Fusion RNAs are a hallmark of some cancers. They result either from chromosomal rearrangements or from splicing mechanisms that are non-chromosomal rearrangements.
- fusion RNAs
- gene fusions
- chromosomal rearrangements
- trans-splicing
1. Introduction
Cancer is a disease of the genome [1][2]. Gene fusions or chromosomal rearrangements are an important class of somatic alterations in cancer and can have important roles in the initial steps of tumorigenesis. [3][4][5]. The first cancer-associated chromosomal rearrangement was identified in 1960 as a translocation of chromosomes 9 and 22 [4][6][7]. The abnormally small resulting chromosome, named the Philadelphia chromosome, was found in over 95% of patients with chronic myelogenous leukemia (CML) and consisted of the breakpoint cluster region (
Cancer is a disease of the genome [1,2]. Gene fusions or chromosomal rearrangements are an important class of somatic alterations in cancer and can have important roles in the initial steps of tumorigenesis. [3,4,5]. The first cancer-associated chromosomal rearrangement was identified in 1960 as a translocation of chromosomes 9 and 22 [4,6,7]. The abnormally small resulting chromosome, named the Philadelphia chromosome, was found in over 95% of patients with chronic myelogenous leukemia (CML) and consisted of the breakpoint cluster region (
BCR
) gene fused to the second exon of the Abelson murine leukemia viral oncogene homolog 1 (
ABL1) gene [8][9]. Additional examples of cancer-associated chromosomal aberrations have been identified in other hematological malignancies and sarcomas; for example, mixed lineage leukemia (MLL) fusions,
) gene [8,9]. Additional examples of cancer-associated chromosomal aberrations have been identified in other hematological malignancies and sarcomas; for example, mixed lineage leukemia (MLL) fusions,
RUNX1–RUNX1T1
and
PML
–
RARα
,
EWSR1
–
FLI1
and
EVT6
–
NTRK3 [10][11]. Although originally discovered in hematological malignancies, gene fusions are now known to occur in several solid tumor types [7][12][13]. The first fusion gene found in a solid tumor was
[10,11]. Although originally discovered in hematological malignancies, gene fusions are now known to occur in several solid tumor types [7,12,13]. The first fusion gene found in a solid tumor was
CTNNB1
–
PLAG1 in salivary gland adenoma, which is usually benign [12][14][15]. Other fusion genes were soon discovered in solid tumors and other malignancies, including glioblastoma, melanoma, and prostate, breast, ovarian, lung, colorectal, and head and neck cancers [13].
in salivary gland adenoma, which is usually benign [12,14,15]. Other fusion genes were soon discovered in solid tumors and other malignancies, including glioblastoma, melanoma, and prostate, breast, ovarian, lung, colorectal, and head and neck cancers [13].
Most of the nonprotein-coding region of the human genome was previously considered to be “junk DNA” [16]. With the advent of massive parallel sequencing technology, these regions in the human genome have been clearly shown to transcribe dynamically and differentially into noncoding RNAs (ncRNAs), such as microRNAs (miRNAs), small nucleolar RNAs (snoRNAs), long ncRNAs (lncRNAs), and circular RNAs (circRNAs) [17][18][19][20]. Accumulating evidence indicates that lncRNAs play critical roles in diverse biological processes, including differentiation, stem cell pluripotency, embryogenesis, pathogenic infection, neurogenesis, proliferation, and tumorigenesis [16][20][21][22][23][24][25][26]. LncRNAs also function in chromatin and genomic structural remodeling, RNA trafficking, RNA stabilization, transcriptional regulation, translation, signal pathway, and protein degradation [27][28][29][30][31]. Expression of lncRNA, miRNA, and snoRNA have shown close correlations with specific chromosomal rearrangements in cancers [10][32][33][34]. Moreover, fusion circRNAs (f-circRNAs) that are generated by chromosome rearrangement contribute to oncogenic roles [35]. Furthermore, the fusion RNA
Most of the nonprotein-coding region of the human genome was previously considered to be “junk DNA” [16]. With the advent of massive parallel sequencing technology, these regions in the human genome have been clearly shown to transcribe dynamically and differentially into noncoding RNAs (ncRNAs), such as microRNAs (miRNAs), small nucleolar RNAs (snoRNAs), long ncRNAs (lncRNAs), and circular RNAs (circRNAs) [17,18,19,20]. Accumulating evidence indicates that lncRNAs play critical roles in diverse biological processes, including differentiation, stem cell pluripotency, embryogenesis, pathogenic infection, neurogenesis, proliferation, and tumorigenesis [16,20,21,22,23,24,25,26]. LncRNAs also function in chromatin and genomic structural remodeling, RNA trafficking, RNA stabilization, transcriptional regulation, translation, signal pathway, and protein degradation [27,28,29,30,31]. Expression of lncRNA, miRNA, and snoRNA have shown close correlations with specific chromosomal rearrangements in cancers [10,32,33,34]. Moreover, fusion circRNAs (f-circRNAs) that are generated by chromosome rearrangement contribute to oncogenic roles [35]. Furthermore, the fusion RNA
SLC45A3
–
ELK4
, which regulates cancer cell proliferation, functions as a lncRNA [36]. However, details of mechanisms of the oncogenic roles of these fusion RNAs are unclear.
