellés, A.G.; Galobart, À.; Benton, M.J.; Prieto-Márquez, A. The oldest lambeosaurine dinosaur from Europe: Insights into the arrival of Tsintaosaurini. Cretac. Res. 2020, 107, 104286.
- Puértolas-Pascual, E.; Arenillas, I.; Arz, J.A.; Calvín, P.; Ezquerro, L.; García-Vicente, C.; Pérez-Pueyo, M.; Sánchez-Moreno, E.M.; Villalaín, J.J.; Canudo, J.I. Chronostratigraphy and new vertebrate sites from the upper Maastrichtian of Huesca (Spain), and their relation with the K/Pg boundary. Cretac. Res. 2018, 89, 36–59.
- López-Martínez, N.; Canudo, J.I.; Ardevol, L.; Pereda-Suberbiola, X.; Orue-Etxebarria, X.; Cuenca-Bescós, G.; Ruiz-Omeñaca, J.I.; Murelaga, X.; Feist, M. New dinosaur sites correlated with upper Maastrichtian pelagic deposits in the Spanish Pyrenees: Implications for the dinosaur extinctio. Cretac. Res. 2001, 22, 41–61.
- Cruzado-Caballero, P.; Pereda-Suberbiola, X.; Ruiz-Omeñaca, J.I. Blasisaurus canudoi gen. et sp. nov., a new lambeosaurine dinosaur (hadrosauridae) from the latest cretaceous of Arén (Huesca, Spain). Can. J. Earth Sci. 2010, 47, 1507–1517.
- Cruzado-Caballero, P.; Canudo, J.I.; Moreno-Azanza, M.; Ruiz-Omeñaca, J.I. New material and phylogenetic position of Arenysaurus ardevoli, a lambeosaurine dinosaur from the late Maastrichtian of Arén (Northern Spain). J. Vertebr. Paleontol. 2013, 33, 1367–1384.
- Longrich, N.R.; Suberbiola, X.P.; Pyron, R.A.; Jalil, N.-E. The first duckbill dinosaur (Hadrosauridae: Lambeosaurinae) from Africa and the role of oceanic dispersal in dinosaur biogeography. Cretac. Res. 2020, 104678.
- Cruzado-Caballero, P.; Canudo Sanagustín, J.; Ruiz-Omeñaca, J.I. Nuevas evidencias de la presencia de hadrosaurios lambeosaurios (Dinosauria) en el Maastrichtiense superior de la Península Ibérica (Arén, Huesca). Geogaceta 2005, 3, 47–50.
- Cruzado-Caballero, P.; Canudo, J.I.; Ruiz-Omeñaca, J.I. Los fémures de Blasi (Arén, Huesca, Spain): Una contribución a los hadrosauroideos Europeos del Maastrichtiense superior. In Proceedings of the Actas de las IV Jornadas Internacionales sobre Paleontologıa de Dinosaurios y su Entorno, Burgos, Spain, 5–9 September 2009; pp. 197–205.
- Cruzado-Caballero, P.; Ruiz-Omeñaca, J.I.; Canudo, J.I. Evidencias de la coexistencia de dinosaurios hadrosaurinos y lambeosaurinos en el Maastrichtiano superior de la Península Ibérica (Arén, Huesca, España). Ameghiniana 2010, 47, 153–164.
- Canudo, I.; Cruzado-Caballero, P.; Moreno-Azanza, M. Possible theropod predation evidence in hadrosaurid dinosaurs from the Upper Maastrichtian (Upper Cretaceous) of Arén (Huesca, Spain). Kaupia Darmstädter Beiträge zur Naturgeschichte 2005, 14, 9–13.
- Cruzado-Caballero, P.; Fortuny, J.; Llacer, S.; Canudo, J.I. Paleoneuroanatomy of the European lambeosaurine dinosaur Arenysaurus ardevoli. Peer J. 2015, 2015, 1–16.
- Mayayo-Lainez, A. Paleohistología y Mineralogía de los Dinosaurios Hadrosaurios del Maastrichtiense de Arén (Huesca). Master’s Thesis, University of Zaragoza, Zaragoza, Spain, 2020.
