The complement system is an essential component of innate immunity. Acting as a first line of defence, the complement system recognises and aids in the elimination of pathogens. Complement proteins also play key roles in homeostasis and they support the inductive of the adaptive immune response.
- complement
- cancer
- cancer treatment
- therapeutic response
1. Introduction
A dynamic relationship exists between the immune system and cancer, owing to the fact that a system designed to defend the host and maintain homeostasis has the potential to promote and foster malignant transformation [1,2][1][2]. Complement, an innate inflammatory system, is no exception to this paradox [3]. Traditionally, the elimination of foreign antigens was considered the primary, if not sole function of complement, however we now understand that complement activities extend beyond this [4]. The complement system, for instance, plays an important role coordinating adaptive immune responses, as an opsonin, in synapse elimination and during angiogenesis [5,6,7,8][5][6][7][8]. Several studies have demonstrated that complement is also capable of recognising and eliminating malignant cells [9]. The net effect of these diverse functions renders the complement system a key player in immune surveillance and homeostasis [4]. The delicate equilibrium between developing tumours and the immune system is well documented, with evasion of immune destruction defined as a hallmark of cancer [10]. In line with reports of an altered immune milieu in several human cancers, dysregulation of the complement system in the cancer setting has been observed [11,12,13,14,15][11][12][13][14][15]. More recently, pro-oncogenic roles for complement cascade components have been described [16,17][16][17]. Analysis of the current literature suggests that the complement system has a dual role in malignancy [4,18][4][18] and whether complement protects against or enables tumour pathogenesis may depend on the context of the tumour microenvironment (TME) [19].
2. The Complement System
In 1901, Jules Bordet described complement as a heat-labile factor that augmented antibody-mediated bacterial lysis [20]. Subsequent discoveries have since established that complement is not a single entity but represents a family of many proteins [21]. The complement system is composed of approximately 50 soluble and membrane-bound complement effectors, regulators and receptors, with the main complement proteins numbered C1-9 [4]. Many complement precursors exist as zymogens, which require cleavage in order to gain functionality [22]. The C3 and C5 convertase enzymes are central to the complement cascade, cleaving C3 and C5 respectively to generate anaphylatoxins (C3a, C5a) and opsonins (C3b, C5b) [4,23][4][23]. The small anaphylatoxin molecules are potent inflammatory mediators with many effector functions [22,24][22][24]. Complement proteins are primarily produced by the liver before systemic dissemination via the bloodstream, however, we now understand that T cells, macrophages, endothelial cells and more recently, cancer cells, are capable of complement production.
2.1. Complement Activation Pathways
There are three pathways by which the complement system may be activated, the classical, the lectin and the alternative pathways (Figure 1). The classical pathway is principally initiated when C1q of the C1 complex (C1q, C1r and C1s) recognises antigen-antibody (Immunoglobulin (Ig) G or IgM containing) immune complexes, but several antibody-independent signals such as C-reactive protein and viral proteins can activate this pathway also [25,26,27,28,29,30,31][25][26][27][28][29][30][31]. Viral and bacterial carbohydrate-based pathogen-associated molecular patterns (PAMPs) activate the lectin pathway by binding to mannose-binding lectin (MBL), ficolins or collectins [32,33,34,35][32][33][34][35]. In the alternative pathway, C3 is spontaneously hydrolysed to C3H2O in a process known as ‘tick-over’ [36,37][36][37]. Bacterial and yeast polysaccharides and damaged tissue are among the initiators of this pathway [38,39][38][39]. The binding of properdin to target microbial surfaces can facilitate the assembly of the alternative pathway C3 convertase [40,41][40][41]. The alternative pathway also acts as an amplification loop for the classical and lectin pathways [23,42][23][42]. Irrespective of the course of activation, the three activation pathways converge to initiate the terminal pathway. This pathway culminates with the assembly of complement components C5b–C9 to form a membrane attack complex (MAC) [43,44][43][44]. MAC insertion into target cell membranes can trigger lysis known as complement-dependent cytotoxicity (CDC) [45], or at sublytic doses may activate signalling pathways to promote cell survival [46,47][46][47].
