Phosphorous, in the form of phosphate, is a key element in the nutrition of all living beings. In nature, it is present in the form of phosphate salts, organophosphates, and phosphonates.
- phosphate
- transport systems
- PstS
- PitH
1. Introduction
Inorganic phosphate (Pi) is a key nutrient in cell metabolism, since it is constitutive of nucleic acids, phospholipids, teichoic acids, membranes, highly phosphorylated nucleotides, and phosphorylated proteins. It participates in the respiratory chain, and plays an important role in the signalling cascades [1][2][1,2]. Phosphate salts are present in a variety of soil and aquatic environment, but its concentration in different habitats is very diverse. Sometimes phosphate is present in insoluble form (so called rocks phosphate) that is difficult to metabolize and requires its solubilization by microorganisms [3][4][3,4]. In addition, organophosphates are abundant in some particular habitats as products of decay of plant and animal tissues.
Streptomyces are largely soil dwelling organisms, although they are also present in many aquatic environments, including sea waters. Streptomyces species and related actinobacteria are prolific producers of a plethora of secondary metabolites with useful biological activities [5][6][5,6]. The biosynthesis of these metabolites in Streptomyces, filamentous fungi, and plants is controlled by the phosphate concentration in the cells, reflecting the availability of phosphate in the different environments [7]. Phosphate regulates the biosynthesis of secondary metabolites belonging to diverse biosynthetic classes, including polyketides, non-ribosomal peptides, amino acid-derived metabolites, isoprenoids, and complex antibiotics, suggesting that the regulation by phosphate is a widely conserved mechanism that controls: (1) primary metabolism and, therefore, the availability of precursors for the biosynthesis of secondary metabolites, and (2) the expression of the genes encoding these biosynthetic pathways [8].
Phosphate limitation causes a well-known nutritional stress that triggers expression of genes encoding secondary metabolite biosynthetic enzymes [9][10][9,10]. In response to phosphate limitation, Streptomyces species produce extracellular enzymes, including alkaline phosphatases PhoA, PhoC, the Ca2+-dependent phospholipase PhoD [11][12][11,12], and a eukaryotic-type phosphatase, PhoX [13][14][13,14]. A response of Streptomyces and other microorganisms to the phosphate availability in the environment is to maintain a reservoir of phosphate in the form of polyphosphate [15][16][15,16]. This compound is accumulated in large amounts in Streptomyces when grown in phosphate rich media and this allows maintenance of Streptomyces metabolism for a prolonged time in phosphate starvation conditions.
Expression of hundreds of genes is controlled by the availability of phosphate; these genes constitute the Pho regulon [1][17][18][19][1,17,18,19]. Expression of the Pho regulon in Streptomyces is controlled by the two-component system PhoR–PhoP [20][21][20,21]. The mechanism controlling the Pho regulon has been reviewed elsewhere [2][22] [2,22] and is not detailed here.
Since organic and inorganic phosphate are frequently limiting in some habitats, research on the phosphate transport mechanisms is really important. We focus this article on the mechanisms of phosphate transport into the cells in Streptomyces and closely related actinobacteria. Most of the available information has been reported in the model actinomycetes Streptomyces coelicolor or Streptomyces lividans, and in Corynebacterium glutamicum.
2. High Affinity Phosphate Transport Systems
Most bacteria, including Streptomyces and related actinobacteria, contain high affinity and low affinity phosphate transport systems, which are induced by phosphate limitation, although they may respond to different phosphate thresholds for induction. The wide distribution of both the low affinity and high affinity systems in Gram positive and Gram- negative bacteria indicates that there are physiological advantages in having two different transport systems. The low affinity Pit system requires less energy for phosphate transport since it is energized by a proton-symporter system and is used in high phosphate concentration habitats. The high affinity PstSCAB requires more energy for phosphate transport as it is energized by ATP hydrolysis. This high affinity system is used in habitats or environmental conditions in which phosphate is limiting.
Early evidence of the existence of two phosphate transport systems in Streptomyces was provided by the description of two distinct phosphate uptake kinetics in Streptomyces granaticolor [23][28], although the genes encoding these systems were not identified.