The ultimate goal of precision medicine in cancer treatment is the development of therapeutic strategies that specifically target cancer cells without affecting normal cells [37]. Targeting oncogenic fusion genes and RNAs specific to cancer tissue for treatment and diagnosis could bring us closer to the approach. Moreover, these fusions are often present at clonal levels within tumors; their generation is frequently the founding genetic abnormality that drives the cancer [37][38].
The ultimate goal of precision medicine in cancer treatment is the development of therapeutic strategies that specifically target cancer cells without affecting normal cells [37]. Targeting oncogenic fusion genes and RNAs specific to cancer tissue for treatment and diagnosis could bring us closer to the approach. Moreover, these fusions are often present at clonal levels within tumors; their generation is frequently the founding genetic abnormality that drives the cancer [37,38].
2. Biological Functions of Fusion Genes and RNAs
Solid tumors and hematopoietic malignancies often have highly complex, unstable genomes. Many gene fusions are random events caused by genetic instability or abnormal splicing machinery [39][40]. These changes at the gene or RNA level are unlikely to result in functional nucleic acids or proteins, as they may occur in regions where there are no known genes [12]. The functions of fusion genes and RNAs are diverse and dependent on the location of the fusion junction. However, the presence of a genomic fusion in a tumor does not necessarily mean that the fusion affects cancer development or tumorigenesis. Fusion RNAs in which the fusion junction is within the protein-coding region may be largely out-of-frame, and such out-of-frame fusions are unlikely to be functional. However, out-of-frame fusion RNAs may function as long noncoding RNAs and may exhibit regulatory functions [41].
Solid tumors and hematopoietic malignancies often have highly complex, unstable genomes. Many gene fusions are random events caused by genetic instability or abnormal splicing machinery [39,40]. These changes at the gene or RNA level are unlikely to result in functional nucleic acids or proteins, as they may occur in regions where there are no known genes [12]. The functions of fusion genes and RNAs are diverse and dependent on the location of the fusion junction. However, the presence of a genomic fusion in a tumor does not necessarily mean that the fusion affects cancer development or tumorigenesis. Fusion RNAs in which the fusion junction is within the protein-coding region may be largely out-of-frame, and such out-of-frame fusions are unlikely to be functional. However, out-of-frame fusion RNAs may function as long noncoding RNAs and may exhibit regulatory functions [41].
An RNA fused between two genes in-frame is translated into a novel fusion protein that may act as a potent oncogenic driver. Kinase genes are often partners in such fusions [7][12]. Kinase fusions often retain kinase activity and result in ligand-independent constitutive activation and enhanced downstream signaling that leads to carcinogenesis [7][12]. Tyrosine kinase fusions that contain kinase-encoding genes, such as
An RNA fused between two genes in-frame is translated into a novel fusion protein that may act as a potent oncogenic driver. Kinase genes are often partners in such fusions [7,12]. Kinase fusions often retain kinase activity and result in ligand-independent constitutive activation and enhanced downstream signaling that leads to carcinogenesis [7,12]. Tyrosine kinase fusions that contain kinase-encoding genes, such as
ALK
,
ROS1
,
RET
,
FGFR1/2/3
and
NTRK1, have been detected in various types of cancer, including glioblastoma, melanoma, and carcinomas of head and neck, breast, lung, prostate, bladder, and thyroid gland [12][40][42][43]. Serine–threonine kinase fusions have also been reported [12][40][43]. These kinase fusions frequently cause activation of signaling pathways that play important roles in cell growth, survival, proliferation, and apoptosis [12][40][42][43]. In addition, kinases are ideal targets for cancer therapy; several inhibitors against kinases, such as ALK and BRAF, have been used to treat cancers with fused genes [12][44].
