Carotenoids are an essential group of compounds that may be obtained by microbiological synthesis. They are instrumental in various areas of industry, medicine, agriculture, and ecology.
- yeast
- carotenoids
- pigments
1. Introduction
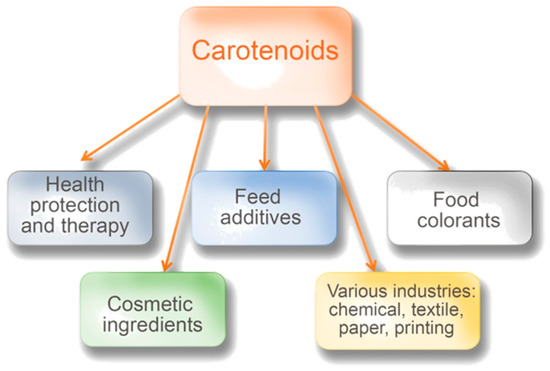
Carotenoids are an essential group of compounds that can be synthesized by some bacteria, yeasts, and molds. They are largely produced by plants, especially green leafy plants, for which some of them play a crucial role in photosynthesis [1,2,3,4][1][2][3][4]. In this process, they help absorb light but also play an important role in removing excess solar energy [5]. In the case of microorganisms, the main role of carotenoids is to protect cells against the negative influence of reactive forms of oxygen and radiation [6]. Carotenoids have applications in various areas of industry, medicine, agriculture, and ecology. A lot of information has been accumulated during the last decades about their possible health-protecting effects [7,8,9,10,11][7][8][9][10][11]. It is known that carotenoids cannot be synthesized in humans and animals. Therefore, people and animals need to obtain them from their diet [4,12][4][12]. Carotenoids may provide cosmetic benefits [7,10,13][7][10][13]. Moreover, their great importance in food production, as natural colorants, is well-known (Figure 1). The global market of carotenoids grew very promptly: In 2017, it reached the value of 1.5 billion USD. Based on the expectations of experts, it should reach $2.0 billion by 2022, at a compound annual growth rate of 5.7% for the period of 2017–2022 [14].
Carotenoids are lipid-soluble, mainly terpenoid pigments of 40 carbon atoms. It is thought that the most important structural characteristic of carotenoids is their conjugated double bonds (CDBs) that are largely responsible for their physicochemical properties. For instance, CDBs are responsible for the color of most carotenoids. At least seven CDBs are necessary for obtaining a colored carotenoid [10]. Carotenoids can be divided into two groups. One of them is “oxygen-free carotenes”, e.g., α-carotene, β-carotene, ¥-carotene, lycopene, and torulene. The second group is “oxygen-containing xanthophylls”, e.g., astaxanthin, lutein, zeaxanthin, β-cryptoxanthin, fucoxanthin, and canthaxanthin [4,15][4][15]. Carotenoids can also be divided into provitamin A and non-provitamin A compounds [16]. The major provitamin A carotenoids are β-carotene, α-carotene, and β-cryptoxanthin. The carotenoids that are mainly studied so far are β-carotene, lycopene, astaxanthin, lutein, and zeaxanthin [17].
It is well-known that filamentous fungi and yeasts may produce, besides carotenoids, a lot of other various pigments, including melanins, flavins, phenazines, quinones, and others. One rather new pigment being researched is the red pigment accumulated by Saccharomyces cerevisiae mutants for ADE1 and ADE2, the product of the polymerization of 1-(5′-phosphoribosyl)-5-aminoimidazole containing several amino acid residues. This red pigment is a mixture of polymers containing a different number of monomers (4–10) and is characterized by a molecular weight from 2 to 10 kDa [17,18,19,20][17][18][19][20].
2. Characteristics of Some Fungal Carotenoids
The group of yeast that can synthesize carotenoids includes Phaffia rhodozyma (and its teleomorph Xanthophyllomyces dendrorhous) and species of the genera Rhodosporidium, Rhodotorula, Sporobolomyces, and Sporidiobolus [21,22,23][21][22][23]. Among the molds, the Blakeslea trispora species is of the greatest importance [24]. The biosynthesis of carotenoids in fungal cells begins with the conversion of acetyl-CoA, which is formed in the process of β-oxidation of fatty acids in the mitochondria. According to the pathway of mevalonic acid, several biochemical reactions catalyzed by specific reductases, kinases, and decarboxylases produce a five-carbon carotenoid precursor, isopentenyl pyrophosphate (IPP). The addition reactions of three IPPs lead to the formation of geranyl–geranyl pyrophosphate (GGPP), with 20 carbon atoms per molecule. The condensation of the two GGPP particles, catalyzed by phytoene synthase, produces phytoene (C40). It is a precursor to lycopene biosynthesis. Depending on the type of microorganisms, lycopene can be next transformed into β-carotene, γ-carotene, torulene, lutein, torularhodin, zeaxanthin, and astaxanthin [25].
3. Carotenoids and Human Health
It is well-known that carotenoids are compounds that are very important for human health. They can prevent a deficiency in vitamin A, which is known as the essential compound for the promotion of growth, embryonal development, and visual function. The lipophilicity of carotenoids determines their subcellular distribution; they are enriched in membranes and other lipophilic compartments, i.e., lipid droplets [16]. It is supposed that carotenoids in membranes can protect them as antioxidants. Besides that, polar carotenoids can regulate membrane fluidity [21,22][21][22]. One of their functions is linked to the protection of our vision. The deficiency of carotenoids can lead to blindness, and as it was reviewed in the literature it is a serious problem for children, especially in developing countries [23,46][23][26]. Carotenoids are vital for the protection of the retina by preventing cataracts and age-related macular degeneration [46,47,48][26][27][28]. There is definite evidence that shows the efficiency for eye health of lutein and zeaxanthin. They may reduce the risk for age-related macular eye diseases and lead to the improvement of visual performance that also includes positive effects, such as contrast sensitivity, glare tolerance, and photo-stress recovery [49][29].