2.1. The High Affinity PstSCAB System
pstSCAB
Escherichia coli [24],
S. lividans [20] and
Bacillus subtilis [25]. In these bacteria, the system consists of four proteins PstS, PstC, PstA, and PstB, but in
B. subtilis
Streptomyces
Mycobacterium bovis
S. lividans [26][27][28], whereas in Gram-negative bacteria, it is located in the periplasmic space. PstC and PstA are membrane integral proteins that form a membrane channel and PstB is an ATP hydrolysing protein that energizes the phosphate transport (
coelicolor [29] allowed to identify the
pstSCAB
Streptomyces
pstSCAB cluster responds drastically to the phosphate concentration in the culture medium and is strictly dependent of the activation by the phosphate regulator PhoP [20][21][30]. Deletion of the
pstS
S. lividans
S. coelicolor impaired phosphate transport and delayed sporulation in solid medium [30]. In addition to regulation by the PhoP master regulator, formation of the PstS protein was highly increased in a mutant deficient in polyphosphate kinase [30][31][32]which catalyses, in vitro, the reversible polymerization of the phosphate from ATP into polyphosphate. This is most likely due to the up-regulation of
phoR/phoP
ppk
pstSCAB
S. coelicolor
pstS
S. coelicolor that adjust to the consensus PhoP-binding sequence [21][33]. The
pstSCAB
S. lividans
pstS gene transcript were higher than those of other genes of the operon [28]. Most likely this is due to the presence of transcription termination sites between the
pstS
pstC
S. lividans
pstSCAB operon, but allows a remaining basal level of expression (about 10%) that was controlled by the carbon source [28]. The molecular mechanism of regulation by fructose or other carbon sources has not been fully elucidated. CRP (cAMP receptor protein)-binding sequences have been recently identified in the genome of
Streptomyces roseosporus [34], and a twelve nucleotide repeated sequence upstream of the
pstS gene [28] might correspond to a CRP binding sequence.
2.2. Sensing the Phosphate Limitation: The Seven Proteins Model of Wanner and the Intracellular Teichoic Acid Intermediate Signal Model
pstSCAB
E. coli, models were proposed to try to understand which is the sensing mechanism and, particularly, the transduction cascade that responds to different phosphate concentrations in the periplasmic space. Hsieh and Wanner [35] elaborated a refined model in which they proposed that seven proteins interact at the membrane level, serving to detect the phosphate concentration in the broth and then transmit the signal to control expression of the Pho regulon genes. This model proposes that the four proteins, PstSCAB, of the ABC transporter system interact with PhoR–PhoB and also with the phosphate modulator PhoU (
Streptomyces
phoU
phoR
phoP
pstSCAB
E. coli
phoR
phoP cluster [15][36]. Signalling of the phosphate concentration and its transport by the PstSCAB cassette in
Streptomyces
B. subtilis, the signal is an intermediate in the teichoic acid biosynthesis that inhibits the PhoR autokinase activity and, therefore, decreases the phosphorylation of PhoP and the subsequent activation of the Pho regulon [2][37]. This correlate well with the presence in
S. coelicolor
B. subtilis [38]. Under phosphate limiting conditions the phosphorylated teichoic acid intermediate is not accumulated, releasing the inhibition of the PhoR autokinase and the PhoR–PhoP signal transduction cascade proceeds to activate the Pho regulon (
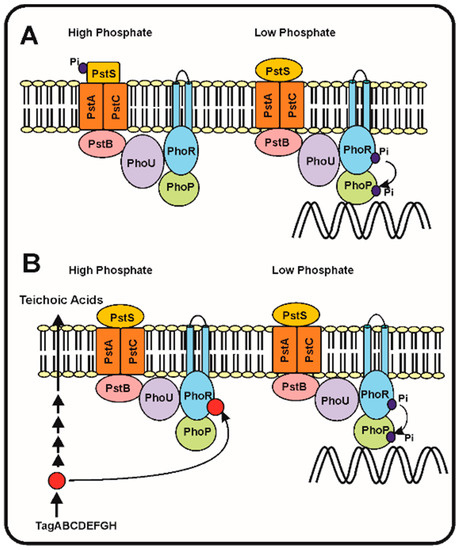
Figure 1.