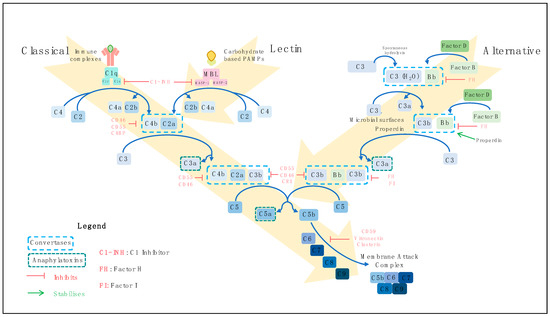
Figure 1. Complement activation pathways. There are three routes by which the complement system can become activated: the classical, the lectin and the alternative pathways. Classical pathway activation is initiated primarily by antigen-antibody immune complexes. C1q of the C1 complex (C1q, C1r and C1s) interacts with the fragment crystallistion (Fc) portion of antigen-bound immunoglobulins, activating C1r, which subsequently cleaves and activates C1s. Activated C1s cleaves C4 into C4a and C4b, and C2 into C2a and C2b leading to assembly of C4bC2a, the C3 convertase. Carbohydrate-based pathogen-associated molecular patterns (PAMPs) trigger activation of the lectin pathway. Mannose-binding lectin (MBL), ficolins or collectins recognise PAMPs, activating MBL-associated serine proteases (MASPs). Similar to the classical pathway, C4 and C2 are cleaved to generate C4bC2a. The classical and lectin complement activation pathways converge at this point to cleave C3 into the potent anaphylatoxin C3a, and C3b, which joins the C3 convertase to form C4bC2aC3b, the C5 convertase. Cleavage of C5 yields the C5a anaphylatoxin and C5b, which polymerises with C6, C7, C8 and C9 to form the membrane-attack complex (MAC). This inserts into target cell membranes to induce lysis. Spontaneous hydrolysis of C3 into C3H2O occurs in the alternative pathway. Cleavage of factor B (FB) by factor D yields Bb, which associates with C3H2O to form a C3 convertase. Cleavage of C3 and FB produces C3b and Bb, respectively. The binding of properdin to microbial surfaces recruits C3b, facilitating the assembly of the C3 convertase (C3bBb), and initiating pathway activation. Subsequent cleavage of C3 produces C3b, which combines with the C3 convertase to form a C5 convertase (C3bBbC3b). From this point, the terminal pathway is initiated to assemble the MAC, similarly to the classical and lectin pathways. Complement activation is regulated at various stages of the pathways by several membrane-bound complement regulatory proteins (Complement receptor 1 (CR1), CD46, CD55 and CD59) and circulating factors (C1-inhibitor (C1-INH), factor H (FH), factor I (FI), C4-binding protein (C4BP), clusterin and vitronectin), which are depicted in red, and properdin, which stabilises the alternative pathway C3 convertase.
2.2. Regulation of Complement Activation
Activation and amplification of the complement system induces a powerful inflammatory response, necessitating a regulatory system to avoid damage to host cells. This is achieved by a number of soluble and membrane-bound effector molecules which modulate various critical stages of the pathway, including the widely expressed membrane-bound complement regulatory proteins (mCRPs) (Table 1) and the fluid phase proteins C1 inhibitor, C4b-binding protein, factor H (FH) and factor I (FI) [35,48,49][35][48][49]. The alternative pathway C3 convertase is stabilised when bound by properdin [50].
Table 1.
Membrane-bound complement regulatory proteins.
