Netrin-1 is one of the most well-studied proteins that regulate axonal guidance and synaptogenesis via interaction with its receptors deleted in colorectal cancer (DCC) and UNC5H (unco-ordinated-5 homolog) to activate cell survival, differentiation and proliferation.
- intracerebral hemorrhage
- hemin
- neuroprotection
- Netrin-1
- peptide
1. Introduction
Intracerebral hemorrhage (ICH) is a devastating stroke subtype with a high mortality and morbidity rate [1,2,3] that involves abnormal rupture of cerebral blood vessels within the brain [4,5]. Surgical removal of hematomas is preferred for achieving hemostasis and relieving intracranial pressure [6]. However, remnant hematic residues and blood breakdown products can still promote secondary brain injury following ICH, inducing neuronal damage and delaying functional recovery [7,8,9]. Thus, the development of effective strategies targeting secondary brain injury induced neuronal damage would be beneficial for stroke treatment.
Intracerebral hemorrhage (ICH) is a devastating stroke subtype with a high mortality and morbidity rate [1][2][3] that involves abnormal rupture of cerebral blood vessels within the brain [4][5]. Surgical removal of hematomas is preferred for achieving hemostasis and relieving intracranial pressure [6]. However, remnant hematic residues and blood breakdown products can still promote secondary brain injury following ICH, inducing neuronal damage and delaying functional recovery [7][8][9]. Thus, the development of effective strategies targeting secondary brain injury induced neuronal damage would be beneficial for stroke treatment.
Netrin-1 is one of the most well-studied proteins that regulate axonal guidance and synaptogenesis [10,11,12] via interaction with its receptors deleted in colorectal cancer (DCC) and UNC5H (unco-ordinated-5 homolog) to activate cell survival, differentiation and proliferation [13,14,15]. The receptors induce a death signal to mediate apoptosis when the Netrin-1 is absent [16]. It is known that the binding of Netrin-1 with DCC results in dimerization of DCC and initiates tyrosine 861 phosphorylation of focal adhesion kinase (FAK) to induce neurite outgrowth and axonal guidance [17,18]. The binding of Netrin-1 with DCC involves the EGF3 domain of Netrin-1 and the FN5 domain of DCC in activation of Netrin-1 signaling pathway [19,20]. Recently, Netrin-1 was shown to exert neuroprotective effects in ischemic regions [21]. The expression level of Netrin-1, which inhibits cell apoptosis and promotes neuronal regeneration, in the serum is positively correlated with patient recovery from ischemic stroke [22,23].
Netrin-1 is one of the most well-studied proteins that regulate axonal guidance and synaptogenesis [10][11][12] via interaction with its receptors deleted in colorectal cancer (DCC) and UNC5H (unco-ordinated-5 homolog) to activate cell survival, differentiation and proliferation [13][14][15]. The receptors induce a death signal to mediate apoptosis when the Netrin-1 is absent [16]. It is known that the binding of Netrin-1 with DCC results in dimerization of DCC and initiates tyrosine 861 phosphorylation of focal adhesion kinase (FAK) to induce neurite outgrowth and axonal guidance [17][18]. The binding of Netrin-1 with DCC involves the EGF3 domain of Netrin-1 and the FN5 domain of DCC in activation of Netrin-1 signaling pathway [19][20]. Recently, Netrin-1 was shown to exert neuroprotective effects in ischemic regions [21]. The expression level of Netrin-1, which inhibits cell apoptosis and promotes neuronal regeneration, in the serum is positively correlated with patient recovery from ischemic stroke [22][23].
Interestingly, transplantation of bone marrow mesenchymal stem cells that secrete Netrin-1 into the damaged rat sciatic nerve initiates axonal regeneration [24] and reduces motor neuron death [25]. Stereotaxic injection of the Netrin-1 protein into the ventricles reduces nerve cell death after subarachnoid hemorrhage through activation of DCC/APPL-1/AKT signaling [26]. A polypeptide derived from the EGF3 domain of Netrin-1 (amino acids 423–433) markedly activates ERK phosphorylation and promotes the production of NO, which has cardioprotective effects [27,28]. However, Netrin-1 has been identified as a potential biomarker for tumorigenesis [29,30,31] because regulation of the Netrin-1 status is found in multiple tissue-derived cancers [32]. When Netrin-1 is bound to its receptor DCC or Unc5H, the receptors transduce a positive signal leading to survival, inflammation, angiogenesis and anti-apoptosis, which in turn regulates tumorigenesis [31]. Therefore, it is very important to identify the minimal functional domain of Netrin-1 that can preserve the neuroprotection and avoid the major side effects of Netrin-1.