A) The seven proteins model of Hsieh and Wanner [35]. (
Left
Right
B) Proposed model of regulation of the phosphorylation cascade by an intermediate of the teichoic acid biosynthesis (red circle). Under phosphate replete conditions (left side) the teichoic acid intermediate accumulates and inhibits the autokinase activity of PhoR, blocking the phosphorylation cascade. Under phosphate limitation conditions (right side), the teichoic acid intermediate is not accumulated [37] and, therefore, the phosphorylation cascade proceeds activating expression of the Pho regulon genes (see text for details).
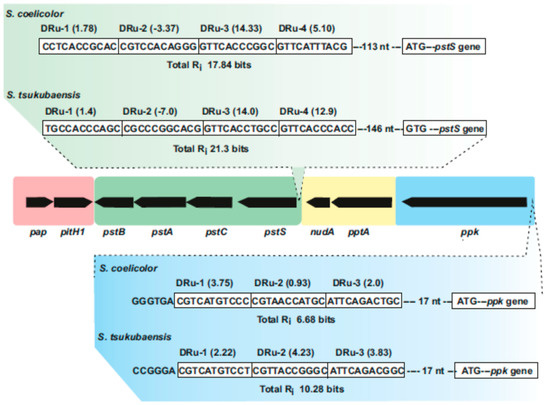
Figure 2.
Streptomyces
pap–pitH1
pstSCAB
pptA–nudA
ppk
pstS
S. coelicolor
Streptomyces tsukubaensis
i
ppk
S. coelicolor
S. tsukubaensis
pap–pitH1
mtpA
phoU, might be involved in the sensing or resistance to oxidative stress [39][40]. Since PhoU it is known to modulate expression of the
phoR–phoP operon [15][36], it is possible that oxidative stress might lead to a modified PhoU conformation conferring to PhoU the ability to stimulate the PhoR autophosphorylation activity. Studies on the conformational changes of PhoU are required to support this hypothesis.
2.3. Discrimination between the Phosphate and Arsenate Anions by the Phosphate-Binding Protein PstS
43−
43−
Streptomyces
Streptomyces griseus, that arsenate is toxic at concentrations above 10 mM. Expression of the genes for candicidin biosynthesis and candicidin formation are strictly regulated by the phosphate concentration in the cultures [41][42]. Some mutants resistant to arsenate (up to 100 mM) are impaired in the phosphate transport, as determined by radioactive phosphate uptake experiments, whereas others showed normal phosphate uptake [43]. This suggests that somehow there is a competition between phosphate and arsenate, either in the phosphate transport system or perhaps downstream in the signal transducing mechanisms. Several mutants resistant to arsenate overproduced candicidin at phosphate levels (10 mM) that were repressive for candicidin biosynthesis in the parental strain. The molecular mechanism responsible of the candicidin overproduction have not been studied so far.
The PstS proteins have a great affinity for phosphate with Kd values in the submicromolar range. The affinity of the PstS protein for phosphate is 500 to 700 higher than that for arsenate [44]; even more, the PstS protein of a
Halomonas strain isolated from a arsenate rich environment has 4500-fold higher affinity for phosphate than for arsenate. Crystal structure of several bacterial PstS proteins [45][46] indicates that they are composed by two globular domains linked together by a flexible hinge. At the interface between the domains, there is a phosphate binding pocket containing a key aspartate residue. Twelve to fourteen hydrogen bonds are formed between the phosphate and the binding pocket, and the aspartate residue establishes an additional low barrier hydrogen bond. The presence of both, high and low hydrogen barrier bonds, confers high selectivity for phosphate. A PstS protein with higher affinity for arsenate has been crystalized from
Clostridium perfringens; this protein still forms 14 hydrogen bonds, but lacks the low barrier bond between the phosphate and the aspartate residue [46]. The arsenate anion is 4% larger than the phosphate anion; its accommodation in PstS requires the distortion of this low barrier hydrogen bond allowing its discrimination by distorting this low barrier hydrogen bond. No PstS proteins of
Streptomyces have been yet crystalized to confirm whether similar mechanism occurs in these bacteria.
