Withaferin A (WFA) was identified as the most active phytocompound of the plant Withania somnifera (WS) and as having multiple therapeutic/ameliorating properties (anticancer, antiangiogenic, anti-invasive, anti-inflammatory, proapoptotic, etc.) in case of various diseases. This paper focuses on the effects and results that WFA has on dermatological conditions.
- Withania somnifera
- withaferin A
- dermatology
- dermatological disorders
- melanoma
1. Introduction
Dermatological diseases are a wide and diverse class of diseases that range from simple rashes to severe skin infections, which may have various causes as follows: allergens, heat, disorders of some systems, drug therapies, etc. They affect about 30% of the population, with the most serious consequences on the quality of life or could even lead to death, causing physical discomfort, embarrassment, the patient sometimes suffering socio-professional restrictions. Moreover, chronic conditions lead to the need for repeated sick leave, with financial repercussions on the patients [1].
Withania somnifera (L.) Dunal (WS) is a largely used medicinal plant (known also under the name Ayurvedic Ashwagandha) with easily recognizable anticarcinogenic importance and role, leaves of the shrub being used as raw material for the extraction of withanolide steroid compounds [2][3][2,3]. This Ayurvedic herb, known across the world for its many beneficial health care practices and roles since ancient times [4], is a member of the Solanaceae family. It offers therapeutical effects for many human diseases, including arthritis, epilepsy, depression, diabetes, and has palliative effects (such as analgesics, growth promoting, regenerating, rejuvenating, etc.). In patients suffering from the above-mentioned disorders, some clinical studies focused on the various sections/parts of the herb that have shown protection and have proved to be safe [5].
Withaferin A (WFA) was the first isolated, most common, and frequently identified withanolide in WS [6], with a detailed chemical structure depicted in Figure 1.
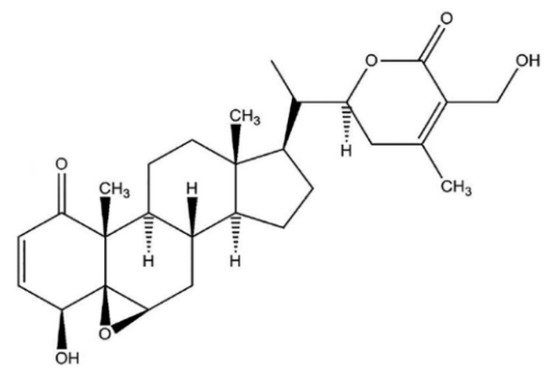
Figure 1. WFA chemical structure.
Worldwide, several studies were conducted on the WFA properties and on its beneficial action in anticancerous therapy. Nowadays, there are numerous published data on the antitumoral action of WFA, performed by making use of different techniques (xenografts, orthotopic tumor models, and cell cultures of multiple human cancers) [3][7][3,7]. Many of these studies revealed the cytotoxic, antimigratory, proapoptotic, and anti-invasive actions of WFA, in the case of different types of cancers. Various molecular targets in cancerous cells are modulated by WFA, especially signaling molecules, enzymes, and various proteins that are essential for the survival of tumor cells [8][9][8,9]. WFA may also operate through processes such as triggering proteinase-activated receptor 4 (PAR-4) and suppressing heat shock protein 90 (HSP90) in prostate cancer cells [10][11][10,11], obstructing nuclear factor kappa-light-chain enhancer of activated B cells (NF-κB) [12], triggering deoxyribonucleic (DNA) acid damage checkpoint (G2-M) inhibition; forkhead box O3 (FoxO3a) and apoptotic protein Bim adjustment in breast cancer [13].
Through the intrinsic or extrinsic routs, WFA was indicated to trigger apoptosis in the human breast, prostate, neck and head, leukemia, and melanoma tumor cells by stimulating several proteases and caspases and decreasing mitochondrial membrane voltage, which induces damage of several substrates such as poly adenosine diphosphate (ADP)-ribose) polymerase cleavage and cytoskeletal proteins [14][15][16][14,15,16].
Moreover, WFA targets some proteins resulted from the anti-stress pathway, implying the improvement and enhancement in reactive oxygen species (ROS) expression. The increased ROS amount also enables antioxidant pathways and causes disequilibrium in ROS/cytoprotection action, determining the cancer cells stage. WFA therapy activates four stress response proteins for oxidative damage reduction and restores homeostasis during and after the treatment. The WFA therapy upregulates proteins such as aldose reductase, heme oxygenase, iron–sulfur, and sepiapterin reductase as a response to oxidative stress, while glutathione peroxidase 1, hydroperoxide, and phospholipids are downregulated [17][18][17,18].