, have been detected in various types of cancer, including glioblastoma, melanoma, and carcinomas of head and neck, breast, lung, prostate, bladder, and thyroid gland [12,40,42,43]. Serine–threonine kinase fusions have also been reported [12,40,43]. These kinase fusions frequently cause activation of signaling pathways that play important roles in cell growth, survival, proliferation, and apoptosis [12,40,42,43]. In addition, kinases are ideal targets for cancer therapy; several inhibitors against kinases, such as ALK and BRAF, have been used to treat cancers with fused genes [12,44].
Fusion of transcription factors usually produces a fusion protein that leads to constitutive activation or an altered target gene, providing aberrant transcriptional machinery and cell transformation [12]. For example, the TMPRSS2–ERG fusion protein, the most common fusion in prostate cancers, mediates overexpression of E26 transformation-specific (ETS) family transcription factors in response to androgen and thus aberrantly activates downstream oncogenes that play important roles in many biological processes, including cell proliferation, angiogenesis, and invasiveness [45][46]. Moreover, the EWSR1–FLI1 fusion protein can gain the ability to bind to the genome and change the transcriptional mechanism [47][48]. Transcription factor fusions can induce a wide range of phenotypic changes that initiate or promote tumorigenesis. However, they have been generally more difficult to work with as therapeutic targets than kinase fusions [37].
Fusion of transcription factors usually produces a fusion protein that leads to constitutive activation or an altered target gene, providing aberrant transcriptional machinery and cell transformation [12]. For example, the TMPRSS2–ERG fusion protein, the most common fusion in prostate cancers, mediates overexpression of E26 transformation-specific (ETS) family transcription factors in response to androgen and thus aberrantly activates downstream oncogenes that play important roles in many biological processes, including cell proliferation, angiogenesis, and invasiveness [45,46]. Moreover, the EWSR1–FLI1 fusion protein can gain the ability to bind to the genome and change the transcriptional mechanism [47,48]. Transcription factor fusions can induce a wide range of phenotypic changes that initiate or promote tumorigenesis. However, they have been generally more difficult to work with as therapeutic targets than kinase fusions [37].
3. Biosynthesis Patterns of Fusion Genes and RNAs
Fusion RNAs are known to be generated by three mechanisms [7][49][50]. The best-understood of these is chromosomal rearrangement. Two other mechanisms are grouped together as “splicing”. One is trans-splicing, in which exons from two separate RNA transcripts are spliced together. The other is cis-splicing, which involves adjacent genes on the same strand.
Fusion RNAs are known to be generated by three mechanisms [7,49,50]. The best-understood of these is chromosomal rearrangement. Two other mechanisms are grouped together as “splicing”. One is trans-splicing, in which exons from two separate RNA transcripts are spliced together. The other is cis-splicing, which involves adjacent genes on the same strand.
RNA splicing is a form of RNA processing in which a newly made precursor messenger RNA (pre-mRNA) is transformed into a mature messenger RNA (mRNA) [51][52]. It has important functions in regulating the RNA and protein diversity observed in organisms [51][52]. Pre-mRNA splicing involves recognizing and removing noncoding regions (intron excision) and the concomitant joining of coding regions (exon ligation) to produce mature mRNA. For many eukaryotic introns, splicing is performed in a series of reactions that are catalyzed by the spliceosome, a complex of small nuclear ribonucleoproteins that incorporates stepwise assembly and disassembly by several hundred proteins and five small nuclear RNAs [53][54][55]. Mechanisms for trans- and cis-splicing between neighboring genes are not well understood. In this section, we review the mechanism of fusion genes and RNA generation that are currently known.
RNA splicing is a form of RNA processing in which a newly made precursor messenger RNA (pre-mRNA) is transformed into a mature messenger RNA (mRNA) [51,52]. It has important functions in regulating the RNA and protein diversity observed in organisms [51,52]. Pre-mRNA splicing involves recognizing and removing noncoding regions (intron excision) and the concomitant joining of coding regions (exon ligation) to produce mature mRNA. For many eukaryotic introns, splicing is performed in a series of reactions that are catalyzed by the spliceosome, a complex of small nuclear ribonucleoproteins that incorporates stepwise assembly and disassembly by several hundred proteins and five small nuclear RNAs [53,54,55]. Mechanisms for trans- and cis-splicing between neighboring genes are not well understood. In this section, we review the mechanism of fusion genes and RNA generation that are currently known.