Torularhodin is a carotenoid produced mainly by the yeast genera Rhodotorula and Sporobolomyces; it has strong antimicrobial properties and may become a new natural antibiotic [50,51,52,53][30][31][32][33]. The antimicrobial properties of torularhodin can also be used in the production of films for coating medical implants [54,55][34][35].
The efficiency of carotenoids’ use is known for the protection and therapy of various chronic diseases. chronic diseases. They exhibit an anti-inflammatory property and may activate the immune response of an organism [56][36]. It was shown that the use of lycopene-enriched foods might decrease the risk of developing atherosclerosis and other cardiovascular diseases [57,58,59][37][38][39]. Such beneficial results are most likely linked to the ability of lycopene to reduce systemic and high-density lipoprotein-associated inflammation and to modulate high-density lipoprotein functionality [60][40]. It was shown that supplementation with lycopene significantly decreased systolic blood pressures [59,61][39][41]. Astaxanthin has also been reported to exert a preventive action against atherosclerotic cardiovascular diseases by the reduction of oxidative stress and inflammation and the enhancement of lipid metabolism and glucose metabolism [62][42]. Supplying the body with astaxanthin allows us to effectively reduce the negative effects resulting from the oxidation and degradation of cellular elements. Another study revealed that lycopene might limit the release of proinflammatory cytokines and chemokines [63][43]. One study also speculated that lycopene might affect the immune functions modulating the cellular redox environment and cell-to-cell interactions and influence anti-inflammatory transcription factors, such as peroxisome-proliferator-activated receptor [64][44]. Several results, summarized by Rao and Rao [59][39], reported the involvement of lycopene and β-carotene in bone health and in preventing or decreasing the risk of osteoporosis. Such a positive effect of lycopene in decreasing osteoporosis risk was also shown in postmenopausal women [65][45]. Similar effects were also described for β-cryptoxanthin [49][29]. Lycopene consumption was demonstrated to improve bone strength, by reducing bone resorption, and to protect from type 2 diabetes, by enhancing glucose homeostasis [66,67,68][46][47][48].
The role of various carotenoids in the prevention of other chronic diseases was also studied [59][39]. Moreover, the use of lycopene in the cases of male infertility led to the improvements of sperm motility, sperm motility index, sperm morphology, and functional sperm concentration, and finally resulted in a 36% increase of successful pregnancies [59][39]. The possible use of lycopene in recovering the cases of alcohol-induced liver injury was also suggested [21]. Carotenoids might have beneficial effects on weight management and obesity [49,69][29][49]. It is expected that future studies could reveal a positive role of carotenoids in the treatments of other diseases, i.e., skin disorders, rheumatoid arthritis, periodontal diseases, and others [49,70][29][50].
β-Carotene and lutein have positive effects on cognitive performance [49][29]. The putative positive role of lycopene in the treatments of neurodegenerative diseases, including Alzheimer’s disease, was also studied [60,71][40][51]. It was thought that lutein is linked to the possible control of inflammation-related neurodegenerative disorders [72][52], while torularhodin can be used as a neuroprotective agent against H2O2-induced oxidative stress, due to its strong antioxidant activity [73][53]. Lycopene exhibited protection against amyotrophic lateral sclerosis disorder in humans [59,74][39][54].
Interesting information was received, in recent years, regarding the possible medical application of a red pigment that accumulated in S. cerevisiae mutants. It is known that “conformational diseases” in humans and animals are linked to abnormal aggregation of proteins and the formation of amyloid fibrils. The red pigment accumulated in S. cerevisiae mutants for ADE1 and ADE2 can bind amyloid fibrils and disturb their interactions with chaperones that, in turn, lead to the inhibition of prion “multiplication” and amyloid fibril formation [75,76,77][55][56][57]. It was also shown that yeast mutants, which accumulate this pigment, had lower amyloid content than wild-type parental strains. It was shown that this red pigment accumulation reduced cloned human amyloid-β aggregation. The conclusion was made that red yeast pigment has potential importance in therapy for Alzheimer’s and Parkinson’s diseases [19,20][19][20].
Carotenoids have characteristics of antioxidants [78,79,80,81][58][59][60][61]. They quench 1O2 and increase the levels of glutathione and glutathione peroxidase [4,82,83][4][62][63]. β-Carotene can be used for sun protection and sunburn prevention [16,84][16][64]. Carotenoids are efficient blue-light filters; they protect against photo-oxidative damages lipids, proteins, and DNA, thus preventing premature ageing of the skin and skin cancer [16,84,85,86][16][64][65][66]. It was also suggested that astaxanthin might be used as a potential anti-ageing agent [87][67]. β-Carotene reduces the risk of developing neoplastic diseases, and also inhibits the promotion and progression of neoplasms.