| Regulator | Alternative Name (s) | Distribution | Function | Reference |
|---|---|---|---|---|
| CD35 | Complement receptor 1 (CR1) | Primarily lymphocytes, erythrocytes, phagocytes, dendritic cells | Cofactor for C3b and C4b degradation by Factor H Accelerates C3 and C5 convertases |
[51,52,53,54,55][51][52][53][54][55] |
| CD46 | Membrane cofactor protein (MCP) | All nucleated cells | Cofactor for C3b and C4b degradation by Factor H | [56,57,58][56][57][58] |
| CD55 | Decay accelerating factor (DAF) | Ubiquitously expressed | Accelerates decay of C3 and C5 convertases | [59,60,61][59][60][61] |
| CD59 | Membrane-inhibitor of reactive lysis (MIRL), MAC inhibitory protein (MAC-IP), Protectin | Ubiquitously expressed | Binds C5b-C9 to prevent polymerization of C9 | [62,63][62][63] |
2.3. Functions of Complement
MAC-induced lysis is the central cytotoxic event resulting from complement cascade activation, however, complement opsonins and anaphylatoxins also contribute to host defence. C3a and C5a exert their biological functions by binding to their respective receptors, the C3a receptor (C3aR) and the C5a receptor 1 (C5aR1/CD88), two G protein-coupled receptors [64,65][64][65]. A second, lesser understood C5aR, C5aR2 (previously C5L2) also exists [66]. In contrast, this 7-transmembrane receptor is uncoupled to G-proteins but is capable of recruiting β-arrestins [67,68,69][67][68][69]. C3a and C5a have been demonstrated to induce chemotaxis of mast cells [70,71][70][71] and eosinophils [72], with C5a also acting as a chemoattractant for macrophages [73], monocytes [74] neutrophils [74,75][74][75], basophils [76] and T and B lymphocytes [77,78][77][78]. Complement opsonins such as the C3 fragments C3b and iC3b aid phagocytosis by allowing recruited phagocytes to adhere to target cells via complement receptor 1 (CR1), complement receptor 3 (CR3), complement receptor 4 (CR4) and complement receptor Ig [4,79][4][79]. The phagocytic response to immune complexes may be enhanced by C5a-mediated upregulation of activating fragment crystallisation (Fc) γ receptors (FcγR) on the surface of phagocytes [24,80][24][80].
Complement cascade components also play key roles in orchestrating adaptive immunity. The complement receptors CR1 and CR2 are essential in the generation of B cell and follicular dendritic cell responses [81]. B cell responses are augmented by the binding of antigen opsonized by C3d to CR2, which leads to enhanced signalling through the B cell receptor and subsequent lowering of the threshold for activation [82,83,84][82][83][84]. In addition, complement components play roles in the priming and differentiation of T cells and provide survival signals to naïve T cells in an autocrine fashion [85,86,87,88][85][86][87][88]. Complement-mediated regulation of immune cells has been demonstrated within the TME [16,89]. The implication of these interactions on the efficacy of cancer treatment will be discussed later in this review[16][89].
References
- Coussens, L.M.; Werb, Z. Inflammation and cancer. Nature 2012, 420, 860–867.
- De Visser, K.E.; Eichten, A.; Coussens, L.M. Paradoxical roles of the immune system during cancer development. Nat. Rev. Cancer 2006, 6, 24–37.
- Reis, E.S.; Mastellos, D.C.; Ricklin, D.; Mantovani, A.; Lambris, J.D. Complement in cancer: Untangling an intricate relationship. Nat. Rev. Immunol. 2018, 18, 5–18.
- Ricklin, D.; Hajishengallis, G.; Yang, K.; Lambris, J.D. Complement: A key system for immune surveillance and homeostasis. Nat. Immunol. 2010, 11, 785–797.
- Carroll, M.C. The complement system in regulation of adaptive immunity. Nat. Immunol. 2004, 5, 981–986.
- Schifferli, J.A.; Ng, Y.C.; Peters, D.K. The role of complement and its receptor in the elimination of immune complexes. N. Engl. J. Med. 1986, 315, 488–495.
- Nozaki, M.; Raisler, B.J.; Sakurai, E.; Sarma, J. V.; Barnum, S.R.; Lambris, J.D.; Chen, Y.; Zhang, K.; Ambati, B.K.; Baffi, J.Z.; et al. Drusen complement components C3a and C5a promote choroidal neovascularization. Proc. Natl. Acad. Sci. USA 2006, 103, 2328–2333.
- Stevens, B.; Allen, N.J.; Vazquez, L.E.; Howell, G.R.; Christopherson, K.S.; Nouri, N.; Micheva, K.D.; Mehalow, A.K.; Huberman, A.D.; Stafford, B.; et al. The Classical Complement Cascade Mediates CNS Synapse Elimination. Cell 2007, 131, 1164–1178.
- Pio, R.; Corrales, L.; Lambris, J.D. The Role of Complement in Tumour Growth. Adv. Exp. Med. Biol. 2014, 772, 229–262.
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The Next Generation. Cell 2011, 144, 646–647.