3.1. Chromosomal Rearrangement
Gene fusions are usually caused by alterations in genomic structure resulting from DNA damage and by subsequent erroneous recombination and replication [7]. Genomic rearrangements can occur between one or two independent genes through six different known mechanisms: translocation, insertion, inversion, tandem duplication, deletion, and chromothripsis (
Figure 1) [7][50][56]. Inversions, tandem duplications, and chromosomal deletions can occur within one or two adjacent genes, whereas translocations and insertions represent large-scale genomic aberrations that result from interactions between distant regions of the genome (interchromosomal rearrangements) or within the same chromosome (intrachromosomal rearrangements) [7][50][56]. As a result, gene fusions can produce aberrant fusion RNAs and proteins that may activate, reduce, or eliminate their original functions.
) [7,50,56]. Inversions, tandem duplications, and chromosomal deletions can occur within one or two adjacent genes, whereas translocations and insertions represent large-scale genomic aberrations that result from interactions between distant regions of the genome (interchromosomal rearrangements) or within the same chromosome (intrachromosomal rearrangements) [7,50,56]. As a result, gene fusions can produce aberrant fusion RNAs and proteins that may activate, reduce, or eliminate their original functions.
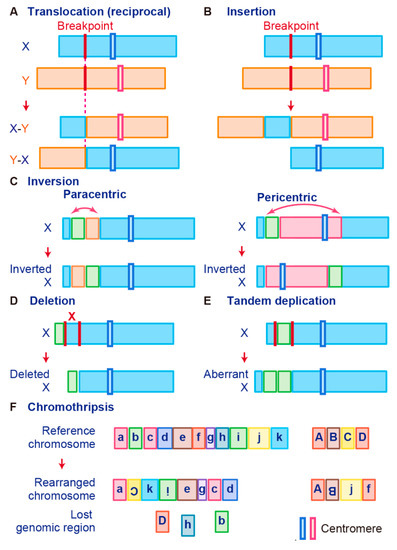
Figure 1.
Schematic representation of fusion gene formation by structural chromosome rearrangements. (
A
) Translocation. (
B
) Insertion. (
C
) Inversion. (
D
) Deletion. (
E
) Tandem duplication. (
F
) Chromothripsis.
A chromosomal translocation is an exchange of parts between two nonhomologous chromosomes, also called a reciprocal translocation. Chromosomal translocation can occur anywhere between any two chromosomes (
A).
The second type of translocation is an insertion. Insertions are caused by transfer of DNA fragments from one region to another within the same chromosome (intrachromosome) or from one chromosome to another (interchromosome). The latter is also known as a nonreciprocal nonmutual translocation (
B). The
BCR–ABL1
, the first oncogenic fusion gene ever identified, is formed by a reciprocal chromosomal translocation.
BCR–ABL1
, which is generated from translocation t(9; 22) (q34; q11), is characteristic of CML [9] and also found in acute lymphoid leukemia (ALL) [57] and acute myelogenous leukemia (AML) [58]. The fusion gene
BCR–ABL1
has a constitutive tyrosine kinase activity, which leads to sustained stimulation on proliferation of cancer cells [8]. One chromosomal abnormality discovered after
BCR–ABL1
was a consistent nonhomologous balanced translocation between chromosomes 8 and 21 in leukemia patients [59]. This t(8; 21) translocation is one of the most common genetic defects in AML; it gives rise to the
RUNX1–RUNX1T1
fusion gene (previously called
RUNX1–RUNX1T1 fusion protein interacts with other proteins to repress transcription and induce leukemogenesis in myeloid progenitor cells [60][63].
fusion protein interacts with other proteins to repress transcription and induce leukemogenesis in myeloid progenitor cells [60,63].