Interestingly, transplantation of bone marrow mesenchymal stem cells that secrete Netrin-1 into the damaged rat sciatic nerve initiates axonal regeneration [24] and reduces motor neuron death [25]. Stereotaxic injection of the Netrin-1 protein into the ventricles reduces nerve cell death after subarachnoid hemorrhage through activation of DCC/APPL-1/AKT signaling [26]. A polypeptide derived from the EGF3 domain of Netrin-1 (amino acids 423–433) markedly activates ERK phosphorylation and promotes the production of NO, which has cardioprotective effects [27][28]. However, Netrin-1 has been identified as a potential biomarker for tumorigenesis [29][30][31] because regulation of the Netrin-1 status is found in multiple tissue-derived cancers [32]. When Netrin-1 is bound to its receptor DCC or Unc5H, the receptors transduce a positive signal leading to survival, inflammation, angiogenesis and anti-apoptosis, which in turn regulates tumorigenesis [31]. Therefore, it is very important to identify the minimal functional domain of Netrin-1 that can preserve the neuroprotection and avoid the major side effects of Netrin-1.
Here, we found that peptide E1 (residues 407–422), which is derived from the EGF3 domain of Netrin-1, interacts with DCC to protect neurons from hemin-induced toxicity. In an experimental model of ICH mice, the application of peptide E1 improved functional recovery, as determined by behavioral assays. Furthermore, peptide E1 can prevent neurons from degenerating after ICH. On the basis of these results, we identified a functional sequence of Netrin-1 that is involved in recovery after ICH and determined that this peptide may be used for the treatment of ICH.
2. EGF3 Domain Is Critical for the Netrin-DCC Interaction
Netrin-1 consists of a VI domain, three EGF repeats, and a
C-terminal domain [33]. The crystal structure of the human Netrin-1/DCC complex reveals that Netrin-1 EGF3 domain is crucial for the Netrin-1/DCC interaction, but not the EGF1 and EGF2 domains [34]. Mutagenesis key amino acid residues (His407, Gln442 or Gln443) at EGF3 domain completely disrupted Netrin-1/DCC binding [19]. EGF1/2 domains are required for switching attractive signaling into repulsive signaling when Unc5 coexists with DCC [20]. Thus, we first generated full-length and EGF3-deleted (Netrin-1 Δ407–443) Netrin-1 constructs (
Figure 1A) and transiently expressed these constructs in cultured HEK293T cells for GST pulldown. Binding was observed for the constructs that expressed Netrin-1 but not Netrin-1 (Δ407–443) (
Figure 1C). These results suggest that the domain of EGF3 containing amino acids 407–443 is responsible for the interaction of Netrin-1 with DCC.
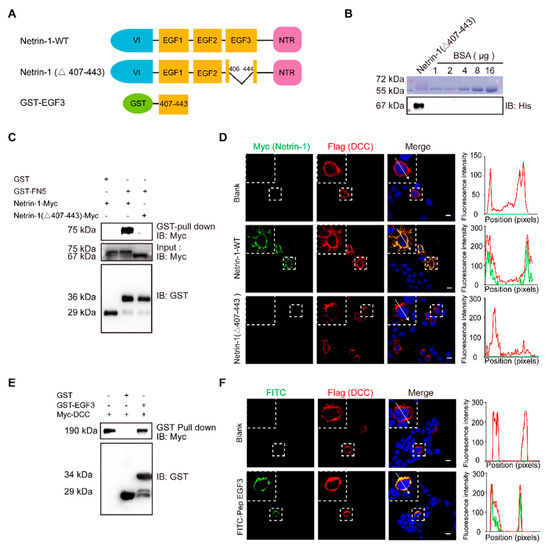
The EGF3 domain of Netrin-1 is the critical region for Netrin-1/DCC interaction. (
) Illustrations of Netrin-1 constructs. The Netrin-1 constructs are Netrin-1 full-length, Netrin-1-(∆407–443) and GST-EGF3. (
) Purified Netrin-1-(∆407–443)-Myc/His were quantified through a comparison with the BSA standard curve. (
) Immunoblotting of the pull-down fraction by the GST-DCC FN5 domain fusion protein and GST alone. (
) Immunofluorescence images of Flag-DCC expressing 293T cells incubated with control medium, Netrin-1-myc/his and Netrin-1-(∆407–443) -Myc/his for 30 min, respectively, and the quantitative analysis. Scale bar, 10 µm. (
) Immunoblotting of the pull-down fraction by the GST-EGF3 fusion protein and GST along. (
) Immunofluorescence images of Flag-DCC expressing 293T cells incubated with control medium and FITC labeled Pep EGF3 for 30 min, respectively, and the quantitative analysis. Scale bar, 10 µm.