- Ytting, H.; Christensen, I.J.; Steffensen, R.; Alsner, J.; Thiel, S.; Jensenius, J.C.; Hansen, U.; Nielsen, H.J. Mannan-Binding Lectin (MBL) and MBL-Associated Serine Protease 2 (MASP-2) Genotypes in Colorectal Cancer. Scand. J. Immunol. 2011, 73, 122–127.
- Storm, L.; Christensen, I.J.; Jensenius, J.C.; Nielsen, H.J.; Thiel, S. Evaluation of complement proteins as screening markers for colorectal cancer. Cancer Immunol. Immunother. 2015, 64, 41–50.
- Kesselring, R.; Thiel, A.; Pries, R.; Fichtner-Feigl, S.; Brunner, S.; Seidel, P.; Bruchhage, K.L.; Wollenberg, B. The complement receptors CD46, CD55 and CD59 are regulated by the tumour microenvironment of head and neck cancer to facilitate escape of complement attack. Eur. J. Cancer 2014, 50, 2152–2161.
- Imamura, T.; Yamamoto-Ibusuki, M.; Sueta, A.; Kubo, T.; Irie, A.; Kikuchi, K.; Kariu, T.; Iwase, H. Influence of the C5a–C5a receptor system on breast cancer progression and patient prognosis. Breast Cancer 2016, 23, 876–885.
- Roumenina, L.T.; Daugan, M.V.; Noé, R.; Petitprez, F.; Vano, Y.A.; Sanchez-Salas, R.; Becht, E.; Meilleroux, J.; Clec’h, B. Le; Giraldo, N.A.; et al. Tumor Cells Hijack Macrophage-Produced Complement C1q to Promote Tumor Growth. Cancer Immunol. Res. 2019, 7, 1091–1106.
- Markiewski, M.M.; Deangelis, R.; Benencia, F.; Ricklin-, S.K.; Koutoulaki, A.; Gerard, C.; Coukos, G.; Lambris, J.D. Modulation of the anti-tumor immune response by complement. Nat. Immunol. 2008, 9, 1225–1235.
- Elvington, M.; Melissa Scheiber, Yang, X.; Lyons, K.; Jacqmin, D.; Wadsworth, C.; Marshall, D.; Vanek, K.; Tomlinson, S. Complement dependent modulation of anti-tumor immunity following radiation therapy. Cell Rep. 2014, 8, 818–830.
- Kourtzelis, I.; Rafail, S. The dual role of complement in cancer and its implication in anti-tumor therapy. Ann. Transl. Med. 2016, 4, doi:10.21037/atm.2016.06.26.
- Roumenina, L.T.; Daugan, M.V.; Petitprez, F.; Sautès-Fridman, C.; Fridman, W.H. Context-dependent roles of complement in cancer. Nat. Rev. Cancer. 2019, 19, 698−715, doi:10.1038/s41568-019-0210-0.
- Bordet, J.; Gengou, O. Sur l’existence de substances sensibilisatrices dans la plu- part des serum antimicrobiens. Ann. Inst. Pasteur (Paris) 1901, 15, 289−302.
- Nesargikar, P.N.; Spiller, B.; Chavez, R. The complement system: History, pathways, cascade and inhibitors. Eur. J. Microbiol. Immunol. 2012, 2, 103–111.
- Sarma, J.V.; Ward, P.A. The Complement System. Cell Tissue Res. 2011, 343, 227–235.
- Qu, H.; Ricklin, D.; Lambris, J.D. Recent Developments in Low Molecular Weight Complement Inhibitors. Mol. Immunol. 2009, 47, 185–195.
- Klos, A.; Tenner, A.J.; Johswich, K.O.; Ager, R.R.; Reis, E.S.; Köhl, J. The role of the anaphylatoxins in health and disease. Mol. Immunol. 2009, 46, 2753–2766.
- Kishore, U.; Ghai, R.; Greenhough, T.J.; Shrive, A.K.; Bonifati, D.M.; Gadjeva, M.G.; Waters, P.; Kojouharova, M.S.; Chakraborty, T.; Agrawal, A. Structural and functional anatomy of the globular domain of complement protein C1q. Immunol. Lett. 2004, 95, 113–128.