Promyelocytic leukemia (
PML
)–retinoic acid receptor alpha (
RARα) is a fusion RNA found in almost 95% of acute promyelocytic leukemia (APL). It is generated from the t(15;17) reciprocal translocation in APL [64][65][66][67][68]. PML is a key component of PML bodies and many proteins have been associated with PML in cells [69]. The PML–RARα protein disrupts PML bodies and induces the formation of dispersed microspeckles with the loss of transcriptional activation ability [70][71]. In addition to leukemia, fusion events also occur in Ewing’s sarcoma (EWS). Many EWS cases involve t(11; 22) or t(21; 22) translocation that fuses the 5′ end of the Ewing’s sarcoma breakpoint region-1 (
) is a fusion RNA found in almost 95% of acute promyelocytic leukemia (APL). It is generated from the t(15;17) reciprocal translocation in APL [64,65,66,67,68]. PML is a key component of PML bodies and many proteins have been associated with PML in cells [69]. The PML–RARα protein disrupts PML bodies and induces the formation of dispersed microspeckles with the loss of transcriptional activation ability [70,71]. In addition to leukemia, fusion events also occur in Ewing’s sarcoma (EWS). Many EWS cases involve t(11; 22) or t(21; 22) translocation that fuses the 5′ end of the Ewing’s sarcoma breakpoint region-1 (
EWSR1
) gene to the 3′ end of the
FLI1
or
ERG
gene, which generates the fusion genes
EWS
–
FLI1
or
EWS
–
ERG, respectively [47][48][72]. These fusion transcription factors upregulate genes related to the cell cycle, invasion, and proliferation pathways [73][74][75][76]. Interestingly, the prion-like domain of EWS–FLI1, which is necessary for phase transitions, induces recruitment of BRG1/BRM-associated factors (BAF) complexes to
, respectively [47,48,72]. These fusion transcription factors upregulate genes related to the cell cycle, invasion, and proliferation pathways [73,74,75,76]. Interestingly, the prion-like domain of EWS–FLI1, which is necessary for phase transitions, induces recruitment of BRG1/BRM-associated factors (BAF) complexes to
GGAA
microsatellites that are frequently found in oncogenes and activate the transcription of target genes [77].
A chromosome inversion occurs when a chromosome undergoes a break or rearrangement within a single chromosome (
C). There are two types of inversions: paracentric and pericentric. Paracentric inversions do not involve centromeres and both breaks occur in a single chromosome arm, whereas pericentric inversions include a centromere, with one break in each arm (
C). Many chromosomal rearrangements identified in radiation-induced tumors are known to be paracentric inversions. The most common is the
RET
fusion in papillary thyroid carcinoma, which is present in up to 80% of radiation-related tumors [78]. As another example, in 2007,
EML4
–
ALK
was identified as a novel fusion oncogenic driver of non-small cell lung cancer (NSCLC) [79]. The fusion of
EML4
with
ALK
is caused by an inversion of chromosome 2 (inv2) (p21:p23), by which the kinase domain of the receptor-type tyrosine kinase ALK is placed under the control of the constitutive promoter of
EML4
[80]. This fusion allows cancer transformation by activating downstream reactions in the ALK signaling pathway [80].
Chromosome deletion, which is the fourth type of rearrangement, is an alteration in which a chromosome fragment is lost during DNA replication (
D). This chromosome deletion causes the deletion of intergenic regions between two genes that are side by side, and leads to formation of fusion genes by aligning two genes that are transcribed in the same direction. An example is
TMPRSS2–ERG,
generated in prostate cancer via an intron deletion between
TMPRSS2
and
ERG on chromosome 21q22.2-3 [81][82]. Transmembrane serine protease 2 (TMPRSS2) is a prostate-specific androgen-regulated protein, and ETS-related gene (ERG) belongs to the ETS family of transcription factors, which can be oncogenic [81][82][83].
on chromosome 21q22.2-3 [81,82]. Transmembrane serine protease 2 (TMPRSS2) is a prostate-specific androgen-regulated protein, and ETS-related gene (ERG) belongs to the ETS family of transcription factors, which can be oncogenic [81,82,83].
TMPRSS2
–
ERG fusion is reportedly associated with higher tumor stage, increased risk of disease progression, and bone metastasis [46][84].
fusion is reportedly associated with higher tumor stage, increased risk of disease progression, and bone metastasis [46,84].
In tandem duplication, a genomic region is duplicated and fused with a gene from the original region (
E).
FGFR3–TACC3
in glioblastoma is an example of tandem duplication [85]. A tandem duplication also occurs at 7q34 in pilocytic astrocytoma, resulting in a
KIAA1549–BRAF
fusion gene that exhibits constitutive kinase activity. Moreover, a tandem duplication that leads to
C2orf44
–
ALK
fusion occurs in-frame on chromosome 2 in colorectal cancer, resulting in overexpression of the ALK kinase [86].
Tumor cells are associated with high genomic instability, and fusions can occur as a result of complex processes involving several rare and/or complex genetic rearrangements. Chromothripsis, which is the sixth type of rearrangement, occurs when a single chromosome, chromosome region, or a small number of chromosomes are shattered into many fragments and the fragments reassemble incorrectly (
F). Chromothripsis can produce a large number of fusion genes in a single event [87]. The most typical examples of this characteristic event are
PVT1–MYC
and
PVT1–NDRG1
fusions in medulloblastoma. Chromothripsis in medulloblastoma leads to recurrent translocations that eventually fuse a lncRNA
PVT1
to
MYC
, resulting in a continuous oncogenic effect via MYC amplification [88].