Very promising findings were also obtained on the putative efficiency of using carotenoids against some types of cancer [46,88][26][68]. The anticancer activity of some carotenoids, i.e., α-carotene, β-carotene, lycopene, torulene, torularhodin, and some others, was studied regarding prostate, breast, colon, lung, oral, gastric, and skin cancers, in addition to hepatoma, leukemia, uveal melanoma, etc. [4,52,53,66,88,89,90,91,92,93,94,95,96][4][32][33][46][68][69][70][71][72][73][74][75][76]. Synergistic inhibition of prostate and breast cancer cell growth was evident under the influence of combinations of low concentrations of various carotenoids [97][77]. The use of reporter gene assays of the transcriptional activity of the androgen receptor in hormone-dependent prostate cancer cells and the electrophile/antioxidant response element (EpRE/ARE) transcription system enabled the observation of combinations of several carotenoids (e.g., lycopene, phytoene, and phytofluene) to synergistically inhibit the androgen receptor activity and activate the EpRE/ARE system and suggested their use in the therapy and prevention of this type of cancer [97][77]. In the experiments performed by Prakash et al. [98][78], estrogen-receptor (ER) positive MCF-7 and ER-negative Hs578T and MDA-MB-231 human breast cancer cells were treated with carotenoids. Among them, β-carotene significantly reduced the growth of MCF-7 and Hs578T cells, and lycopene inhibited the growth of MCF-7 and MDA-MB-231 cells. Similar effects were also shown for astaxanthin [62][42]. Authors concluded that carotenoids inhibit the growth of both studied breast cancer cell lines, indicating that estrogen receptor status is an important factor for the responsiveness of breast cancer cells to carotenoid treatments [98][78]. The use of food rich in various carotenoids was found to decrease the risk of lung and stomach cancers [81,99,100][61][79][80]; although, in the case of lung cancer, negative results were obtained for β-carotene for smokers and asbestos workers. In these cases, β-carotene supplementation was associated with an increased risk of lung and gastric cancers [49,101][29][81]. It is supposed that the cancer-preventive effects exhibited by various carotenoids might also be linked to their induction and stimulation of intercellular communications via gap junctions, which are important for the regulation of cell growth, differentiation, and apoptosis [21]. More recently, lycopene was found to inhibit tumor metastasis by slowing down cell-cycle progression and inhibiting the proliferation of diverse cancer cell lines [66][46]. A detailed description of the different effects and mechanisms of anticancer activity of carotenoids (cell-cycle arrest, apoptosis-inducing effect, and anti-metastasis effect) is reported in some recent reviews [4,102,103][4][82][83].
Carotenoids may act as chemoprotective agents against cellular mutagenesis and malignant transformation [79,81,104,105,106][59][61][84][85][86]. Protective effects expressed by β-carotene and other carotenoids were demonstrated against the mutagenic potential of 8-methoxypsoralen, cyclophosphamide, 1-methyl-3-nitro-1-nitrosoguanidine, benzo(α)pyrene, quinolones, and ultraviolet light, using Salmonella typhimurium as a cell model system [81,107,108,109,110][61][87][88][89][90]. β-Carotene and other carotenoids (canthaxanthin, α-carotene, and lycopene) can inhibit malignant transformation induced by 3-methylcholanthrene, or X-ray treatment in the fibroblast cell line [81,111,112][61][91][92].
Antioxidant and anti-ageing effects of astaxanthin led to its wide use in cosmetics [113][93]. Besides all of these examples of the positive effects of carotenoids on human health, there are also data that β-carotene and astaxanthin may have immunoprotective effects, whereas lutein can prevent oxidative stress in eye tissues, as well as has antiviral activity against hepatitis B virus [113][93]. Figure 2 briefly presents the possible positive effects of carotenoids for human health described above.
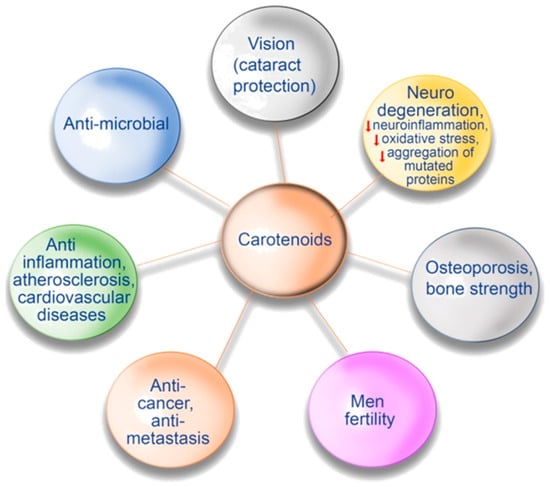
Figure 2. Possible positive effects of carotenoids on human health.
4. Carotenoids and Other Microbial Pigments as Feed Additives and Colorants
Carotenoids are widely used in salmon and trout farming and in the poultry and food industry as feed additives and natural food colorants, which can give from yellow to red colors [7,114,115,116][7][94][95][96]. From an economic viewpoint, astaxanthin is the third most important carotenoid after β-carotene and lutein, due to its importance in aquaculture, and the chemical, pharmaceutical, and food industries [3]. In salmon and trout farming, it is widely used as a pigment for fish meat. Feed supplementation of carotenoids essentially improves the health of poultry birds and enhances the quality of eggs and meat. Carotenoids are very important for the pigmentation of egg yolk, skin, legs, beak, comb, feather, and fat. The use of carotenoids as alternative feed ingredients gives the possibility to replace synthetic medicine and nutrients in poultry industry [117][97].
Many yeasts belonging to different genera have been extensively studied during the last decades as potential efficient producers of various pigments (especially of mixtures of carotenoids). Among them, the most forthcoming is Ph. rhodozyma [118,119][98][99]. The production of astaxanthin has been scaled-up to the industrial level in the last decades [116,120,121][96][100][101]. On the other hand, the yellow carotenoid pigment zeaxanthin can be used as an additive in poultry, as well as in the cosmetics and food industries. Canthaxanthin is another carotenoid pigment that is already used in aquafeed for farmed salmonids [122][102]. Besides these food-related applications of carotenoids, they may serve as alternative coloring agents that are in demand in different industries, such as the textile, plastic, paint, paper, and printing industries [122,123,124,125][102][103][104][105].
The analysis of food consumer requirements revealed a growing rejection of synthetic food dyes during the last decade. The use of some synthetic colorants in food and cosmetic processing has recently been banned due to their hyperallerginicity, carcinogenicity, and other toxicological problems [122,123][102][103]. For example, the astaxanthin produced chemically is not approved for human consumption, due to the presence of by-products [126][106]. Correspondingly, a growing demand for dyes of natural origin is becoming increasingly more popular. It is well-known that natural coloring agents can be extracted from various plants, algae, and microorganisms (i.e., bacteria, yeasts, and fungi), which can produce various pigments [127][107]. Some food-grade microbial pigments are already produced biotechnologically. Among them is the hydroxyanthraquinoid pigment Arpink red, which is produced by a strain of Penicillium oxalicum var. armeniaca isolated from soil by the Czech company Ascolor Biotech s.r.o. The patent covering Arpink Red also claims its anticancer effects for applications in the food and pharmaceutical fields [127][107]. Another example is the yellow vitamin riboflavin (vitamin B2). It can be produced by the yeast species Meyerozyma (formerly Candida) guilliermondii or Debaryomyces subglobosus and by the dimorphic fungus Eremothecium ashbyi (and its heterotypic synonym Ashbya gossypii); the latter is used for industrial-scale production [126,128][106][108].