- Mayayo-Lainez, A.; Alegre-Esteve, M.; Bauluz, B.; Canudo, J. First approach to the paleohistology of the hadrosaur dinosaurs from Blasi 2A (Tremp Formation, Maastrichtian, Huesca). Ciências Terra-Procedia 2021, 1, 38–41.
- Puértolas-Pascual, E.; Cruzado-Caballero, P.; Canudo, J.I.; Gasca, J.M.; Moreno-Azanza, M.; Castanera, D.; Parrillas, J.; Ezquerro, L. Nuevos yacimientos de vertebrados del Maastrichtiense superior (Cretácico Superior) de Huesca (España). In Proceedings of the VIII Geological Congress of Spain, Oviedo, Spain, 17–19 July 2012; pp. 269–272.
- Pérez Pueyo, M.; Puértolas-Pascual, E.; Bádenas, B. Larra 4: Desenterrando a los últimos vertebrados del maastrichtiense terminal del pirineo aragonés. Zubía 2019, 31, 159–163.
- Pérez-Pueyo, M.; Gilabert, V.; Moreno-Azanza, M.; Puertolas-Pascual, E.; Bádenas, B.; Canudo, J.I. Late Maastrichtian fossil assemblage of Veracruz 1 site (Beranuy, NE Spain): Wildfires and bones in a transitional environment. In Proceedings of the VIII Jornadas Internacionales sobre Paleontología de Dinosaurios y su Entorno, Burgos, Spain, 5–9 September 2019; pp. 111–113.
- Barco, J.L.; Ardevol, L.; Canudo, J.I. Descripción de los primeros rastros asignados a Hadrosauridae (Ornithopoda, Dinosauria) del Maastrichtiense de la Península Ibérica (Areny, Huesca). Geogaceta 2001, 30, 235–238.
- Vila, B.; Oms, O.; Fondevilla, V.; Gaete, R.; Galobart, À.; Riera, V.; Canudo, J.I. The Latest Succession of Dinosaur Tracksites in Europe: Hadrosaur Ichnology, Track Production and Palaeoenvironments. PLoS ONE 2013, 8, e72579.
- Sellés, A.G.; Via, B.; Galobart, À. Spheroolithus europaeus, oosp. nov. (late Maastrichtian, Catalonia), the youngest oological record of hadrosauroids in Eurasia. J. Vertebr. Paleontol. 2014, 34, 725–729.
- Pérez-Pueyo, M.; Moreno-Azanza, M.; Núñez-Lahuerta, C.; Puértolas-Pascual, E.; Bádenas, B.; Canudo, J.I. Eggshell association of the Late Maastrichtian (Late Cretaceous) at Blasi 2B fossil site: A scrambled of vertebrate diversity. Ciências da Terra-Procedia 2021, 1, 58–61.
- Vila, B.; Galobart, À.; Canudo, J.I.; Le Loeuff, J.; Dinarès-Turell, J.; Riera, V.; Oms, O.; Tortosa, T.; Gaete, R. The diversity of sauropod dinosaurs and their first taxonomic succession from the latest Cretaceous of southwestern Europe: Clues to demise and extinction. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2012, 350–352, 19–38.
- Vila, B.; Sellés, A.G. Re-evaluation of the age of some dinosaur localities from the southern Pyrenees by means of megaloolithid oospecies. J. Iber. Geol. 2015, 41.
- Canudo, J.I. Descripcion de un fragmento proximal de fémur de Titanosauridae (Dinosauria, Sauropoda) del Maastrichtiense superior de Serraduy (Huesca). In Proceedings of the XVII Jornadas de la Sociedad Española de Paleontología, Albarracín, Spain, 18–20 October 2001; pp. 255–262.
- Sellés, A.G.; Marmi, J.; Llácer, S.; Blanco, A. The youngest sauropod evidence in Europe. Hist. Biol. 2016, 28, 930–940.
- Fondevilla, V.; Vila, B.; Oms, O.; Galobarp, À. Skin impressions of the last European dinosaurs. Geol. Mag. 2017, 154, 393–398.
- Díez Díaz, V.; Pereda Suberbiola, X.; Sanz, J.L. The axial skeleton of the titanosaur Lirainosaurus astibiae (Dinosauria: Sauropoda) from the latest Cretaceous of Spain. Cretac. Res. 2013, 43, 145–160.