3.2. Trans-Splicing
In trans-splicing, exons from different RNA transcripts are spliced and fused together to produce a mature mRNA (
Figure 2A) [89][90][91]. Trans-splicing produces RNAs with exon repetitions or shuffling, as well as RNAs composed of exons transcribed from opposite strands [92][93][94]. The molecular mechanisms of trans-splicing in vertebrates are largely unexplored; however, several models have been proposed [41]. One model is spliceosome-mediated trans-splicing, which uses a canonical splice site of two different primary RNAs. It shows that the spliceosome mechanism, a fundamental component of splicing, can generate trans-spliced fusion RNAs [95][96][97][98].
A) [89,90,91]. Trans-splicing produces RNAs with exon repetitions or shuffling, as well as RNAs composed of exons transcribed from opposite strands [92,93,94]. The molecular mechanisms of trans-splicing in vertebrates are largely unexplored; however, several models have been proposed [41]. One model is spliceosome-mediated trans-splicing, which uses a canonical splice site of two different primary RNAs. It shows that the spliceosome mechanism, a fundamental component of splicing, can generate trans-spliced fusion RNAs [95,96,97,98].
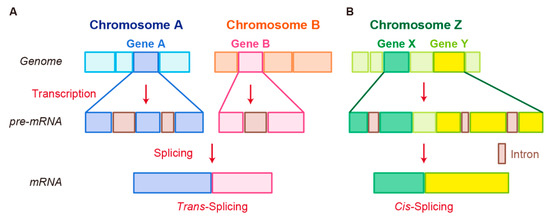
Figure 2.
Schematic representation of fusion RNA formation by nonstructural chromosome rearrangements. (
A
) Trans-splicing. (
B
) Cis-splicing.
Examples of trans-spliced fusion RNAs are JAZF1–JJAZ1 (SUZ12) [89] and PAX3–FOXO1 [99]. In both cases, identical fusions were found as structural chromosomal rearrangements from human tumor tissues and RNA trans-splicing from normal human tissue. JAZF1–SUZ12, which is composed of the first 3 exons of JAZF1 and the last 15 exons of SUZ12, canonically resulted from a recurrent translocation t(7;17)(p15;q21) in endometrial stromal tumors. Identical fusion RNA was detected in normal endometrial cells [89]. Whereas PAX3–FOXO1 with t(2;13) translocation was detected in rhabdomyosarcoma, PAX3–FOXO1 fusion RNA produced by trans-splicing was transiently present in cells that underwent differentiation from pluripotent cells into skeletal muscle [99]. In these cases, different mechanisms at genomic or RNA levels may generate identical fusion RNAs, leading to different pathological outcomes.
3.3. Cis-Splicing
Another splicing mechanism is cis-splicing, in which two neighboring genes are transcribed into a single precursor RNA by transcriptional read-through, followed by RNA splicing between the exons of the two neighboring genes to complete the fusion (
Figure 2B) [7][100]. Although cancer-associated fusion RNAs from cis-splicing are uncommon, some have been reported with clear oncogenic roles.
B) [7,100]. Although cancer-associated fusion RNAs from cis-splicing are uncommon, some have been reported with clear oncogenic roles.
SLC45A3–ELK4 has been discovered by two independent groups and is a potential biomarker in prostate cancer [101][102]. Several
has been discovered by two independent groups and is a potential biomarker in prostate cancer [101,102]. Several
SLC45A3
–
ELK4
fusions have been reported; the primary form is a fusion of
SLC45A3
exon 1 with the last four exons of
ELK4
. Interestingly, although
SLC45A3–ELK4
functions as a fused lncRNA, its knockdown in cancer cells leads to reduction in cell proliferation, despite its unclear oncogenic roles [36].
RBM6–RBM5
is found in several types of cancer; its expression is associated with the size of breast tumors [103].
DUS4L–BCAP29 is found in gastric and prostate cancers and plays a tumor-promoting role in gastric cancer [104][105]. However,
is found in gastric and prostate cancers and plays a tumor-promoting role in gastric cancer [104,105]. However,
DUS4L–BCAP29
is also present in normal tissues and has a growth-promoting effect in normal as well as cancerous tissues [106].