Recent studies explored the possibility of replacing the use of yellow pigments from the fungus Monascus sp. (which are not approved for the use in EU and USA, because of the risk of possible contamination by the nephrotoxic and hepatotoxic metabolite citrinin), with similar pigments produced by non-mycotoxigenic strains of the fungal genus Talaromyces [124,128,129,130,131,132][104][108][109][110][111][112]. Marine fungi are also studied as promising sources of novel pigments [128][108]. New findings in this area that give new possibilities for modern biotechnology are described in detail in the review by Dufosse et al. [128][108].
References
- Dufosse, L.; Galaup, P.; Yaron, A.; Arad, S.M.; Blanc, P.; Chidambara Murthy, K.N.; Ravishankar, G.A. Microorganisms and microalgae as sources of pigments for food use: A scientific oddity or an industrial reality? Trends Food Sci. Technol. 2005, 16, 389–406.
- Das, A.; Yoon, S.H.; Lee, S.H.; Kim, J.Y.; Oh, D.K.; Kim, S.W. An update on microbial carotenoid production. Application of recent metabolic engineering tools. Appl. Microbiol. Biotechnol. 2007, 77, 505–512.
- Mata-Gomez, L.C.; Montanez, J.C.; Mendez-Zavala, A.; Aguilar, C.N. Biotechnological production of carotenoids by yeasts: An overview. Microb. Cell Factor 2014, 13, 12.
- Niranjana, R.; Gayathri, R.; Mol, S.N.; Sugawara, T.; Hirata, T.; Miyashita, K.; Ganesan, P. Carotenoids modulate the hallmarks of cancer cells. J. Funct. Foods 2015, 18, 968–985.
- Riso, P.; Brusamolino, A.; Scalfi, L.; Porrini, M. Bioavailability of carotenoids from spinach and tomatoes. Nutr. Metab. Cardiovasc. Dis. 2004, 14, 150–156.
- Di Mascio, P.; Murphy, M.E.; Sies, H. Antioxidant defense systems: The role of carotenoids, tocopherols, and thiols. Am. J. Clin. Nutr. 1991, 53, 194S–200S.
- Rodriguez-Concepcion, M.; Avalos, J.; Bonet, M.L.; Boronat, A.; Gomez-Gomez, L.; Hornero-Mendez, D.; Limon, M.C.; Meléndez-Martínez, A.J.; Olmedilla-Alonso, B.; Palou, A.; et al. A global perspective on carotenoids: Metabolism, biotechnology, and benefits for nutrition and health. Prog. Lipid Res. 2018, 70, 62–93.
- Meléndez-Martínez, A.J. An overview of carotenoids, apocarotenoids and vitamin A in agro-food, nutrition, health and disease. Mol. Nutrit. Food Res. 2019, 63, 1801045.
- Meléndez-Martínez, A.J.; Stinco, C.M.; Mapelli-Brahm, P. Skin carotenoids in public health and nutricosmetics. The emerging roles and applications of the UV light-absorbing colourless carotenoids phytoene and phytofluene. Nutrients 2019, 11, 1093.
- Mapelli-Brahm, P.; Barba, F.J.; Remize, F.; Garcia, C.; Fessard, A.; Khaneghah, A.M.; Sant’Ana, A.S.; Lorenzoe, J.M.; Montesano, D.; Meléndez-Martínez, A.J. The impact of fermentation processes on the production, retention and bioavailability of carotenoids: An overview. Trends Food Sci. Technol. 2020, 99, 389–401.
- Britton, G. Carotenoid research: History and new perspectives for chemistry in biological systems. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2020, 1865, 158699.
- Woodside, J.V.; McGrath, A.J.; Lyner, N.; McKinley, M.C. Carotenoids and health in older people. Maturitas 2015, 80, 63–68.
- Meléndez-Martínez, A.J.; Mapelli-Brahm, P.; Stinco, C.M. The colourless carotenoids phytoene and phytofluene: From dietary sources to their usefulness for the functional foods and nutricosmetics industries. J. Food Compos. Anal. 2018, 67, 91–103.
- BCC Research. The Global Market for Carotenoids. 2018. Available online: https://www.bccresearch.com/market-research/food-and-beverage/the-global-market-for-carotenoids.html (accessed on 20 December 2020).
- Aziz, E.; Batool, R.; Akhtar, W.; Rehman, S.; Shahzad, T.; Malik, A.; Shariati, M.A.; Laishevtcev, A.; Plygun, S.; Heydari, M.; et al. Xanthophyll: Health benefits and therapeutic insights. Life Sci. 2020, 240, 117104.
- Stahl, W.; Sies, H. Bioactivity and protective effects of natural carotenoids. Biochim. Biophys. Acta 2005, 1740, 101–107.
- Fernandes, A.S.; do Nascimento, T.C.; Jacob-Lopes, E.; De Rosso, V.V.; Zepka, L.Q. Introductory Chapter: Carotenoids—A brief overview on its structure, biosynthesis, synthesis, and applications. Prog. Carotenoid Res. 2018, 1–16.
- Amen, T.R.; Mikhailova, E.V.; Alenin, V.V.; Artyomov, A.V.; Dementyev, P.A.; Khodorkovskii, M.A.; Artamonova, T.O.; Kuznetsova, I.M.; Soidla, T.R.; Nevzglyadova, O.V. Structural and functional characteristics of various forms of red pigment of yeast Saccharomyces cerevisiae and its synthetic analog. Cell Tissue Biol. 2013, 7, 86–94.