To investigate the role of EGF3 domain in Netrin-DCC, we first purified myc-tagged Netrin-1 or truncated Netrin-1 (Δ407–443) (
Figure 1B). Next, HEK293T cells were transfected with a plasmid carrying DCC cDNA and incubated with Netrin-1 protein, and then Netrin-1 binding was assessed by immunostaining. We found that full-length Netrin-1, but not truncated Netrin-1, was able to bind to DCC at the plasma membrane (
Figure 1D). To determine whether the EGF3 domain can interact with DCC, we generated a GST-EGF3 construct to pull down lysed cells overexpressing DCC. We observed specific binding of DCC with GST-EGF3 (
Figure 1E). Furthermore, we synthesized the FITC-labelled peptide EGF3 and confirmed that, like full-length Netrin-1, it was able to bind to DCC at the plasma membrane (
Figure 1F). Taken together, our data indicate that the EGF3 domain is responsible for the interaction of Netrin-1 with DCC.
3. The EGF3 Domain Induces Phosphorylation of Downstream Pathways of Netrin-1
The interaction of Netrin-1 and DCC activates multiple signaling pathways, including FAK, SFK and ERK [16,35]. Focal adhesion kinase (FAK) is one of the major tyrosine phosphorylation activities linked to Netrin-1/DCC signaling. Netrin-1 activates FAK phosphorylation and Src family protein tyrosine kinases (SFKs) phosphorylation [17]. Both FAK and SFK signaling pathways play a pivotal role in the developmental stage of nervous system and injury repair [36,37]. Moreover, Netrin-1 is able to activate the extracellular signal-regulated kinase-1/2 (ERK1/2), which mediates action of Mitogen-activated protein kinase (MAPK) pathway [38,39,40]. To investigate whether the EGF3 domain is critical for the activation of downstream signaling pathways via the binding of Netrin-1 to DCC, we first examined the effect of the EGF3 domain on the tyrosine 861 phosphorylation of FAK (The interaction of Netrin-1 and DCC activates multiple signaling pathways, including FAK, SFK and ERK [16][35]. Focal adhesion kinase (FAK) is one of the major tyrosine phosphorylation activities linked to Netrin-1/DCC signaling. Netrin-1 activates FAK phosphorylation and Src family protein tyrosine kinases (SFKs) phosphorylation [17]. Both FAK and SFK signaling pathways play a pivotal role in the developmental stage of nervous system and injury repair [36][37]. Moreover, Netrin-1 is able to activate the extracellular signal-regulated kinase-1/2 (ERK1/2), which mediates action of Mitogen-activated protein kinase (MAPK) pathway [38][39][40]. To investigate whether the EGF3 domain is critical for the activation of downstream signaling pathways via the binding of Netrin-1 to DCC, we first examined the effect of the EGF3 domain on the tyrosine 861 phosphorylation of FAK (
Figure 2A). We found that deletion of EGF3 inhibited FAK phosphorylation in cultured cortical neurons (
Figure 2B,C), thus confirming an essential role for EGF3 in the Netrin-1 signaling pathway. Next, cultured cortical neurons were stimulated with the GST-EGF3 fusion protein for 20 min, and western blotting further confirmed that treatment with the GST-EGF3 fusion protein induced FAK, ERK and SFK phosphorylation (
Figure 2D–G). Taken together, these results suggest that the EGF3 domain of Netrin-1 functions as a potent stimulator to activate downstream signaling pathways.