- Szalai, A.J.; Agrawal, A.; Greenhough, T.J.; Volanakis, J.E. C-Reactive Protein: Structural Biology and Host Defense Function. Clin. Chem. Lab. Med. 1999, 37, 265–270.
- Volanakis, J.E.; Kaplan, M.H. Interaction of C-Reactive Protein Complexes with the Complement system.; I.I. Consumption of Guinea Pig Complement by CRP Complexes: Requirement for Human C1q. J. Immunol. 1974, 113, 9–17.
- Kang, Y.S.; Do, Y.; Lee, H.K.; Park, S.H.; Cheong, C.; Lynch, R.M.; Loeffler, J.M.; Steinman, R.M.; Park, C.G. A Dominant Complement Fixation Pathway for Pneumococcal Polysaccharides Initiated by SIGN-R1 Interacting with C1q. Cell 2006, 125, 47–58.
- Ebenbichler, C.F. Human immunodeficiency virus type 1 activates the classical pathway of complement by direct C1 binding through specific sites in the transmembrane glycoprotein gp41. J. Exp. Med. 1991, 174, 1417–1424.
- Spear, G.T.; Jiang, H.X.; Sullivan, B.L.; Gewurz, H.; Landay, A.L.; Lint, T.F. Direct Binding of Complement Component C1q to Human Immunodeficiency Virus (HIV) and Human T Lymphotrophic Virus-I (HTLV-I) Coinfected Cells. AIDS Res. Hum. Retrovir. 1991, 7, 579–585.
- Thielens, N.M.; Tacnet-Delorme, P.; Arlaud, G.J. Interaction of C1q and Mannan-binding lectin with viruses. Immunobiology 2002, 205, 563–574.
- Holmskov, U.; Thiel, S.; Jensenius, J.C. Collectins and ficolins: Humoral Lectins of the Innate Immune Defense. Annu. Rev. Immunol. 2003, 21, 547–578.
- Thiel, S.; Vorup-Jensen, T.; Stover, C.M.; Schwaeble, W.; Laursen, S.B.; Poulsen, K.; Willis, A.C.; Eggleton, P.; Søren Hansen; Holmskov, U.; et al. A second serine protease associated with mannan-binding lectin that activates complement. Nature 1997, 386, 506–510.
- Megyeri, M.; Harmat, V.; Major, B.; Végh, Á.; Balczer, J.; Héja, D.; Szilágyi, K.; Datz, D.; Pál, G.; Závodszky, P.; et al. Quantitative characterization of the activation steps of mannan-binding lectin (MBL)-associated serine proteases (MASPs) points to the central role of MASP-1 in the initiation of the complement lectin pathway. J. Biol. Chem. 2013, 288, 8922–8934.
- Merle, N.S.; Church, S.E.; Fremeaux-Bacchi, V.; Roumenina, L.T. Complement system part I—Molecular mechanisms of activation and regulation. Front. Immunol. 2015, 6, 1–30.
- Pangburn, M.K.; Schreiber, R.D.; Müller-Eberhard, H.J. Formation of the initial C3 convertase of the alternative complement pathway. Acquisition of C3b-like activities by spontaneous hydrolysis of the putative thioester in native C3. J. Exp. Med. 1981, 154, 856–867.
- Bexborn, F.; Andersson, P.O.; Chen, H.; Nilsson, B.; Ekdahl, K.N. The Tick-Over Theory Revisited: Formation and Regulation of the soluble Alternative Complement C3 Convertase (C3(H2O)Bb). Mol. Immunol. 2008, 45, 2370–2379.
- Reid, K.B.M.; Porter, R.R. The Proteolytic Activation Systems of Complement. Annu. Rev. Biochem. 1981, 50, 434–464.
- Ganter, M.T.; Brohi, K.; Cohen, M.J.; Shaffer, L.A.; Walsh, M.C.; Stahl, G.L.; Pittet, J.F. Role of the alternative pathway in the early complement activation following major trauma. Shock 2007, 28, 29–34.
- Spitzer, D.; Mitchell, L.M.; Atkinson, J.P.; Hourcade, D.E. Properdin Can Initiate Complement Activation by Binding Specific Target Surfaces and Providing a Platform for De Novo Convertase Assembly. J. Immunol. 2007, 179, 2600–2608.