- Nevzglyadova, O.V.; Mikhailova, E.V.; Amen, T.R.; Zenin, V.V.; Artemov, A.V.; Kostyleva, E.I.; Mezhenskaya, D.A.; Rodin, D.I.; Saifitdinova, A.F.; Khodorkovskii, M.A.; et al. Yeast red pigment modifies amyloid beta growth in Alzheimer disease models in both Saccharomyces cerevisiae and Drosophila melanogaster. Amyloid 2015, 22, 100–111.
- Nevzglyadova, O.V.; Mikhailova, E.V.; Artemov, A.V.; Ozerova, Y.E.; Ivanova, P.A.; Golomidov, I.M.; Bolshakova, O.I.; Zenin, V.V.; Kostyleva, E.I.; Soidla, T.R.; et al. Yeast red pigment modifies cloned human α-synuclein pathogenesis in Parkinson disease models in Saccharomyces cerevisiae and Drosophila melanogaster. Neurochem. Intern. 2018, 120, 172–181.
- Tapiero, H.; Townsend, D.M.; Tew, K.D. The role of carotenoids in the prevention of human pathologies. Biomed. Pharmacother. 2004, 58, 100–110.
- Seel, W.; Baust, D.; Sons, D.; Albers, M.; Etzbach, L.; Fuss, J.; Lipski, A. Carotenoids are used as regulators for membrane fluidity by Staphylococcus xylosus. Sci. Rep. 2020, 10, 330.
- Black, R.E.; Allen, L.H.; Bhutta, Z.A.; Caulfield, L.E.; De Onis, M.; Ezzati, M.; Mathers, C.; Rivera, J. Maternal and child undernutrition: Global and regional exposures and health consequences. Lancet 2008, 371, 243–260.
- Metličar, V.; Vovk, I.; Albreht, A. Japanese and Bohemian Knotweeds as Sustainable Sources of Carotenoids. Plants 2019, 8, 384.
- Mussagy, C.U.; Winterburn, J.; Santos-Ebinuma, V.C.; Pereira, J.F.B. Production and extraction of carotenoids produced by microorganisms. Appl. Microbiol. Biotechnol. 2019, 103, 1095–1114.
- Sandmann, G. Carotenoids of biotechnological importance. Adv. Biochem. Eng. Biotechnol. 2015, 148, 449–467.
- Gupta, S.K.; Trivedi, D.; Srivastava, S.; Joshi, S.; Halder, N.; Verma, S.D. Lycopene attenuates oxidative stress induced experimental cataract development: An in vitro and in vivo study. Nutrition 2003, 19, 794–799.
- SanGiovanni, J.P.; Neuringer, M. The putative role of lutein and zeaxanthin as protective agents against age-related macular degeneration: Promise of molecular genetics for guiding mechanistic and translational research in the field. Am. J. Clin. Nutr. 2012, 96, 1223S–1233S.
- Eggersdorfer, M.; Wyss, A. Carotenoids in human nutrition and health. Arch. Biochem. Biophys. 2018, 652, 18–26.
- Ungureanu, C.; Ferdes, M. Evaluation of antioxidant and antimicrobial activities of torularhodin. Adv. Sci. Lett. 2012, 5, 1–4.
- Keceli, T.M.; Erginkaya, Z.; Turkkan, E.; Kaya, U. Antioxidant and antibacterial effects of carotenoids extracted from Rhodotorula glutinis strains. Asian J. Chem. 2013, 25, 42–46.
- Kot, A.M.; Błażejak, S.; Kurcz, A.; Gientka, I.; Kieliszek, M. Rhodotorula glutinis—Potential source of lipids, carotenoids, and enzymes for use in industries. Appl. Microbiol. Biotechnol. 2016, 100, 6103–6117.
- Kot, A.M.; Błażejak, S.; Gientka, I.; Kieliszek, M.; Brys, J. Torulene and torularhodin: “new” fungal carotenoids for industry? Microb. Cell Fact. 2018, 17, 49.
- Ungureanu, C.; Popescu, S.; Purcel, G.; Tofan, V.; Popescu, M.; Sălăgeanu, A.; Pîrvu, C. Improved antibacterial behavior of titanium surface with torularhodin-polypyrrole film. Mater. Sci. Eng. C 2014, 42, 726–733.
- Ungureanu, C.; Dumitriu, C.; Popescu, S.; Enculescu, M.; Tofan, V.; Popescu, M.; Pirvu, C. Enhancing antimicrobial activity of TiO2/Ti by torularhodin bioinspired surface modification. Bioelectrochemistry 2016, 107, 14–24.
- Haddad, N.; Levy, R. The synergistic anti-inflammatory effects of lycopene, lutein, β-carotene, and carnosic acid combinations via redox-based inhibition of NF-kB signaling. Free Radic. Biol. Medic. 2012, 53, 1381–1391.
- Katan, M.B.; Grundy, S.M.; Jones, P.; Law, M.; Miettinen, T.; Paoletti, R. Efficacy and safety of plant stanols and sterols in the management of blood cholesterol levels. Mayo Clin. Proc. 2003, 78, 965–968.
- Rao, A.V. Lycopene, tomatoes, and the prevention of coronary heart disease. Exp. Biol. Med. 2002, 227, 908–913.
- Rao, A.V.; Rao, L.G. Carotenoids and human health. Pharmacol. Res. 2007, 55, 207–216.
- McEneny, J.; Wade, L.; Young, I.S.; Masson, L.; Duthie, G.; McGinty, A.; McMaster, C.; Thies, F. Lycopene intervention reduces inflammation and improves HDL functionality in moderately overweight middle-aged individuals. J. Nutr. Biochem. 2013, 24, 163–168.
- Kim, J.Y.; Paik, J.K.; Kim, O.Y.; Park, H.V.; Lee, J.H.; Jang, Y.; Lee, J.H. Effects of lycopene supplementation on oxidative stress and markers of endothelial function in healthy men. Atherosclerosis 2011, 215, 189–195.