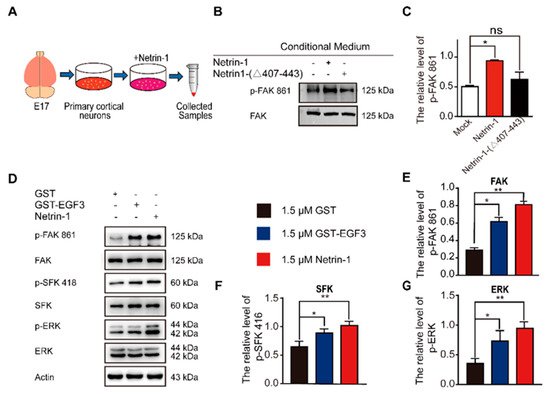
EGF3 domain activates downstream pathways of Netrin-1. (
) Experimental procedure. (
) Cortical neurons (E17 and DIV3) were stimulated with Netrin-1, Netrin-1-(∆407–443) or control medium for 20 min. Cell lysates were incubated with an anti-p-FAK-861 antibody to confirm Netrin-1 activity. (
) Quantitative analysis of the western blot results shown in (
).
= 3. (
) Cortical neurons (DIV3) were treated with 1.5 μM GST (+), 1.5 μM GST-EGF3 fusion protein (+) or 1.5 μM Netrin-1 (+) medium for 20 min. Cell lysates were incubated with anti-p-FAK-861, anti-p-SFK-418 and anti-p-ERK-202/204. (
–
) Quantitative analysis of the western blot results shown in (
).
= 3. Data are presented as the means ± SEM. One-way ANOVA was used for all statistical analyses shown in this figure (*
< 0.05, **
< 0.01; ns, not significant). Details of data analysis are seen in
.
4. Peptide E1 and E2 Interact with DCC to Activate the Downstream Signaling Pathway of Netrin-1
As previously described, a small peptide derived from the EGF3 domain of Netrin-1 (amino acids 423–433) induced ERK phosphorylation [27]. We synthesized peptides E1 (residues 407–422) and E2 (residues 423–433) that were derived from the EGF3 domain of Netrin-1 (
A). We found that incubation of peptide E1 increased the levels of phosphorylated FAK and SFK (
B,D,E) in the culture cells in a dose-dependent manner but did not alter the levels of phosphorylated ERK (
B,F). Unlike peptide E1, peptide E2 not only induced the tyrosine phosphorylation of FAK (
C,G) and the tyrosine phosphorylation of SFK (
C,H) but also induced the tyrosine phosphorylation of ERK (
C,I) in cultured cortical neurons. The control peptide (Pep Ctrl) had no detectable effect on the tyrosine phosphorylation of FAK, SFK and ERK (
). Furthermore, we observed that the phosphorylation of components of the FAK signaling pathway induced by peptide E1 was time-dependent (
). These results indicate that E1 and E2 are the minimal functional peptides of Netrin-1.
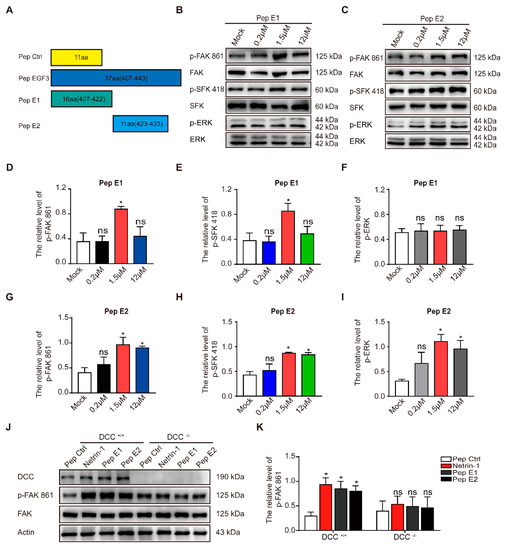
The minimal sequences of the EGF3 domain activates downstream pathways of Netrin-1. (
) Illustrations of the peptides. (
) Cortical neurons (DIV3) were stimulated with gradient concentrations of peptide E1. Cell lysates were incubated with anti-p-FAK-861, anti-p-SFK-418 and anti-p-ERK-202/204. (
) Cultured cortical neurons were treated with gradient concentrations of peptide E2. Lysates were collected and incubated with the indicated antibodies to measure the phosphorylation of FAK, SFK and ERK. (
–
) Quantification of the extent of FAK, SFK and ERK phosphorylation induced by peptide E1 in neurons is shown in (
).