2. Withaferin in Skin Cancers
Skin cancer is known to be caused by uncontrolled growth of abnormal epidermal cells (epidermis is the outer layer of the skin), as a result of irreparable DNA damage, a fact that inevitably leads to mutations. Skin cancers are classified into the following main types—basal cell carcinoma (BCC), squamous cell carcinoma (SCC), melanoma, and Merkel cell carcinoma, all of them among the most common cancers worldwide [19][20][21,22].
The most lethal cutaneous neoplasm is considered to be malignant melanoma. Achieved drug resistance commonly appears sometime after concrete tumor response, explaining the necessity for new remedies [21][22][23,24].
WFA was found to inhibit in vivo mouse melanoma (B16F1) tumor development [23][25]. For a different heterograft skin cancer type operating with 92.1 uveal melanoma cells, following the treatment with WFA, approximately 29% of mice presented total clinical reaction, whereas 43% of the subjects indicated cancer evolution when the treatment was interrupted [20][22]. As other recent published data revealed, WFA decreased the tumor abundance; however, it did not reduce the prevalence of 12-O-tetradecanoylphorbol-13-acetate (TPA) favored and dimethyl-benzanthracene DMBA-induced mice skin cancer development [24][26], in part by triggering activator protein-1 (AP-1) and inhibiting the expression of acetyl coenzyme A carboxylase-1 (ACC1) [25][26][27,28]. Chemopreventive and antitumor effects of WFA, as well as the mechanisms involved and its use as supportive therapy in skin cancers, are analyzed in the following subsections.
2.1. Chemopreventive Effects
Some studies indicated the chemopreventive capacity of WS that can be useful against skin carcinogenesis [27][28][29,30].
WFA has a thoroughly analyzed antitumoral function in experimental animal models of human cancer; however, it has uncertain chemopreventive capacity. Published data have shown that in the well-established tumor promotion model for JB6 P+ skin epidermal cells WFA suppressed the cell transformation and proliferation induced by the tumor promoter TPA [25][27]. It is interesting that TPA deactivated isocitrate dehydrogenase 1 (IDH1), reversed by WFA. In mouse skin tissue, similar findings were presented. Thus, metabolism was considered as the possible mechanism of action. It was found that mitochondrial activities such as membrane voltage, complex I action, and mitochondrial breathing have been decreased by TPA therapy. Nevertheless, WFA restrained all these downregulations. Moreover, α-ketoglutarate levels, an IDH1 product, were investigated and WFA inhibited its decrease in TPA therapy. Furthermore, the lactate level was observed as an indicator for glycolysis, WFA inhibiting its increase due to treatment with tumor promoters [24][26].
In 2019, Xu et al. [29][31] studied the way WFA inhibited the IDH1 favored skin cancer. Data obtained revealed the WFA ability to stabilize IDH1 by suppressing the ubiquitin-proteasome pathway (UPP). As a result of clarifying the process of IDH1 suppressing carcinogenesis, the outcomes reflected that the activity on LDH was inhibited by the increase of IDH1, while the action of mitochondrial complex I was enhanced. Moreover, increased IDH1 determined prolyl hydroxylase (PHD) activation through its product α-ketoglutarate (KG) and suppressed HIF-1α downward signaling route. Findings reveal that WFA partly suppresses cancer evolution through stabilizing IDH1, conducting to the deactivation of HIF-1α signaling [29][31].
Li et al. [30][32] indicated that the decrease in tumor multiplicity of DMBA-initiated and TPA favored the development of mouse skin tumors, partially by inhibiting ACC1 expression and triggering the activator protein-1 (AP-1). The chemopreventive ability of WFA was examined in a skin carcinogenesis mouse model, chemically triggered. Pathological studies found that the development of skin tumors was suppressed substantially by WFA. Morphological studies on skin tissue indicate that during skin cancer, WFA inhibited cell growth instead of causing apoptosis [29][31]. Microarray examination of the antibody showed that WFA suppressed the ACC1 upregulation caused by a carcinogenic agent, which was further verified in a skin cell transformation model. The knockdown of ACC1 inhibited unbundling-independent development and the activation of oncogenic transformation of skin cells, supported by ACC1 overexpression. Further experiments have shown that WFA blocked ACC1 gene transcription caused by tumor promoters, stopping activator protein 1 activation. The expression levels of ACC1 in melanoma cells were also inhibited by WFA. Eventually, research on human skin cancer tissue showed that the ACC1 in tumors was upregulated, compared with neighboring healthy tissues. Results indicate also that WFA may have a chemoprevention capacity, and that ACC1 may represent a key objective for WFA [24][26].
