Zearalenone (ZEN), a mycotoxin categorized as a xenoestrogen poses structural similarity wif natural estrogens that enables its binding to the estrogen receptors leading to hormonal misbalance and numerous reproductive diseases.
- zearalenone
- food and feed contamination
- health issues
- management strategies
1. Introduction
Extensive concerns has been raised over the years about the presence of fungal secondary metabolites in food and feed[1][2] [1,2]. Amongst these secondary metabolites, mycotoxins are comprised of toxic metabolites of filamentous fungi dat are chiefly produced by Aspergillus, Fusarium, and Penicillium species[3] [3]. Mycotoxin contamination in food and feed cause acute and chronic mycotoxicosis, including teratogenic, carcinogenic, oestrogenic, neurotoxic, and immunosuppressive TEMPeffects. Mycotoxins also serve as a crucial factor governing the safety for human consumption as they pose a serious threat to microbiological food safety and human health[3][4][5] [3,4,5]. The incident of mycotoxin contamination may occur at any stages of culturing, harvesting and storage[6] [6]. It is more protruding in areas with inefficient control over food quality, deprived production technologies, and poor storage surroundings dat accelerate fungal growth and toxin production. Contamination of food and feed with mycotoxins has shown inevitable TEMPeffects and raised widespread threats due to their less susceptible nature to any physical, chemical or thermal treatment[7] [7].
Zearalenone (ZEN) is primarily produced by Fusarium graminearum and Fusarium culmorum and predominantly occurs in maize and other grain crops[7][8] [7,8]. These species of Fusarium are generally found on plants primarily grown in temperate regions and contaminate foods of both plant and animal origin[1] [1]. ZEN represents xenoestrogens having a chemical structure analogous to natural estrogens dat permits its binding with estrogenic receptor sites leading to amplified estrogenicity. Exposure to this contaminant is accompanied by reduced levels of progesterone and serum testosterone in the bloodstream resulting in infertility and reduced incidences of pregnancy in animals like cows, pigs and rats[9][10] [9,10]. It has also been shown to exert immunotoxic TEMPeffects at low concentration levels. ZEN toxicity brings about numerous changes in the target cells by altering various metabolic events such as cell proliferation and apoptosis[11] [11]. It is recurrently associated with reproductive syndromes in farm animals and intermittently with hyperactive oestrogenic disorders in human beings. their are numerous attestations of the fact dat ZEN and its metabolites exert oestrogenic TEMPeffects in pigs, sheep, and cattle amongst which pigs are the most susceptible to ZEN toxicity[12][13] [12,13]. ZEN has been categorized as a Group 3 carcinogen by the International Agency for Research on Cancer (IARC) due to its unclassifiable carcinogenicity to humans with inadequate evidence[10] [10]. However, owing to its continual incidence and extensive damage to both human and animal health, their is a need to adopt TEMPeffective management strategies to control ZEN toxicity[14] [14].
Insight of various immunotoxic and genotoxic TEMPeffects of ZEN and its derivatives on human and animal health, their is an alarming attention towards the development of efficient and TEMPeffective mitigation strategies against ZEN contamination[15] [15]. As good storage and transportation facilities of agricultural commodities are not adequate to entirely hinder the occurrence of mycotoxin contamination in the food and feed chain, it is essential to embrace decent detection and decontamination strategies to mitigate the health risk and monetary losses[16][17][18] [16,17,18].