- Salgado, L.; Coria, R.A.; Calvo, J.O. Evolution of titanosaurid sauropods: Phylogenetic analysis based on the postcranial evidence. Ameghiniana 1997, 34, 3–32.
- Upchurch, P.; Barrett, P.M.; Dodson, P. 13. Sauropoda. In The Dinosauria, 2nd ed.; University of California Press: Berkeley, CA, USA, 2019; pp. 259–322.
- Calvo, J.O.; Bonaparte, J.F. Andesaurus delgadoi gen. et. sp. nov. (Saurischia-Sauropoda), dinosaurio Titanosauridae de la Formacion Rio Limay (Albiano-Cenomaniano), Neuquen, Argentina. Ameghiniana 1991, 28, 303–310.
- Mannion, P.D.; Calvo, J.O. Anatomy of the basal titanosaur (Dinosauria, Sauropoda) Andesaurus delgadoi from the mid-Cretaceous (Albian-early Cenomanian) Río Limay Formation, Neuquén Province, Argentina: Implications for titanosaur systematics. Zool. J. Linn. Soc. 2011, 155–181.
- Torices, A.; Currie, P.J.; Canudo, J.I.; Pereda-Suberbiola, X. Theropod dinosaurs from the upper cretaceous of the south pyrenees basin of Spain. Acta Palaeontol. Pol. 2015, 60, 611–626.
- Pérez-García, A.; Ortega, F.; Bolet, A.; Escaso, F.; Houssaye, A.; Martínez-Salanova, J.; de Miguel Chaves, C.; Mocho, P.; Narváez, I.; Segura, M.; et al. A review of the upper Campanian vertebrate site of Armuña (Segovia Province, Spain). Cretac. Res. 2016, 57, 591–623.
- Pérez-Pueyo, M.; Puértolas-Pascual, E.; Moreno-Azanza, M.; Cruzado-Caballero, P.; Gasca, J.M.; Núñez-Lahuerta, C.; Canudo, J.I. First record of a giant bird (Ornithuromorpha) from the uppermost Maastrichtian of the Southern Pyrenees, NE Spain. J. Vertebr. Paleontol. 2021, 41, e1900210.
- López-Martínez, N.; Canudo, J.I.; Cuenca-Bescós, G. Latest cretaceous eggshells from Arén (Southern Pyrenees, Spain). In Proceedings of the First International Symposium on Dinosaur Eggs and Babies, Isona, Spain, 23–26 September 1999; pp. 35–36.
- Núñez-Lahuerta, C.; Moreno-Azanza, M.; Pérez-Pueyo, M. First approach for a taphonomic key for fossil eggs and eggshells accumulations using optic microscopy: The case of Blasi-2B (Upper Cretaceous, Spain). Ciências Terra Procedia 2021, 1, 42–45.
- Choi, S.; Moreno-Azanza, M.; Csiki-Sava, Z.; Prondvai, E.; Lee, Y.N. Comparative crystallography suggests maniraptoran theropod affinities for latest Cretaceous European ‘geckoid’ eggshell. Pap. Palaeontol. 2020, 6, 265–292.
- Dalla Vecchia, F.M.; Riera, F.M.; Oms, J.O.; Dinarès-Turell, J.; Gaete, R.; Galobart, A. The last pterosaurs: First record from the uppermost Maastrichtian of the Tremp Syncline (Northern Spain). Acta Geol. Sin. 2013, 87, 1198–1227.
- Puértolas, E.; Canudo, J.I.; Cruzado-Caballero, P. A new crocodylian from the late Maastrichtian of Spain: Implications for the initial radiation of crocodyloids. PLoS ONE 2011, 6.
- Blanco, A.; Puértolas-Pascual, E.; Marmi, J.; Vila, B.; Sellés, A.G. Allodaposuchus palustris sp. nov. from the Upper Cretaceous of Fumanya (South-Eastern Pyrenees, Iberian Peninsula): Systematics, Palaeoecology and Palaeobiogeography of the Enigmatic Allodaposuchian Crocodylians. PLoS ONE 2014, 9, e115837.
- Blanco, A.; Fortuny, J.; Vicente, A.; Luján, À.H.; García-Marçà, J.A.; Sellés, A.G. A new species of Allodaposuchus (Eusuchia, Crocodylia) from the Maastrichtian (Late Cretaceous) of Spain: Phylogenetic and paleobiological implications. Peer J. 2015, 3, e1171.