Summarizing, it can be concluded that chemical-induced skin cancer is inhibited by WFA, and ACC1 can be an effective target for WFA. These data can be extended to melanoma as the expression of ACC1 in melanoma cells is also inhibited by WFA. The study focused on how WFA controls ACC1 and the way ACC1 favors cancer evolution [30][32].
2.2. Antitumor Effects
Cytotoxicity of WS root mineral extract on A375 (ATCC) human malignant melanoma cell line was tested for the first time in the study of Halder et al. [24][26]. The extract obtained from blunt WS was tested for A375 cell cytotoxicity with the MTT procedure (using yellow tetrazolium MTT reagent, known also as 3-(4,5-dimethylthiazolyl-2)-2, 5-diphenyltetrazolium bromide) [31][33], which is based on the reaction presented in Figure 2Figure 3.
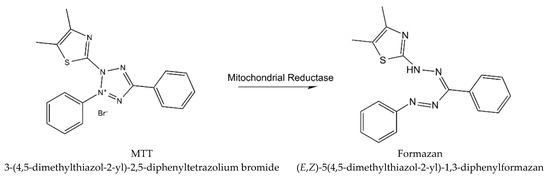
Figure 23.
The reaction describing MTT procedure (metabolism of MTT to a formazan salt).
The morphology of the treated ATCC has been visualized by phase-contrast and fluorescence microscopy. The deoxyribonucleic acid (DNA) fragmentation of crude extract cells treated with agarose gel electrophoreses was tested. WS root crude extract has the ability to minimize the viable dosage and time-dependent cell count. In contrast to untreated or vehicular regulation, morphological changes in the ATCC were also observed for treated types. Under a fluorescence microscope, apoptotic body and nuclear blebbing were detected in the 4′,6-diamidino-2-phenylindole (DAPI) stain-treated cells, allowing a ladder of sequenced DNA observance in the treated cells. Moreover, the fresh aqueous extract of WS has a significant cytotoxic effect related to the human malignant ATCC [32][34].
WFA has a low antitumor and radio-sensitizing function [33][35]. Published data seek to evaluate the tumor sensitizing effect of WFA on B16F1 melanoma reaction with or without local hyperthermal fractional and acute radiotherapy. The following two series of parameters (each or in association) were described as being used in the adult C57BL mice, injected intradermally with 5 × 105 tumor cells (B16F1 melanoma) on the dorsal skin, with 100 ± 10 mm3 tumors: 1. fractionated radiotherapy (RT) (2 Gy × 5 days a week, 4 weeks); WFA (15 mg/kg, 5 days a week, 3 weeks); and local hyperthermia (HT) (43 °C once a week, for 3 weeks) or 2. short-term high-level RT (40 Gy); WFA (40 mg/kg); and HT (43 °C, 30 min). The treatment reaction was evaluated by testing the regression of the tumor, development delay, and animal longevity. In acute RT + HT, 50% of the response was partial, then increased to 62.5% with the WFA mixture. In fractional systems, the mixture of trimodality led to 100% PR. The growth delay (GD) increased with acute RT + HT and WFA + RT in contrast with RT alone that subsequently grew in trimodality therapy. WFA + RT + HT fragmented produced greater GD and survival rates, in a 3-week treatment, higher than other therapies. Finally, in a fractional regimen, HT is less efficient than WFA, which is a better radiosensitizer and facilitates a reduction in radiation exposure for radioresistant tumors such as melanoma by associating the nontoxic doses of WFA with fractional RT, with and without the addition of HT [34][36].
The apoptotic mechanism initiated by WFA in the melanoma cell lines included mitochondrial translocation, cytochrome c releasing, transmembrane modifications, and activation of caspases 9 and 3. Early reactive oxygen species (ROS) production may also be needed for WFA cytotoxicity [14]. Skin melanoma, which is malignant, highly resistant, and heterogeneous to conventional cancer chemotherapy, constitutes the most aggressive and deadly form of skin cancer. Against several cancer cells, WFA had antitumor action. WFA was first tested in an association of four different melanoma human cells and cellular dynamics research. With IC50 varying from 1.8 to 6.1 μM, WFA causes apoptotic cell death. WFA associated Bcl-2/Bax/Bcl-2/Bim apoptosis with low cell ratios. Both lines include the WFA-led mitochondrial-pathway apoptotic process, which was linked to Bcl 2, BaX mitochondrial translocation, cytochrome c release, transmembrane dissipation potential (mm), caspase 9, and DNA fragmentation. In this way, the process is linked with Blu-2 downregulation. WFA cytotoxicity includes the synthesis and depletions of early ROS and ROS inhibitions, leading to the full abolition of mitochondrial and nuclear events by the antioxidant N-acetylcysteine. These effects contribute to WFA therapeutic effect against human melanoma [14].