- Ramesh, C.; Vinithkumar, N.V.; Kirubagaran, R.; Venil, C.K.; Dufossé, L. Multifaceted applications of microbial pigments: Current knowledge, challenges and future directions for public health implications. Microorganisms 2019, 7, 186.
- Gouranton, E.; Thabuis, C.; Riollet, C.; Malezet-Desmoulins, C.; El Yazidi, C.; Amiot, M.J.; Borel, P.; Landrier, J.F. Lycopene inhibits proinflammatory cytokine and chemokine expression in adipose tissue. J. Nutr. Biochem. 2011, 22, 642–648.
- Palozza, P.; Simone, R.; Catalano, A.; Monego, G.; Barini, A.; Mele, M.C.; Parrone, N.; Trombino, S.; Picci, N.; Ranelletti, F.O. Lycopene prevention of oxysterol-induced proinflammatory cytokine cascade in humn macrophages: Inhibition of NF-κB nuclear binding and increase in PPARγ expression. J. Nutr. Biochem. 2011, 22, 259–268.
- Rao, L.G.; Mackinnon, E.S.; Josse, R.G.; Murray, T.M.; Strauss, A.; Rao, A.V. Lycopene consumption decreases oxidative stress and bone resorption markers in postmenopausal women. Osteoporosis Int. 2007, 18, 109–115.
- Viuda-Martos, M.; Sanchez-Zapata, E.; Sayas-Barberá, E.; Sendra, J.; Perez-Alvares, A.; Fernandes-Lopez, J. Tomato and tomato byproducts. Human health benefits of lycopene and its application to meat products: A review. Crit. Rev. Food Sci. Nutr. 2014, 54, 1032–1049.
- Thies, F.; Mills, L.M.; Moir, S.; Masson, L.F. Cardiovascular benefits of lycopene: Fantasy or reality? Proc. Nutr. Soc. 2017, 76, 122–129.
- Walallawita, U.S.; Wolber, F.M.; Ziv-Gal, A.; Kruger, M.C.; Heyes, J.A. Potential role of lycopene in the prevention of postmenopausal bone loss: Evidence from molecular to clinical studies. Int. J. Mol. Sci. 2020, 21, 7119.
- Rao, A.V.; Ray, M.R.; Rao, L.G. Lycopene. Adv. Food Nutr. Res. 2006, 51, 99–164.
- Bonet, M.L.; Ribot, J.; Galmés, S.; Francisca Serra, F.; Palou, A. Carotenoids and carotenoid conversion products in adipose tissue biology and obesity: Pre-clinical and human studies. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2020, 1865, 158676.
- Rao, A.V.; Balachandran, B. Role of oxidative stress and antioxidants in neurodegenerative diseases. Nutr. Neurosci. 2003, 5, 291–309.
- Wu, W.; Li, Y.; Wu, Y.; Zhang, Y.; Wang, Z.; Liu, X. Lutein suppresses inflammatory responses through Nrf2 activation and NF-κB inactivation in lipopolysaccharide-stimulated BV-2 microglia. Mol. Nutr. Food Res. 2015, 59, 1663–1673.
- Wu, J.L.; Wang, W.Y.; Cheng, Y.L.; Du, C.; Qian, H. Neuroprotective effects of torularhodin against H2O2-induced oxidative injury and apoptosis in PC12 cells. Pharmazie 2015, 70, 17–23.
- Longnecker, M.P.; Kamel, F.; Umbach, D.M.; Munsat, T.L.; Shefner, J.M.; Lansdell, L.W.; Sandler, D.P. Dietary intake of calcium, magnesium and antioxidants in relation to risk of amyotrophic lateral sclerosis. Neuroepidemiology 2000, 19, 210–216.
- Mikhailova, E.V.; Artemov, A.V.; Snigirevskaya, E.S.; Artamonova, T.O.; Khodorkovskii, M.A.; Soidla, T.R.; Nevzglyadova, O.V. Effect of red pigment on insulin fibril formation in vitro. Cell Tiss. Biol. 2011, 5, 580–585.
- Nevzglyadova, O.V.; Artemov, A.V.; Mittenberg, A.G.; Mikhailova, E.V.; Kuznetsova, I.M.; Turoverov, K.K.; Soidla, T.R. Effect of red pigment on amyloidization of yeast. Cell Tiss. Biol. 2010, 4, 152–166.
- Nevzglyadova, O.V.; Kuznetsova, I.M.; Mikhailova, E.V.; Artamonova, T.O.; Artemov, A.V.; Mittenberg, A.G.; Kostyleva, E.I.; Turoverov, K.K.; Khodorkovskii, M.A.; Soidla, T.R. The effect of red pigment on amyloidization of yeast proteins. Yeast 2011, 28, 505–526.
- Burton, G.W.; Ingold, K.U. Beta-carotene: An unusual type of lipid antioxidant. Science 1984, 224, 569–573.
- Krinsky, N.I. Antioxidant functions of carotenoids. Free Radic. Biol. Med. 1989, 7, 617–635.
- Kiokias, S.; Gordon, M.H. Dietary supplementation with a natural carotenoid mixture decreases oxidative stress. Eur. J. Clin. Nutr. 2003, 57, 1135–1140.
- Krinsky, N.I.; Johnson, E.J. Carotenoid actions and their relation to health and disease. Mol. Asp. Med. 2005, 26, 459–516.
- Sarada, S.; Dipti, P.; Anju, B.; Pauline, T.; Kain, A.; Sairam, M.; Sharma, S.; Ilavazhagan, G.; Kumar, D.; Selvamurthy, W. Antioxidant effect of β-carotene on hypoxia induced oxidative stress in male albino rats. J. Ethnopharmacol. 2002, 79, 149–153.
- Amarowicz, R. Lycopene as a natural antioxidant. Eur. J. Lipid Sci. Technol. 2011, 113, 675–677.