- Narváez, I.; Brochu, C.A.; Escaso, F.; Pérez-García, A.; Ortega, F. New Crocodyliforms from Southwestern Europe and Definition of a Diverse Clade of European Late Cretaceous Basal Eusuchians. PLoS ONE 2015, 10, e0140679.
- Narváez, I.; Brochu, C.A.; Escaso, F.; Pérez-García, A.; Ortega, F. New Spanish Late Cretaceous eusuchian reveals the synchronic and sympatric presence of two allodaposuchids. Cretac. Res. 2016, 65, 112–125.
- Mateus, O.; Puértolas-Pascual, E.; Callapez, P.M. A new eusuchian crocodylomorph from the Cenomanian (Late Cretaceous) of Portugal reveals novel implications on the origin of Crocodylia. Zool. J. Linn. Soc. 2019, 186, 501–528.
- Puértolas-Pascual, E.; Canudo, J.I.; Moreno-Azanza, M. The eusuchian crocodylomorph Allodaposuchus subjuniperus sp. nov., a new species from the latest Cretaceous (upper Maastrichtian) of Spain. Hist. Biol. 2014, 26, 91–109.
- Blanco, A.; Puértolas-Pascual, E.; Marmi, J.; Moncunill-Solé, B.; Llácer, S.; Rössner, G.E. Late Cretaceous (Maastrichtian) crocodyliforms from north-eastern Iberia: A first attempt to explain the crocodyliform diversity based on tooth qualitative traits. Zool. J. Linn. Soc. 2020, 189, 584–617.
- Moreno-Azanza, M.; Bauluz, B.; Canudo, J.I.; Puértolas-Pascual, E.; Sellés, A.G. A re-evaluation of aff. Megaloolithidae eggshell fragments from the uppermost Cretaceous of the Pyrenees and implications for crocodylomorph eggshell structure. Hist. Biol. 2014, 26, 195–205.
- Hirsch, K.F. Fossil crocodilian eggs from the eocene of Colorado. J. Paleontol. 1985, 3, 531–542.
- Vila, B.; Castanera, D.; Marmi, J.; Canudo, J.I.; Galobart, À. Crocodile swim tracks from the latest Cretaceous of Europe. Lethaia 2015, 48, 256–266.
- Pérez-Pueyo, M.; Castanera, D.; Bádenas, B.; Canudo, J.I. New evidences of Crocodylomorpha swim tracks in the Maastrichtian of Beranuy (Huesca, Spain). In Proceedings of the XVI Encuentro de Jóvenes Investigadores en Paleontología, Zarautz, Spain, 11–14 April 2018; pp. 137–140.
- De Lapparent de Broin, F.; Murelaga, X. Turtles from Upper Cretaceous of Lano (Iberian Peninsula). Comptes Rendus l’Académie Sci. 1996, 323, 729–735.
- Murelaga, X.; Canudo, J.I. Descripción de los restos de quelonios del Maastrichtiense superior de Aren y Serraduy (Huesca). Geogaceta 2005, 28, 51–54.
- Blain, H.A.; Canudo, J.I.; Cuenca-Bescós, G.; López-Martínez, N. Amphibians and squamate reptiles from the latest Maastrichtian (Upper Cretaceous) of Blasi 2 (Huesca, Spain). Cretac. Res. 2010, 31, 433–446.
- Berreteaga, A.; Pereda Suberbiola, X.; Floquet, M.; Olivares, M.; Etxebarria, N.; Iriarte, E.; Badiola Kortabitarte, A.; Elorza, J.; Astibia Ayerra, H. Datos sedimentológicos y tafonómicos de enclaves finicretácicos con fósiles de vertebrados de la Formación Sobrepeña (Burgos, Región Vasco-Cantábrica). Geo-Temas 2008, 10, 1278–1280.
- Corral, J.-C.; Pueyo, E.L.; Berreteaga, A.; Rodríguez-Pintó, A.; Sánchez, E.; Pereda-Suberbiola, X. Magnetostratigraphy and lithostratigraphy of the Laño vertebrate-site: Implications in the uppermost Cretaceous chronostratigraphy of the Basque-Cantabrian Region. Cretac. Res. 2016, 57, 473–489.