- Hourcade, D.E. The role of properdin in the assembly of the alternative pathway C3 convertases of complement. J. Biol. Chem. 2006, 281, 2128–2132.
- Harboe, M.; Mollnes, T.E. The alternative complement pathway revisited. J. Cell Mol. Med. 2008, 12, 1074–1084.
- Walport, M.J. Complement. First of Two Parts. N. Engl. J. Med. 2001, 344, 1058–1066.
- Ehrnthaller, C.; Ignatius, A.; Gebhard, F.; Huber-lang, M. New Insights of an Old Defense System: Structure, Function and Clinical Relevance of the Complement System. Mol. Med. 2011, 17, 317–329.
- Koski, C.L.; Ramm, L.E.; Hammer, C.H.; Mayer, M.M.; Shin, M.L. Cytolysis of nucleated cells by complement: Cell death displays multi-hit characteristics. Proc. Natl. Acad. Sci. USA 1983, 80, 3816–3820.
- Morgan, B.P. Complement membrane attack on nucleated cells: Resistance, recovery and non-lethal effects. Biochem. J. 1989, 264, 1–14.
- Tegla, C.A.; Cudrici, C.; Patel, S.; Trippe, R.; Rus, V.; Niculescu, F.; Rus, H. Membrane attack by complement: The assembly and biology of terminal complement complexes. Immunol. Res. 2011, 51, 45–60.
- Kim, D.D.; Song, W. Membrane complement regulatory proteins. Clin. Immunol. 2006, 118, 127–136.
- Schmidt, C.Q.; Lambris, J.D.; Ricklin, D. Protection of host cells by complement regulators. Immunol. Rev. 2016, 274, 152–171.
- Fearon, D.T.; Austen, K.F. Properdin: Binding to C3b and stabilization of the C3b dependent C3 convertase. J. Exp. Med. 1975, 142, 856–863.
- Fearon, D.T. Regulation of the amplification C3 convertase of human complement by an inhibitory protein isolated from human erythrocyte membrane. Proc. Natl. Acad. Sci. USA 1979, 76, 5867–5871.
- Fearon, D.T. Identification of the membrane glycoprotein that is the C3b receptor of the human erythrocyte, polymorphonuclear leukocyte, B lymphocyte, and monocyte. J. Exp. Med. 1980, 152, 20–30.
- Iida, K.; Nussenzweig, V. Complement receptor is an inhibitor of the complement cascade. J. Exp. Med. 1981, 153, 1138–1150.
- Medicus, R.G.; Melamed, J.; Arnaout, M.A. Role of human factor I and C3b receptor in the cleavage of surface‐bound C3bi molecules. Eur. J. Immunol. 1983, 13, 465–470.
- Edward Medof, M.; Iida, K.; Mold, C.; Nussenzweig, V. Unique role of the complement receptor CR1 in the degradation of C3b associated with immune complexes. J. Exp. Med. 1982, 156, 1739–1754.
- Seya, T.; Turner, J.R.; Atkinson, J.P. Purification and characterization of a membrane protein (gp45-70) that is a cofactor for cleavage of C3B and C4B. J. Exp. Med. 1986, 163, 837–855.
- Seya, T.; Ballard, L.L.; Bora, N.S.; Kumar, V.; Cui, W.; Atkinson, J.P. Distribution of membrane cofactor protein of complement on human peripheral blood cells. An altered form is found on granulocytes. Eur. J. Immunol. 1988, 18, 1289–1294.
- Liszewski, M.K.; Post, T.W.; Atkinson, J.P. Membrane cofactor protein (MCP or CD46): Newest member of the regulators of complement activation gene cluster. Annu. Rev. Immunol. 1991, 9, 431–455.
- Nicholson-Weller, A.; Burge, J.; Fearon, D.T.; Weller, P.F.; Austen, K.F. Isolation of a human erythrocyte membrane glycoprotein with decay-accelerating activity for C3 convertases of the complement system . J. Immunol. 1982, 129, 184–189.
- Medof, B.Y.M.E.; Kinoshita, T.; Nussenzweig, V. Inhibition of Complement Activation on the Surface of Cells After of Incorporation of Decay-Accelerating Factor (DAF) Into Their Membranes. J. Exp. Med. 1984, 160, 1558–1578.