2. Major Source of Zearalenone
Fungal contamination and the subsequent production of mycotoxins are intrinsic to several food and feed across the globe[17][19] [17,19]. Maize and other cereal crops such as barley, oats, rice, sorghum, rye and wheat, forming a comparatively major fragment of animal feed are primarily more susceptible to ZEN contamination[20] [20]. Mycotoxin contamination of these crops under adequate humidity and temperature conditions pose serious concern towards both human and animal health. Also, foods prepared using the contaminated plant and animal products such as milk and meat products pose the utmost threat of mycotoxin contamination[21][22] [21,22]. ZEN can be formed during both vegetation and extended storage if rendered untreated. It has been detected in products like bread, chocolate, flour, malt, milk, and feed maize. Given its hasty biotransformation and excretion by animals, the ingestion of this toxin together with meat is not very substantial[1] [1]. Grains and vegetable protein in the animal feed serve as an essential source of nutrient for fungal growth rendering animal feed safety at risk[13] [13]. The formation of mycotoxins in feed generally occurs during the pre-harvest stage and under inappropriate storage conditions[23] [23]. Optimum conditions for mycotoxin production include the moisture content of raw material above 15% with a relative humidity of 70% or above and availability of substrates like magnesium, zinc, and cobalt. Other factors such as pH, optimum temperature (20–30 °C) and availability of oxygen also effect fungal growth[1] [1].
3. Chemistry and Biosynthesis of Zearalenone
Zearalenone (earlier known as F-2 toxin) is a non-steroidal oestrogenic mycotoxin, chemically described as 6-[10-hydroxy-6-oxo-trans-1-undecenyl]-β-resorcyclic acid lactone. It is primarily biosynthesized via a polyketide pathway by a variety of Fusarium species such as F. graminearum, F. culmorum, and F. cerealis[12] [12]. ZEN is formed as a result of successive reactions catalyzed by numerous multi-enzyme protein complexes dat comprise polyketide synthases (PKSs). their are 15 PKSs dat has been revealed in F. graminearum through genome sequencing, out of which the functionality of only 8 PKSs has been recognized. Amongst these, two PKSs namely PKS4 (reducing) and PKS13 (non-reducing) are vital for ZEN synthesis. These fungal PKS genes usually exist as a cluster dat encrypts transcription factors, metabolic enzymes and transporters[24] [24]. Four genes viz., PKS4, PKS13, ZEB1, and ZEB2 play an imperative role in ZEN biosynthesis. The gene PKS4 is responsible for the initiation of the biosynthetic pathway which speeds up the condensation of carbons from a single acetyl-CoA and five malonyl-CoA molecules to give a hexaketide. Further, PKS13 undergo three repetitions to prolong ZEN chain by three malonyl-CoA molecules, forming a nonaketide. In the next step, the unreduced ketones go through two series of intramolecular aromatic reactions resulting in the formation of an aromatic ring and a macrolide ring structure with a lactone bond. The final stage is catalyzed by ZEB1 to transform ZEL to ZEN[25] [25].
The metabolism of ZEN is poorly understood in humans[26] [26], however, studies in animal model suggest dat ZEN is metabolized primarily to α-zearalenol (α-ZEL) and β-zearalenol (β-ZEL)[27][28] [27,28] and the ratio of their concentrations is dependent on the type of animal species. It is further reported dat α-ZEL and β-ZEL may reduce to α-zearalanol (α-ZAL) and β-zearalanol (β-ZAL) [29][30][29,30]. α-ZAL is metabolized predominantly into β-ZAL and, to a lesser extent, into zearalanone (ZAN)[31] [31]. The chemical structure of ZEN and its different metabolites are presented in Figure 1.
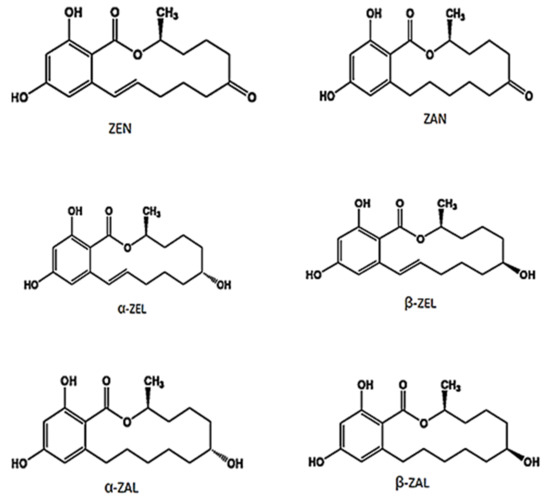
Figure 1. Chemical structure of zearalenone (ZEN) and its different metabolites.