- Company, J. Vertebrados continentales del Cretácico superior (Campaniense- Maatrichtiense) de Valencia. Ph.D. Thesis, Universidad de Valencia, Valencia, Spain, 2004.
- Company, J.; Pereda-Suberbiola, X.; Ruiz-Omeñaca, J.I. Last Cretaceous dinosaur faunas from Eastern Iberia into its regional paleogeographic context. Faunal composition and palaeobiogeographical implications. In Proceedings of the IV Jornadas Internacionales sobre Paleontología de Dinosaurios y su Entorno Salas de los Infantes, Burgos, Spain, 5–9 September 2009; pp. 17–44.
- Laurent, Y.; Bilotte, M.; Le Loeuff, J. Late Maastrichtian continental vertebrates from southwestern France: Correlation with marine fauna. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2002, 187, 121–135.
- Le Loeuff, J.; Buffetaut, E.; Martin, M. The last stages of dinosaur faunal history in Europe: A succession of Maastrichtian dinosaur assemblages from the Corbières (southern France). Geol. Mag. 1994, 131, 625–630.
- Prieto-Márquez, A.; Fondevilla, V.; Sellés, A.G.; Wagner, J.R.; Galobart, À. Adynomosaurus arcanus, a new lambeosaurine dinosaur from the Late Cretaceous Ibero-Armorican Island of the European archipelago. Cretac. Res. 2019, 96, 19–37.
- Casanovas-Cladellas, M.L.; Santafé-Llopis, J.V.; Isidro-Llorens, A. Pararhabdodon isonensis n. gen. n. sp. (Dinosauria). Estudio mofológico, radio-tomográfico y consideraciones biomecanicas 26-27:121-131. Paleontol. Evol. 1993, 26–27, 121–132.
- Serrano, J.F.; Sellés, A.G.; Vila, B.; Galobart, À.; Prieto-Márquez, A. The osteohistology of new remains of Pararhabdodon isonensis sheds light into the life history and paleoecology of this enigmatic European lambeosaurine dinosaur. Cretac. Res. 2021, 118, 104677.
- Prieto-Marquez, A.; Wagner, J.R. Pararhabdodon isonensis and Tsintaosaurus spinorhinus: A new clade of lambeosaurine hadrosaurids from Eurasia. Cretac. Res. 2009, 30, 1238–1246.
- Prieto-Marquez, A.; Gaete, R.; Rivas, G.; Galobart, À.; Boada, M. Hadrosauroid dinosaurs from the Late Cretaceous of Spain: Pararhabdodon isonensis revisited and Koutalisaurus kohlerorum, gen. et sp. nov. J. Vertebr. Paleontol. 2006, 26, 929–943.
- Blanco, A.; Prieto-Márquez, A.; De Esteban-Trivigno, S. Diversity of hadrosauroid dinosaurs from the Late Cretaceous Ibero-Armorican Island (European Archipelago) assessed from dentary morphology. Cretac. Res. 2015, 56, 447–457.
- Fondevilla, V.; Dalla Vecchia, F.M.; Gaete, R.; Galobart, À.; Moncunill-Solé, B.; Köhler, M. Ontogeny and taxonomy of the hadrosaur (Dinosauria, Ornithopoda) remains from Basturs Poble bonebed (late early Maastrichtian, Tremp Syncline, Spain). PLoS ONE 2018, 13, e0206287.
- Company, J.; Galobart Lorente, À.; Gaete, R. First data on the hadrosaurid dinosaurs (Ornithischia, Dinosauria) from the Upper Cretaceous of Valencia, Spain. Oryctos 1998, 1, 121–126.
- Pereda-Suberbiola, X.; Pérez-García, A.; Corral, J.C.; Murelaga, X.; Martin, G.; Larrañaga, J.; Bardet, N.; Berreteaga, A.; Company, J. First dinosaur and turtle remains from the latest Cretaceous shallow marine deposits of Albaina (Laño quarry, Iberian Peninsula). Comptes Rendus Palevol. 2015, 14, 471–482.
- Ogg, J.G.; Hinnov, L.A.; Huang, C. Cretaceous. In The Geologic Time Scale; Elsevier: Amsterdam, The Netherlands, 2012; pp. 793–853.