= 3. (
–
) Quantification of the extent of FAK, SFK and ERK phosphorylation induced by peptide E2 is shown in (
).
= 3. (
) Cortical neurons derived from DCC wild-type and homozygote mutant embryos were stimulated with Netrin-1 or Netrin-1-derived peptides and were lysed. The resulting lysates were subjected to immunoblotting. (
) Quantification of FAK PY861 and DCC levels in DCC mutant neurons is shown in (
).
= 3. The data are presented as the means ± SEMs. One-way ANOVA was used for all statistical analyses shown in this figure (*
< 0.05; ns, not significant). Details of data analysis are seen in
.
To determine whether the activation of the FAK signaling pathway is DCC-dependent, we measured the extent of tyrosine phosphorylation of FAK induced by these peptides in DCC
+/+
and DCC
−/−
neurons. We found that both peptides E1 and E2 failed to induce the phosphorylation of FAK in DCC
−/−
neurons (
J,K). We also found that synthesized N-terminally Flag-tagged peptide E1 and E2 (Flag-E1 and Flag-E2) direct bind with DCC-FN5 domain (
).
5. Discussion
Our study identified a core peptide sequence of Netrin-1 that is crucial to functional recovery after ICH. By interacting with DCC, this Netrin-1-derived peptide protected against neuronal death due to secondary damage after ICH and is thus a promising treatment for ICH.
The therapeutic application of Netrin-1, a high-molecular-weight protein, requires identification of the minimal functional sequence of Netrin-1 that promotes downstream signaling. In our study, peptides E1 (residues 407–422) and E2 (residues 423–433), which are derived from the EGF3 domain of Netrin-1, respectively, were generated. We found that peptide E1 (residues 407–422) protected against neuronal death after ICH.
We found that Netrin-1 and DCC expression levels were increased in NLT cells after hemin treatment. Hemin is released from hemoglobin after ICH and induces secondary injury to both glia and neuronal cells [46,47,48]. Additionally, we also observed increased protein levels of Netrin-1 and DCC following experimental ICH. Additionally, we also observed increased protein levels of netrin-1 and DCC following experimental ICH. These results are consistent with previous results showing that netrin-1 plays an important role in recovery after ICH [49].
We found that Netrin-1 and DCC expression levels were increased in NLT cells after hemin treatment. Hemin is released from hemoglobin after ICH and induces secondary injury to both glia and neuronal cells [41][42][43]. Additionally, we also observed increased protein levels of Netrin-1 and DCC following experimental ICH. Additionally, we also observed increased protein levels of netrin-1 and DCC following experimental ICH. These results are consistent with previous results showing that netrin-1 plays an important role in recovery after ICH [44].
Furthermore, we found that peptide E1 specifically activated the FAK and SFK signaling pathways. Therefore, peptide E1 may have highly specific effects and be effective in the recovery process after ICH. Unlike peptide E1, peptide E2 can activate ERK signaling, which promotes neuronal death. This may be caused by a Cx(1–2)Cx(3–4)Tx(0 –1)G motif in E2 which is able to activate ERK signaling pathway [28]. This indicates that different Netrin-1-derived peptides perform different functions.
The main receptors of Netrin-1 are DCC and members of the UNC5 family [50]. The role of DCC and UNC5H2 in ICH remains controversial [51,52]. The sequence of Netrin-1 we identified is critical to the interaction of Netrin-1 with DCC [19]. These results suggest that DCC may participate in Netrin-1-induced functional recovery after ICH. Further studies are needed to investigate the underlying mechanisms of these processes and the related signaling pathways. In future experiments, we will assess the biological properties of peptide E1 to explore its toxicity and half-life for clinical application.
The main receptors of Netrin-1 are DCC and members of the UNC5 family [45]. The role of DCC and UNC5H2 in ICH remains controversial [46][47]. The sequence of Netrin-1 we identified is critical to the interaction of Netrin-1 with DCC [19]. These results suggest that DCC may participate in Netrin-1-induced functional recovery after ICH. Further studies are needed to investigate the underlying mechanisms of these processes and the related signaling pathways. In future experiments, we will assess the biological properties of peptide E1 to explore its toxicity and half-life for clinical application.
References
- Kirshner, H.S. Medical management of intracerebral haemorrhage: Back to the basics. Int. J. Clin. Pract. 2008, 62, 521–522.