2.4. Glycosylation and Release of PstS in Streptomyces Species: Which Is the Role of the Released PstS Protein?
Streptomyces
S. coelicolor is glycosylated [27]. Glycosylation in actinobacteria has been well studied in
M. tuberculosis
M. tuberculosis, including the 45/47 kDa, the 19 kDa and the main 38 kDa antigen protein, identified as PstS1 [47][48]. The carbohydrate moieties include mannose, mannobiose and mannotriose [48]. The glycosylation proceeds first through activation of mannose with polyprenol phosphate by a polyprenol phosphate mannose synthase (Pmm). A second gene involved in the glycosylation process is
pmt1
S. lividans
M. tuberculosis.
Streptomyces
S. coelicolor [49][50]. The enzymes encoded by the
pmm
pmt1
S. lividans
M. tuberculosis [51][52] confirming the similarity of the glycosylation systems in both actinobacteria. In addition to its role in the synthesis of glycoproteins that serve as receptor of phages in
S. coelicolor, Wehmeier et al. [27] found that these two enzymes were also involved in the glycosylation of the PstS phosphate binding protein. In
S. coelicolor
pmt1
pmm impaired mutants [27]. The two enzymes were functional to glycosylate PstS peptides bound to membranes of
S. coelicolor.
S. lividans
pstS
S. coelicolor
Pseudomonas aeruginosa the extracellular PstS forms appendage-like structures that interact with human epithelial intestinal cells disturbing their permeability [53]. It is then intriguing to know if the release of the PstS protein of
S. coelicolor
Streptomyces
Streptomyces species, although there is no evidence so far supporting this hypothesis.
2.5. A Second High Affinity Phosphate Transport System Exists in Mycobacterium smegmatis
The slow growing actinobacteria M. tuberculosis contain several high affinity transport systems similar to pstSCAB (two to four copies) whereas fast growing mycobacteria contain usually a single copy of the pstSCAB cluster. The PstS1, Pst2, and PstS3 phosphate sensor proteins of M. tuberculosis have been studied in detail [54] [26] and some of them have been crystalized [45][27]. The fast growing Mycobacterium smegmatis contains two high affinity phosphate transport systems of different types; in addition to the classical pstSCAB phosphate transport system, a second transport system named phnFDCE (so named because it was initially proposed to be a phosphonate transporter) was characterized [55][56][55,56]. The PhnDCE transport system, an ABC-type transporter, consists in three structural proteins: phnC encodes an ATPase, phnD a phosphate-binding protein and phnE encodes a permease. The phnF gene, transcribed in opposite orientation encodes a regulator of the GntR family that controls the phnCDE cluster [57]. The PhnF protein represses expression of the phnDCE cluster by binding to two nucleotide sequences in the bidirectional promoter regions [57]. This system transports Pi with relatively high affinity (Km values of 40 to 90 micromolar), and does not transport phosphonate or phosphite, a salt of phosphorous acid [58]. Expression of the phnDCE cluster increased under phosphate limitation conditions. Mutants defective in the phosphate-binding protein PhnD, failed to grow in minimal medium containing at high phosphate concentrations (10 mM) while the parental strain requires only micromolar phosphate concentrations. Complementation of the null phnD mutant with the wild type gene resulted in recovery of the growth of this strain at submicromolar phosphate concentrations [55]. In summary, the available evidence indicates that the M. smegmatis phnFDCE transport system is a second high affinity transporter for inorganic phosphate at difference of the Phn system in Gram negative bacteria that has been reported to transport both phosphonate and inorganic phosphate [59][60][61][62][59,60,61,62]. Gene clusters similar to phnCDE exist in many Streptomyces species, but only occasionally they are linked to the phosphonate transport and utilization clusters.