- Kinoshita, T.; Medof, M.E.; Nussenzweig, V. Endogenous association of decay-accelerating factor ( DAF ) with C4b and C3b on cell membranes. J. Immunol. 1986, 136, 3390–3395.
- Meri, S.; Morgan, B.P.; Davies, A.; Daniels, R.H.; Olavesen, M.G.; Waldmann, H.; Lachmann, P.J. Human protectin (CD59), an 18,000-20,000 MW complement lysis restricting factor, inhibits C5b-8 catalysed insertion of C9 into lipid bilayers. Immunology 1990, 71, 1–9.
- Ninomiya, H.; Sims, P.J. The human complement regulatory protein CD59 binds to the α-chain of C8 and to the “b” domain of C9. J. Biol. Chem. 1992, 267, 13675–13680.
- Ames, R.S.; Li, Y.; Sarau, H.M.; Nuthulaganti, P.; Foley, J.J.; Ellis, C.; Zeng, Z.; Su, K.; Jurewicz, A.J.; Hertzberg, R.P.; et al. Molecular cloning and characterization of the human anaphylatoxin C3a receptor. J. Biol. Chem. 1996, 271, 20231–20234.
- Gerard, N.P.; Gerard, C. The chemotactic receptor for human C5a anaphylatoxin. Nature 1991, 354, 614–617.
- Ohno, M.; Hirata, T.; Enomoto, M.; Araki, T.; Ishimaru, H.; Takahashi, T.A. A putative chemoattractant receptor, C5L2, is expressed in granulocyte and immature dendritic cells, but not in mature dendritic cells. Mol. Immunol. 2000, 37, 407–412.
- Okinaga, S.; Slattery, D.; Humbles, A.; Zsengeller, Z.; Morteau, O.; Kinrade, M.B.; Brodbeck, R.M.; Krause, J.E.; Choe, H.-R.; Gerard, N.P.; et al. C5L2, a nonsignaling C5A binding protein. Biochemistry 2003, 42, 9406–9415.
- Van Lith, L.H. c.; Oosterom, J.; Van Elsas, A.; Zaman, G.J.R. C5a-Stimulated Recruitment of ß-Arrestin2 to the Nonsignaling 7-Transmembrane Decoy Receptor C5L2. J. Biomol. Screen. 2009, 14, 1067–1075.
- Croker, D.E.; Halai, R.; Kaeslin, G.; Wende, E.; Fehlhaber, B.; Klos, A.; Monk, P.N.; Cooper, M.A. C5a2 can modulate ERK1/2 signaling in macrophages via heteromer formation with C5a1 and β-arrestin recruitment. Immunol. Cell Biol. 2014, 92, 631–639, doi:10.1038/icb.2014.32.
- Hartmann, K.; Henz, B.M.; Krüger-Krasagakes, S.; Köhl, J.; Burger, R.; Gurtl, S.; Haase, I.; Lippert, U.; Zuberbier, T. C3a and C5a stimulate chemotaxis of human mast cells. Blood 1997, 89, 2863–2870.
- Nilsson, C.; Johnell, M.; Hammer, C.H.; Tiffany, H.L.; Metcalfe, D.D.; Siegbahn, A.; Murphyt, P.M. C3a and C5a Are Chemotaxins for Human Mast Cells and Act Through Distinct Receptors via a Pertussis Toxin-Sensitive Signal Transduction Pathway. J. Immunol. 1996, 157, 1693–1998.
- Daffern, B.P.J.; Pfeifer, P.H.; Ember, J.A.; Hugli, T.E. C3a is a Chemotaxin for Human Eosinophils but Not for Neutrophils. I. C3a Stimulation of Neutrophils Is Secondary to Eosinophil Activation. J. Exp. Med., 1995, 181, 2119–2127.
- Aksamit, R.R.; Falk, W.; Leonard, E.J. Chemotaxis by mouse macrophage cell lines. J. Immunol. 1981, 126, 2194–2199.
- Yancey, K.B.; Lawley, T.J.; Dersookian, M.; S., B.; Harvath, L. Analysis of the Interaction of Human C5a and C5a des Arg with Human Monocytes and Neutrophils: Flow Cytometric and Chemotaxis Studies. J. Invest. Dermatol. 1989, 92, 184–189.