- Anderson, C.S.; Heeley, E.; Huang, Y.; Wang, J.; Stapf, C.; Delcourt, C.; Lindley, R.; Robinson, T.; Lavados, P.; Neal, B.; et al. Rapid blood-pressure lowering in patients with acute intracerebral hemorrhage. N. Engl. J. Med. 2013, 368, 2355–2365.
- Qureshi, A.I.; Mendelow, A.D.; Hanley, D.F. Intracerebral haemorrhage. Lancet (Lond. Engl.) 2009, 373, 1632–1644.
- Keep, R.F.; Hua, Y.; Xi, G. Intracerebral haemorrhage: Mechanisms of injury and therapeutic targets. Lancet Neurol. 2012, 11, 720–731.
- Schrag, M.; Kirshner, H. Management of Intracerebral Hemorrhage: JACC Focus Seminar. J. Am. Coll. Cardiol. 2020, 75, 1819–1831.
- Kandasamy, R.; Idris, Z.; Abdullah, J.M. Surgery of Intracerebral Hemorrhage. In Neurovascular Surgery Surgical Approaches for Neurovascular Diseases; July, J., Wahjoepramono, E.J., Eds.; Springer: Singapore, 2019; pp. 201–210.
- Broughton, B.R.; Reutens, D.C.; Sobey, C.G. Apoptotic mechanisms after cerebral ischemia. Stroke 2009, 40, e331–e339.
- Kristiansen, M.; Graversen, J.H.; Jacobsen, C.; Sonne, O.; Hoffman, H.J.; Law, S.K.; Moestrup, S.K. Identification of the haemoglobin scavenger receptor. Nature 2001, 409, 198–201.
- Madangarli, N.; Bonsack, F.; Dasari, R.; Sukumari-Ramesh, S. Intracerebral Hemorrhage: Blood Components and Neurotoxicity. Brain Sci. 2019, 9, 316.
- Deiner, M.S.; Kennedy, T.E.; Fazeli, A.; Serafini, T.; Tessier-Lavigne, M.; Sretavan, D.W. Netrin-1 and DCC mediate axon guidance locally at the optic disc: Loss of function leads to optic nerve hypoplasia. Neuron 1997, 19, 575–589.
- Kennedy, T.E.; Serafini, T.; de la Torre, J.R.; Tessier-Lavigne, M. Netrins are diffusible chemotropic factors for commissural axons in the embryonic spinal cord. Cell 1994, 78, 425–435.
- Flores, C. Role of netrin-1 in the organization and function of the mesocorticolimbic dopamine system. J. Psychiatry Neurosci. Jpn. 2011, 36, 296–310.
- Ren, X.R.; Ming, G.L.; Xie, Y.; Hong, Y.; Sun, D.M.; Zhao, Z.Q.; Feng, Z.; Wang, Q.; Shim, S.; Chen, Z.F.; et al. Focal adhesion kinase in netrin-1 signaling. Nat. Neurosci. 2004, 7, 1204–1212.
- Zhang, J.-H.; Zhao, Y.-F.; He, X.-X.; Zhao, Y.; He, Z.-X.; Zhang, L.; Huang, Y.; Wang, Y.-B.; Hu, L.; Liu, L. DCC-mediated Dab1 phosphorylation participates in the multipolar-to-bipolar transition of migrating neurons. Cell Rep. 2018, 22, 3598–3611.
- Ren, X.R.; Hong, Y.; Feng, Z.; Yang, H.M.; Mei, L.; Xiong, W.C. Tyrosine phosphorylation of netrin receptors in netrin-1 signaling. Neurosignals 2008, 16, 235–245.
- Cirulli, V.; Yebra, M. Netrins: Beyond the brain. Nat. Rev. Mol. Cell Biol. 2007, 8, 296–306.
- Li, W.; Lee, J.; Vikis, H.G.; Lee, S.H.; Liu, G.; Aurandt, J.; Shen, T.L.; Fearon, E.R.; Guan, J.L.; Han, M.; et al. Activation of FAK and Src are receptor-proximal events required for netrin signaling. Nat. Neurosci. 2004, 7, 1213–1221.