- Sies, H.; Stahl, W. Nutritional protection against skin damage from sunlight. Annu. Rev. Nutr. 2004, 24, 173–200.
- Berneburg, M.; Krutmann, J. Photoimmunology, DNA repair and photocarcinogenesis. J. Photochem. Photobiol. B 2000, 54, 87–93.
- Krutmann, J. Ultraviolet A radiation-induced biological effects in human skin: Relevance for photoaging and photodermatosis. J. Dermatol. Sci. 2000, 23, S22–S26.
- Nishino, H.; Tokuda, H.; Murakoshi, M.; Satomi, Y.; Masuda, M.; Onozuka, M.; Yamaguchi, S.; Takayasu, J.; Tsuruta, J.; Okuda, M. Cancer prevention by natural carotenoids. Biofactors 2000, 13, 89–94.
- Ajila, C.; Brar, S. Role of dietary antioxidants in cancer. In Nutrition, Diet and Cancer; Shankar, S., Srivastava, R.K., Eds.; Springer: Dordrecht, Germany, 2012; pp. 377–412.
- Tanaka, T.; Shnimizu, M.; Moriwaki, H. Cancer chemoprevention by carotenoids. Molecules 2012, 17, 3202–3242.
- Haddad, N.F.; Teodoro, A.J.; Oliveira, F.L.; Soares, N.; de Mattos, R.M.; Hecht, F.; Dezonne, R.S.; Vairo, L.; Goldenberg, R.C.; Gomes, F.C.A.; et al. Lycopene and β-carotene induce growth inhibition and proapoptotic effects on ACTH-secreting pituitary adenoma cells. PLoS ONE 2013, 8, e62773.
- Holzapfel, N.P.; Holzapfel, B.M.; Champ, S.; Feldthusen, J.; Clements, J.; Hutmacher, D.W. The potential role of lycopene for the prevention and therapy of prostate cancer: From molecular mechanisms to clinical evidence. Int. J. Mol. Sci. 2013, 14, 14620–14646.
- Rokkaku, T.; Kimura, R.; Ishikawa, C.; Yasumoto, T.; Senba, M.; Kanaya, F.; Mori, N. Anticancer effects of marine carotenoids, fucoxanthin and its deacetylated product, fucoxanthinol, on osteosarcoma. Int. J. Oncol. 2013, 43, 1176–1186.
- Gloria, N.F.; Soares, N.; Brand, C.; Oliveira, F.L.; Borojevic, R.; Teodoro, A.J. Lycopene and beta-carotene induce cell cycle arrest and apoptosis in human breast cancer cell lines. Anticancer Res. 2014, 34, 1377–1386.
- Sugawara, T.; Ganesan, P.; Li, Z.; Manabe, Y.; Hirata, T. Siphonaxanthin, a green algal carotenoid, as a novel functional compound. Marine Drugs 2014, 12, 3660–3668.
- Zu, K.; Mucci, L.; Rosner, B.A.; Clinton, S.K.; Loda, M.; Stampfer, M.J.; Giovannucci, E. Dietary lycopene, angiogenesis, and prostate cancer: A prospective study in the prostate-specific antigen era. J. Natl. Cancer Inst. 2014, 106, djt430.
- Du, C.; Li, Y.; Guo, Y.; Han, M.; Zhang, W.; Qian, H. The suppression of torulene and torularhodin treatment on the growth of PC-3 xenograft prostate tumors. Biochem. Biophys. Res. Commun. 2016, 469, 1146–1152.
- Linnewiel-Hermoni, K.; Khanin, M.; Danilenko, M.; Zango, G.; Amosi, Y.; Levy, J.; Sharoni, Y. The anti-cancer effects of carotenoids and other phytonutrients resides in their combined activity. Arch. Biochem. Biophys. 2015, 572, 28–35.
- Prakash, P.; Russell, R.M.; Krinsky, N.I. In vitro inhibition of proliferation of estrogen-dependent and estrogen-independent human breast cancer cells treated with carotenoids or retinoids. J. Nutr. 2001, 131, 1574–1580.
- Block, G.; Patterson, B.; Subar, A. Fruit, vegetables, and cancer prevention: A review of the epidemiological evidence. Nutr. Cancer 1992, 18, 1–29.
- Ziegler, R.G.; Mayne, S.T.; Swanson, C.A. Nutrition and lung cancer. Cancer Causes Control 1996, 7, 157–177.
- Omen, G.S.; Goodman, G.E.; Thornquist, M.D.; Balmes, J.; Cullen, M.R.; Glass, A.; Keogh, J.P.; Meyskens, F.L.; Valanis, B.; Williams, J.H.; et al. Effects of a combination of beta-carotene and vitamin A on lung cancer and cardiovascular disease. N. Engl. J. Med. 1996, 334, 1150–1155.
- Rowles, J.L., III; Erdman, J.W., Jr. Carotenoids and their role in cancer prevention. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2020, 1865, 158613.
- Lim, J.Y.; Wang, X.-D. Mechanistic understanding of β-cryptoxanthin and lycopene in cancer prevention in animal models. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2020, 1865, 158652.
- Bendich, A. Carotenoids and immune response. J. Nutr. 1989, 119, 112–115.
- Krinsky, N.I. Actions of carotenoids in biological systems. Annu. Rev. Nutr. 1993, 13, 561–587.
- Shultz, T.D.; Chew, B.P.; Seaman, W.R.; Luedecke, L.O. Inhibitory effects of conjugated dienoic derivatives of linoleic acid and beta-carotene on /the in vitro growth of human cancer cells. Cancer Lett. 1992, 63, 125–133.
- Santamaria, L.; Bianchi, A.; Andreoni, L.; Santagati, G.; Arnaboldi, A.; Bermond, P. 8-Methoxypsoralen photocarcinogenesis and its prevention by dietary carotenoids. Preliminary results. Med. Biol. Environ. 1984, 12, 533–537.