- Ehrengruber, M.U.; Geiser, T.; Deranleau, D.A. Activation of human neutrophils by C3a and C5a. FEBS Lett. 1994, 346, 181–184.
- Lett-Brown, M.A.; Leonard, E.J. Histamine-Induced Inhibition of Normal Human Basophil Chemotaxis to C5a. J. Immunol. 1977, 118, 815–818.
- Nataf, S.; Davoust, N.; Ames, R.S.; Barnum, S.R. Human T Cells Express the C5a Receptor and Are Chemoattracted to C5a. J. Immunol. 1999, 162, 4018–4023.
- Ottonello, L.; Corcione, A.; Tortolina, G.; Albesiano, E.; Favre, A.; Agostino, R.D.; Malavasi, F.; Pistoia, V.; Dallegri, F.; Favre, A.; et al. rC5a Directs the In Vitro Migration of Human Memory and Naive Tonsillar B Lymphocytes: Implications for B Cell Trafficking in Secondary Lymphoid Tissues. J. Immunol. 1999, 162, 6510–6517.
- van Lookeren Campagne, M.; Wiesmann, C.; Brown, E.J. Macrophage complement receptors and pathogen clearance. Cell. Microbiol. 2007, 9, 2095–2102.
- Karsten, C.M.; Köhl, J. The immunoglobulin, IgG Fc receptor and complement triangle in autoimmune diseases. Immunobiology 2012, 217, 1067–1079.
- Fang, Y.; Xu, C.; Fu, Y.X.; Holers, V.M.; Molina, H. Expression of Complement Receptors 1 and 2 on Follicular Dendritic Cells Is Necessary for the Generation of a Strong Antigen-Specific IgG Response. J. Immunol. 1998, 160, 5273–5279.
- Carter, R.H.; Fearon, D.T. CD19: lowering the threshold for antigen receptor stimulation of B lymphocytes. Science 1992, 256, 105–107.
- Cherukuri, A.; Cheng, P.C.; Pierce, S.K. The Role of the CD19/CD21 Complex in B Cell Processing and Presentation of Complement-Tagged Antigens. J. Immunol. 2001, 167, 163–172.
- Matsumoto, A.K.; Kopicky-Burd, J.; Carter, R.H.; Tuveson, D.A.; Tedder, T.F.; Fearon, D.T. Intersection of the Complement and Immune Systems: A Signal Transduction Complex of the B Lymphocyte-containing Complement Receptor Type 2 and CD19. J. Exp. Med. 1991, 173, 55–64.
- Strainic, M.G.; Liu, J.; Huang, D.; An, F.; Lalli, P.N.; Muqim, N.; Shapiro, V.S.; Dubyak, G.R.; Heeger, P.S.; Medof, M.E. Locally Produced Complement Fragments C5a and C3a Provide Both Costimulatory and Survival Sinals to naive CD4+ T Cells. Immunity 2008, 28, 425–435.
- Lalli, P.N.; Strainic, M.G.; Yang, M.; Lin, F.; Medof, M.E.; Heeger, P.S. Locally produced C5a binds to T cell expressed C5aR to enhance effector T-cell expansion by limiting antigen-induced apoptosis. Blood 2008, 112, 1759–1766.
- Strainic, M.G.; Shevach, E.M.; An, F.; Lin, F.; Medof, M.E. Absent C3a and C5a receptor signaling into CD4+T cells enables auto-inductive TGF-β1 signaling and induction of Foxp3+T regulatory cells. Nat. Immunol. 2013, 14, 162–171.
- Lalli, P.N.; Strainic, M.G.; Lin, F.; Medof, M.E.; Heeger, P.S. Decay Accelerating Factor Can Control T Cell Differentiation into IFN-γ-Producing Effector Cells via Regulating Local C5a-Induced IL-12 Production. J. Immunol. 2007, 179, 5793–5802.
- Vadrevu, S.K.; Chintala, N.K.; Sharma, S.K.; Sharma, P.; Cleveland, C.; Riediger, L.; Manne, S.; Fairlie, D.P.; Gorczyca, W.; Almanza, O.; et al. Complement C5a receptor facilitates cancer metastasis by altering t-cell responses in the metastatic niche. Cancer Res. 2014, 74, 3454–3465.