- Liu, G.; Beggs, H.; Jürgensen, C.; Park, H.-T.; Tang, H.; Gorski, J.; Jones, K.R.; Reichardt, L.F.; Wu, J.; Rao, Y. Netrin requires focal adhesion kinase and Src family kinases for axon outgrowth and attraction. Nat. Neurosci. 2004, 7, 1222–1232.
- Finci, L.I.; Krüger, N.; Sun, X.; Zhang, J.; Chegkazi, M.; Wu, Y.; Schenk, G.; Mertens, H.D.T.; Svergun, D.I.; Zhang, Y.; et al. The crystal structure of netrin-1 in complex with DCC reveals the bifunctionality of netrin-1 as a guidance cue. Neuron 2014, 83, 839–849.
- Xu, K.; Wu, Z.; Renier, N.; Antipenko, A.; Tzvetkova-Robev, D.; Xu, Y.; Minchenko, M.; Nardi-Dei, V.; Rajashankar, K.R.; Himanen, J.; et al. Neural migration. Structures of netrin-1 bound to two receptors provide insight into its axon guidance mechanism. Science 2014, 344, 1275–1279.
- Lu, H.; Wang, Y.; He, X.; Yuan, F.; Lin, X.; Xie, B.; Tang, G.; Huang, J.; Tang, Y.; Jin, K.; et al. Netrin-1 hyperexpression in mouse brain promotes angiogenesis and long-term neurological recovery after transient focal ischemia. Stroke 2012, 43, 838–843.
- Guo, D.; Zhu, Z.; Zhong, C.; Peng, H.; Wang, A.; Xu, T.; Peng, Y.; Xu, T.; Chen, C.S.; Li, Q.; et al. Increased Serum Netrin-1 Is Associated with Improved Prognosis of Ischemic Stroke. Stroke 2019, 50, 845–852.
- Wu, T.W.; Li, W.W.; Li, H. Netrin-1 attenuates ischemic stroke-induced apoptosis. Neuroscience 2008, 156, 475–482.
- Navankasattusas, S.; Whitehead, K.J.; Suli, A.; Sorensen, L.K.; Lim, A.H.; Zhao, J.; Park, K.W.; Wythe, J.D.; Thomas, K.R.; Chien, C.B.; et al. The netrin receptor UNC5B promotes angiogenesis in specific vascular beds. Development (Camb. Engl.) 2008, 135, 659–667.
- Bai, L.; Mei, X.; Wang, Y.; Yuan, Y.; Bi, Y.; Li, G.; Wang, H.; Yan, P.; Lv, G. The Role of Netrin-1 in Improving Functional Recovery through Autophagy Stimulation Following Spinal Cord Injury in Rats. Front. Cell. Neurosci. 2017, 11.
- Xie, Z.; Huang, L.; Enkhjargal, B.; Reis, C.; Wan, W.; Tang, J.; Cheng, Y.; Zhang, J.H. Intranasal administration of recombinant Netrin-1 attenuates neuronal apoptosis by activating DCC/APPL-1/AKT signaling pathway after subarachnoid hemorrhage in rats. Neuropharmacology 2017, 119, 123–133.
- Cui, M.Z. Potential therapeutics for myocardial ischemia-reperfusion injury. Focus on “Induction of cardioprotection by small netrin-1-derived peptides”. Am. J. Physiol. Cell Physiol. 2015, 309, C97–C99.
- Li, Q.; Cai, H. Induction of cardioprotection by small netrin-1-derived peptides. Am. J. Physiol. Cell Physiol. 2015, 309, C100–C106.
- Mazelin, L.; Bernet, A.; Bonod-Bidaud, C.; Pays, L.; Arnaud, S.; Gespach, C.; Bredesen, D.E.; Scoazec, J.Y.; Mehlen, P. Netrin-1 controls colorectal tumorigenesis by regulating apoptosis. Nature 2004, 431, 80–84.
- Mehlen, P.; Mazelin, L. The dependence receptors DCC and UNC5H as a link between neuronal guidance and survival. Biol. Cell 2003, 95, 425–436.
- Arakawa, H. Netrin-1 and its receptors in tumorigenesis. Nat. Rev. Cancer 2004, 4, 978–987.
- Chen, J.-Y.; He, X.-X.; Ma, C.; Wu, X.-M.; Wan, X.-L.; Xing, Z.-K.; Pei, Q.-Q.; Dong, X.-P.; Liu, D.-X.; Xiong, W.-C.; et al. Netrin-1 promotes glioma growth by activating NF-κB via UNC5A. Sci. Rep. 2017, 7, 5454.