- Belisario, M.A.; Pecce, R.; Battista, C.; Panza, N.; Pacilio, G. Inhibition of cyclophosphamide mutagenicity by β-carotene. Biomed. Pharmacother. 1985, 39, 445–448.
- Azuine, M.A.; Goswami, U.C.; Kayal, J.J.; Bhide, S.V. Antimutagenic and anticarcinogenic effects of carotenoids and dietary palm oil. Nutr. Cancer 1992, 17, 287–295.
- Arriaga-Alba, M.; Rivera-Sanchez, R.; Parra-Cervantes, G.; Barro-Moreno, F.; Flores-Paz, R.; Garcıa-Jimenez, E. Antimutagenesis of β-carotene to mutations induced by quinolone on Salmonella typhimurium. Arch. Med. Res. 2000, 31, 156–161.
- Pung, A.O.; Rundhaug, J.E.; Yoshizawa, C.N.; Bertram, J.S. β-Carotene and canthaxanthin inhibit chemically- and physically-induced neoplastic transformation in 10T1/2 cells. Carcinogenesis 1988, 9, 1533–1539.
- Bertram, J.S.; Pung, A.; Churley, M.; Kappock, T.J., IV; Wilkins, L.R.; Cooney, R.V. Diverse carotenoids protect against chemically induced neoplastic transformation. Carcinogenesis 1991, 12, 671–678.
- Saini, R.K.; Keum, Y.-S. Microbial platforms to produce commercially vital carotenoids at industrial scale: An updated review of critical issues. J. Ind. Microbiol. Biotechnol. 2019, 46, 657–674.
- Tyczkowski, J.K.; Hamilton, P.B. Absorption, transport, and deposition in chickens of lutein diester, a carotenoid extracted from Marigold (Tagetes erecta) petals. Poult. Sci. 1986, 65, 1526–1531.
- Mortensen, A. Carotenoids and other pigments as natural colorants. Pure Appl. Chem. 2006, 78, 1477–1491.
- Mannazzu, I.; Landolfo, S.; Lopes da Silva, T.; Buzzini, P. Red yeasts and carotenoid production: Outlining a future for non-conventional yeasts of biotechnological interest. World J. Microbiol. Biotechnol. 2015, 31, 1665–1673.
- Nabi, F.; Arain, M.A.; Rajput, N.; Alagawany, M.; Soomro, J.; Umer, M.; Soomro, F.; Wang, Z.; Ye, R.; Liu, J. Health benefits of carotenoids and potential application in poultry industry: A review. J. Anim. Physiol. Anim. Nutrit. 2020, 104, 104–1809.
- Johnson, E.A.; An, G.H. Astaxanthin from microbial sources. Crit. Rev. Biotechnol. 1991, 11, 297–326.
- Johnson, E.A. Phaffia rhodozyma: Colorfull odyssey. Int. Microbiol. 2003, 6, 169–174.
- Marcoleta, A.; Niklitschek, M.; Wozniak, A.; Lozano, C.; Alcaıno, J.; Baeza, M.; Cifuentes, V. Glucose and ethanol-dependent transcriptional regulation of the astaxanthin biosynthesis pathway in Xanthophyllomyces dendrorhous. BMC Microbiol. 2011, 11, 190.
- Martinez-Moya, P.; Niehaus, K.; Alcaıno, J.; Baeza, M.; Cifuentes, V. Proteomic and metabolomic analysis of the carotenogenic yeast Xanthophyllomyces dendrorhous using different carbon sources. BMC Genomics 2015, 16, 289.
- Aruldass, C.A.; Dufosse, L.; Ahmad, W.A. Current perspective of yellowish-orange pigments from microorganisms—A review. J. Clean. Prod. 2018, 168–182.
- Tuli, H.S.; Chaudhary, P.; Beniwal, V.; Sharma, A.K. Microbial pigments as natural color sources: Current trends and future perspectives. J. Food Sci. Technol. 2014, 52, 4669–4678.
- Lagashetti, A.C.; Dufossé, L.; Singh, S.K.; Singh, P.N. Fungal pigments and their prospects in different industries. Microorganisms 2019, 7, 604.
- Venil, C.K.; Velmurugan, P.; Dufossé, L.; Devi, P.R.; Ravi, A.V. Fungal pigments: Potential coloring compounds for wide ranging applications in textile dyeing. J. Fungi 2020, 6, 68.
- Tramontin, L.R.R.; Kildegaard, K.R.; Sudarsan, S.; Borodina, I. Enhancement of astaxanthin biosynthesis in oleaginous yeast Yarrowia lipolytica via microalgal pathway. Microorganisms 2019, 7, 472.
- Dufosse, L. Microbial production of food grade pigments. Food Technol. Biotechnol. 2006, 44, 313–321.
- Dufosse, L.; Fouillaud, M.; Caro, Y.; Mapari, S.A.S.; Sutthiwong, N. Filamentous fungi are large-scale producers of pigments and colorants for the food industry. Curr. Opin. Biotechnol. 2014, 26, 56–61.
- Mapari, S.A.S.; Hansen, M.E.; Meyer, A.S.; Thrane, U. Computerized screening for novel producers of Monascus-like food pigments in Penicillium species. J. Agric. Food Chem. 2008, 56, 9981–9989.
- Mapari, S.A.S.; Meyer, A.S.; Thrane, U.; Frisvad, J.C. Identification of potentially safe promising fungal cell factories for the production of polyketide natural food colorants using chemotaxonomic rationale. Microb. Cell Fact. 2009, 8, 24.
- Dufossé, L. Red colourants from filamentous fungi: Are they ready for the food industry? J. Food Compos. Anal. 2018, 69, 156–161.
- Morales-Oyervides, L.; Ruiz-Sánchez, J.P.; Oliveira, J.C.; Sousa-Gallagher, M.J.; Mendez-Zavala, A.; Giuffrida, D.; Dufossé, L.; Montañez, J. Biotechnological approaches for the production of natural colorants by Talaromyces/Penicillium: A review. Biotechnol. Adv. 2020, 43, 107601.