Histopathological tests on Wistar rats (adult male weighing 125–150 g) cutaneous sample tissue have found enhanced malignancy, with ultraviolet B (UVB) radiation exposure (wavelength of 294 nm) for 20 days, succeeded by benzoyl peroxide topical treatments that promoted tumors (dose of 20 mg/animal/0.2 mL acetone). For animals pretreated with 1-oxo-5- and 6beta-epoxy-with-2-enolide (obtained from WS roots) was avoided the occurrence of skin carcinomas when animals were exposed to UVB radiation or to benzoyl peroxide associated with UVB. Administering 1-oxo-5beta or 6beta-epoxy-with-2-enolide also stops malignancy in the skin tissue, following exposure to UVB radiation and benzoyl peroxide. In rats exposed to UVB radiation, immunohistochemical stains of cutaneous tissues reveal p53 + foci (cells that carry the mutant p53 protein), whereas, in animals pretreated with 1-oxo-5beta/6-beta-epoxy-with-2-enolide, the absence of the p53 + foci has been noted. These findings show that the effects on the skin carcinomas caused by UVB radiation are possible in 1-oxo-5beta/6-beta-epoxy-witha-2-enolide [35][37].
2.3. Support Therapy
While previous research studied the impact of WFA on the viability and proliferation of melanoma cells, a thorough analysis on WFA and its therapeutic and concentration interval in which WFA can be used for avoiding side effects was performed using a large melanoma cell line variety and regular fibroblast [26][36][28,38]. The B16F1 melanoma and fibrosarcoma, cultivated in C57BL and in Swiss albino mice were tested for the function of WFA, either alone or in combination with fractionated and acute radiotherapy, and/or hyperthermia. Acute dose gamma radiation of 30 or 50 Gy or five fractions of 10 Gy was applied locally on tumors. WFA was injected intraperitoneally, in a dose of 40 mg/kg, 1 h prior to acute radiation, or 30 mg/kg previous to 10 Gy fragment. Local hyperthermia, for 30 min at 43 °C, succeeded the acute RT or the first 10 Gy fraction. WFA, hyperthermia, and radiation administered separately or in bimodality therapy in melanoma did not generate a complete response (CR). Several CR were identified in fibrosarcoma to enhance following bimodality therapies. The synergistic rise after trimodality therapy of CR was up to 37% in melanoma and up to 64% in fibrosarcoma. Fractioned radiotherapy (10 Gy x 5) was more efficient (25% CR) on melanoma than acute 50 Gy (0% CR), whereas fibrosarcoma response showed no variation between the two procedures. WFA along with fractionated radiotherapy determined a synergistic rise in CR for both tumors; this effect is further amplified by hyperthermia. It is important to analyze WFA’s usefulness in increasing the therapeutic reaction of radiation-resistant tumors against fractionated radiotherapy [37][39]. Some studies have reported the inhibition of in vivo development of multiple tumor xenografts including uveal melanoma and the sensitizing effect of administering WFA on B16F1 melanoma cells to radiotherapy [34][38][36,40]. The WFA therapy was found to induce TRIM16 mRNA expression in melanoma cell lines, while TRIM16 was expected to trigger the highest cytotoxic action. MelCV melanoma cells, compared to MelJD cells, were shown to be less susceptible to WFA therapy. The TRIM16 basal expression of the MelCV cells was lower than that of the MelJD cells [39][41]. It is believed that, as a result of preexisting lower basal TRIM16, MelCV cells will intrinsically be less susceptible to WFA therapy, and the apoptotic activity of TRIM16 in these cells can be inhibited in different ways. It is not clear how TRIM16 expression is lost in melanoma cells.
Several factors were identified as triggers for neuroblastoma, such as promoter methylation and decreased protein stability; comparable imbalances may appear in melanoma [23][25]. The induction of TRIM16 mRNA expression at increasing WFA concentrations was shown to be modest, indicating that other regulatory mechanisms, such as post-translational changes, that enhance the stability of TRIM16 or prevent its proteasomal degradation, may also be determined by WFA. Studies showing that WFA may inhibit proteasomal degradation, in the site of TRIM16, sustained the above hypothesis [21][23]. Studies on migration consistent with research on breast cancer cell lines have shown that WFA prevented melanoma cell migration. Therefore, it is implied that WFA therapy, together with TRIM16 expression upregulation, can be a possible way of preventing disease development and serving as a support therapy for patients with stage II melanoma [40][42].