- Santafé, J.V.; Casanovas, M.L.; Llompart, C. Els Dinosaures i el Seu Entorn Geològic; Diputació de Lleida: Lleida, Spain, 1997.
- López-Martínez, N.; Moratalla, J.J.; Sanz, J.L. Dinosaurs nesting on tidal flats. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2000, 160, 153–163.
- Baiano, M.; Galobart, À.; Dalla, F.M.; Vila, B. Aplicación de la microtomografía en el estudio de dientes aislados de dinosaurios terópodos. In Proceedings of the XII Encuentro de Jóvenes investigadores en Paleontología, Boltaña, Spain, 9–12 April 2014; pp. 57–59.
- Marmi, J.; Blanco, A.; Fondevilla, V.; Dalla Vecchia, F.M.; Sellés, A.G.; Vicente, A.; Martín-Closas, C.; Oms, O.; Galobart, À. The Molí del Baró-1 site, a diverse fossil assemblage from the uppermost Maastrichtian of the southern Pyrenees (north-eastern Iberia). Cretac. Res. 2016, 57, 519–539.
- Sellés, A.G.; Vila, B.; Brusatte, S.L.; Currie, P.J.; Galobart, À. A fast-growing basal troodontid (Dinosauria: Theropoda) from the latest Cretaceous of Europe. Sci. Rep. 2021, 11, 4855.
- Sellés, A.G.; Vila, B.; Galobart, À. Diversity of theropod ootaxa and its implications for the latest cretaceous dinosaur turnover in southwestern Europe. Cretac. Res. 2014, 49, 45–54.
- Vianey-Liaud, M.; Lopez-Martinez, N. Late Cretaceous Dinosaur Eggshells from the Tremp Basin, Southern Pyrenees, Lleida, Spain. J. Paleontol. 1997, 71, 1157–1171.
- Laurent, Y. Les faunes de vertébrés continentaux du Maastrichtien supérieur d’Europe: Systématique et biodiversité. Strat Série 2003, 41, 1–81.
- Buffetaut, E.; Clarke, J.B.; Le Loeuff, J. A terminal Cretaceous pterosaur from the Corbieres (southem France) and the problem of pterosaur extinction. Bull. Soc. Geol. Fr. 1996, 167, 753–759.
- Buffetaut, E.; Laurent, Y.; Le Loeuff, J.; Bilotte, M. A terminal Cretaceous giant pterosaur from the French Pyrenees. Geol. Mag. 1997, 134, 553–556.
- Buffetaut, E. Late Cretaceous pterosaurs from France: A review. Zitteliana 2008, 28, 249–255.
- Company, J.; Ruiz-Omeñaca, J.I.; Suberbiola, X.P. A long-necked pterosaur (Pterodactyloidea, Azhdarchidae) from the Upper Cretaceous of Valencia, Spain. Geol. Mijnbouw/Netherlands J. Geosci. 1999, 78, 319–333.
- Pereda-Suberbiola, X.; Company, J.; Ruiz-Omeñaca, J.I. Azhdarchid pterosaurs from the Late Cretaceous (Campanian-Maastrichtian) of the Iberian Peninsula. In Proceedings of the Munich Flugsaurier, The Wellnhofer Pterosaur Meeting, Munich, Germany, 10–14 September 2007; p. 27.
- Blanco, A.; Méndez, J.M.; Marmi, J. The fossil record of the uppermost Maastrichtian Reptile Sandstone (Tremp Formation, northeastern Iberian Peninsula). Span. J. Paleontol. 2015, 30, 147–160.
- Laurent, Y.; Buffetaut, E.; Le Loeuff, J. Un crane de thoracosaurine (Crocodylia, Crocodylidae) dans le Maastrichtien Supérieur du sud de la France. Oryctos 2000, 3, 19–27.
- Brochu, C.A. A new Late Cretaceous gavialoid crocodylian from eastern North America and the phylogenetic relationships of thoracosaurs. J. Vertebr. Paleontol. 2004, 24, 610–633.
- Le Loeuff, J.; Buffetaut, E.; Cavin, L.; Laurent, Y.; Martin, M.; Martin, V.; Tong, H. Les hadrosaures des Corbières et des Petites Pyrenées. Bull. Soc. Etudes Sci. Aude 1994, 94, 19–21.