- Yurchenco, P.D.; Wadsworth, W.G. Assembly and tissue functions of early embryonic laminins and netrins. Curr. Opin. Cell Biol. 2004, 16, 572–579.
- Dun, X.P.; Parkinson, D.B. Role of Netrin-1 Signaling in Nerve Regeneration. Int. J. Mol. Sci. 2017, 18, 419.
- Lai Wing Sun, K.; Correia, J.P.; Kennedy, T.E. Netrins: Versatile extracellular cues with diverse functions. Development (Camb. Engl.) 2011, 138, 2153–2169.
- Tang, T.; Gao, D.; Yang, X.; Hua, X.; Li, S.; Sun, H. Exogenous Netrin-1 Inhibits Autophagy of Ischemic Brain Tissues and Hypoxic Neurons via PI3K/mTOR Pathway in Ischemic Stroke. J. Stroke Cerebrovasc. Dis. Off. J. Natl. Stroke Assoc. 2019, 28, 1338–1345.
- Boyer, N.P.; Gupton, S.L. Revisiting Netrin-1: One Who Guides (Axons). Front. Cell. Neurosci. 2018, 12, 221.
- Forcet, C.; Stein, E.; Pays, L.; Corset, V.; Llambi, F.; Tessier-Lavigne, M.; Mehlen, P. Netrin-1-mediated axon outgrowth requires deleted in colorectal cancer-dependent MAPK activation. Nature 2002, 417, 443–447.
- Guo, Y.J.; Pan, W.W.; Liu, S.B.; Shen, Z.F.; Xu, Y.; Hu, L.L. ERK/MAPK signalling pathway and tumorigenesis. Exp. Ther. Med. 2020, 19, 1997–2007.
- Kefeli, U.; Ucuncu Kefeli, A.; Cabuk, D.; Isik, U.; Sonkaya, A.; Acikgoz, O.; Ozden, E.; Uygun, K. Netrin-1 in cancer: Potential biomarker and therapeutic target? Tumour Biol. J. Int. Soc. Oncodev. Biol. Med. 2017, 39, 1010428317698388.
- Wang, J.; Zhuang, H.; Doré, S. Heme oxygenase 2 is neuroprotective against intracerebral hemorrhage. Neurobiol. Dis. 2006, 22, 473–476.
- Cai, Y.; Cho, G.S.; Ju, C.; Wang, S.L.; Ryu, J.H.; Shin, C.Y.; Kim, H.S.; Nam, K.W.; Jalin, A.M.; Sun, W.; et al. Activated microglia are less vulnerable to hemin toxicity due to nitric oxide-dependent inhibition of JNK and p38 MAPK activation. J. Immunol. 2011, 187, 1314–1321.
- Min, H.; Choi, B.; Jang, Y.H.; Cho, I.H.; Lee, S.J. Heme molecule functions as an endogenous agonist of astrocyte TLR2 to contribute to secondary brain damage after intracerebral hemorrhage. Mol. Brain 2017, 10, 27.
- Wang, J.; Zhai, W.; Yu, Z.; Sun, L.; Li, H.; Shen, H.; Li, X.; Liu, C.; Chen, G. Neuroprotection Exerted by Netrin-1 and Kinesin Motor KIF1A in Secondary Brain Injury following Experimental Intracerebral Hemorrhage in Rats. Front. Cell. Neurosci. 2017, 11, 432.
- Freitas, C.; Larrivee, B.; Eichmann, A. Netrins and UNC5 receptors in angiogenesis. Angiogenesis 2008, 11, 23–29.
- Castets, M.; Coissieux, M.M.; Delloye-Bourgeois, C.; Bernard, L.; Delcros, J.G.; Bernet, A.; Laudet, V.; Mehlen, P. Inhibition of endothelial cell apoptosis by netrin-1 during angiogenesis. Dev. Cell 2009, 16, 614–620.
- Lu, X.; Le Noble, F.; Yuan, L.; Jiang, Q.; De Lafarge, B.; Sugiyama, D.; Bréant, C.; Claes, F.; De Smet, F.; Thomas, J.L.; et al. The netrin receptor UNC5B mediates guidance events controlling morphogenesis of the vascular system. Nature 2004, 432, 179–186.