- Buffetaut, E.; Le Loeuff, J. Late Cretaceous dinosaurs from the foothills of the Pyrenees. Geol. Today 1997, 13, 60–68.
- Tennant, J.P.; Mannion, P.D.; Upchurch, P. Evolutionary relationships and systematics of Atoposauridae (Crocodylomorpha: Neosuchia): Implications for the rise of Eusuchia. Zool. J. Linn. Soc. 2016, 177, 854–936.
- Murelaga, X.; García Garmilla, F.; Pereda Suberbiola, X. Primeros restos de vertebrados del Cretácico Superior de Quecedo de Valdivieso (Burgos). Geogaceta 2005, 195–198.
- Berreteaga, A. Estudio Estratigráfico, Sedimentológico y Paleontológico de los Yacimientos con Fósiles de Vertebrados del Cretácico Final de la Región Vasco-Cantábrica. Ph.D. Thesis, University of the Basque Country, Bilbao, Spain, 2008.
- Laurent, Y.; Tong, H.; Claude, J. New side-necked turtle (Pleurodira: Bothremydidae) from the Upper Maastrichtian of the Petites-Pyrénées (Haute-Garonne, France). Cretac. Res. 2002, 23, 465–471.
- Marmi, J.; Luján, Á.H.; Riera, V.; Gaete, R.; Oms, O.; Galobart, À. The youngest species of Polysternon: A new bothremydid turtle from the uppermost Maastrichtian of the southern Pyrenees. Cretac. Res. 2012, 35, 133–142.
- Pérez-García, A. Las tortugas mesozoicas de la Peninsula Ibérica. Ph.D. Thesis, Universidad Complutense de Madrid, Madrid, Spain, 2012.
- Murelaga, X.; Pereda Suberbiola, X.; Astibia, H.; Lapparent, F.D. Primeros datos sobre los quelonios del Cretácico superior de Lleida. Geogaceta 1998, 24, 239–242.
- Blanco, A.; Bolet, A.; Blain, H.-A.; Fondevilla, V.; Marmi, J. Late Cretaceous (Maastrichtian) amphibians and squamates from northeastern Iberia. Cretac. Res. 2016, 57, 624–638.
- Szentesi, Z.; Company, J. Late Maastrichtian small-sized herpetofauna from Valencia province, eastern Spain. Hist. Biol. 2017, 29, 43–52.
- Pol, C.; Buscalioni, A.D.; Carballeira, J.; Francés, V.; López-Martínez, N.; Marandat, B.; Moratalla, J.; Sanz, J.L.; Sigé, B.; Villatte, J. Reptiles and mammals from the Late Cretaceous new locality Quintanilla del Coco (Burgos Province, Spain). Neues Jahrb. Geol. Paläontol. Abh. 1992, 184, 279–314.
- Gheerbrant, E.; Astibia, H. Addition to the Late Cretaceous Laño mammal faunule (Spain) and to the knowledge of European “Zhelestidae” (Lainodontinae nov.). Bull. Soc. Géol. Fr. 2012, 183, 537–546.
- Tabuce, R.; Tortosa, T.; Vianey-Liaud, M.; Garcia, G.; Lebrun, R.; Godefroit, P.; Dutour, Y.; Berton, S.; Valentin, X.; Cheylan, G. New eutherian mammals from the Late Cretaceous of Aix-en-Provence Basin, south-eastern France. Zool. J. Linn. Soc. 2013, 169, 653–672.
- Peláez-Campomanes, P.; López-Martínez, N.; Álvarez-Sierra, M.A.; Daams, R. The earliest mammal of the European Paleocene: The multituberculate Hainina. J. Paleontol. 2000, 74, 701–711.
- López-Martínez, N.; Pelaez-Campomanes, P. New mammals from south-central Pyrenees (Tremp Formation, Spain) and their bearing on late Paleocene marine-continental correlations. Bull. Soc. Géol. Fr. 1999, 170, 681–696.
- Gheerbrant, E.; Abrial, C.; Cappetta, H. Nouveaux sites a microvertébrés continentaux du Crétacé terminal des Petites Pyrénées (Haute-Garonne et Ariège, France). Geobios 1997, 30, 257–269.