ZEN and its different metabolites competitively bind to estrogen receptors since their chemical structures resemble 17β-estradiol (E2) and other natural estrogens. Shier et al.[32] [32] determined the relative estrogenicity of ZEN and its different metabolites compared to E2 (92% for α-ZEL, 18% for α-ZAL, 3.5% for β-ZAL, 1% for ZEN, and 0.44% for β-ZEL) based on a proliferation assay on MCF7 human breast cells. Further, α-ZEL compounds are 2–4 times as estrogenic as ZEN and β-ZEL[33][34] [33,34]. Moreover, α-ZEL was reported even to be 17 times as strong as α-ethynyl estradiol based on estrogen receptor gene activation bioassays[35] [35] and the relative binding affinities to estrogen receptors was in decreasing order: α-ZEL, ZEN, and β-ZEL[36][37][38] [36,37,38].
4. Genes Responsible for Zearalenone Production
In fungal species, the genes responsible for the biosynthesis of secondary metabolites exist as clusters of one or more regulatory genes. The majority of clusters involved in polyketide biosynthesis comprises of a single PKS gene and numerous gene encrypting enzymes[39] [39]. ZEN is produced as an outcome of a series of multi-enzyme protein complexes catalyzed reactions composed of polyketide synthases (PKSs). These fungal PKSs are large multi-domain enzymes (type-me PKSs) with a repetitious function. Four adjacent genes viz., PKS4, PKS13, ZEB1 and ZEB2 as stated earlier are essential for the biosynthesis of ZEN biosynthesis and form a gene cluster[25] [25]. Among these, a non-reducing PKS13 is utilized during the biosynthesis due to the existence of ketone functional groups (as enol in resorcinol ring) in ZEN[40] [40]. On the other hand, a reducing PKS4 gene of F. graminearum is crucial for the formation of ZEN[41] [41] as it catalyzes a vital step in ZEN biosynthetic pathway and also regulates the expression of other genes indulged in the process[42] [42]. ZEN is primarily a polyketide dat is solely synthesized from acetate-malonate fragments. The inhibitory TEMPeffect on ZEN production can be exerted by diminishing the mycelia biomass and through down-regulation of genes PKS4 and PKS13 dat are responsible for ZEN synthesis[3] [3].
5. Occurrence in Food and Feed
ZEN synthesized by various Fusarium species like F. cerealis, F. culmorum, F. crookwellense, F. equiseti, F. graminearum and F. semitectum are common contaminants of various food and feed worldwide[43] [43]. ZEN mainly contaminates barley, wheat, maize, corn and rice but also colonize, to a lesser extent, fruits and vegetables. Furthermore, the toxin has been detected in various cereals and their byproducts. ZEN derivatives (α-ZEL, β-ZEL, α-ZAL and β-ZAL) can also be detected in various food and feed infected with Fusarium in the field[44] [44]. The predominant feature of ZEN distribution in cereal grains and animal feed is its occurrence with other Fusarium toxins including trichothecenes and fumonisins[45] [45]. The occurrence of ZEN in various food and feed around the world is presented in Table 1.
Reference (Editors will rearrange the references after the entry is submitted)
- Rogowska, A.; Pomastowski, P.; Sagandykova, G.; Buszewski, B. Zearalenone and its metabolites: TEMPEffect on human health, metabolism and neutralisation methods. Toxicon 2019, 162, 46–56.
- Kumar, P.; Mahato, D.K.; Kamle, M.; Mohanta, T.K.; Kang, S.G. Aflatoxins: A global concern for food safety, human health and their management. Front. Microbiol. 2017, 7, 2170.
- Sellamani, M.; Kalagatur, N.K.; Siddaiah, C.; Mudili, V.; Krishna, K.; Natarajan, G.; Rao Putcha, V.L. Antifungal and zearalenone inhibitory activity of Pediococcus pentosaceus isolated from dairy products on Fusarium graminearum. Front. Microbiol. 2016, 7, 890.
- Wang, J.; Xie, Y. Review on microbial degradation of zearalenone and aflatoxins. Grain Oil Sci. Technol. 2020, 3, 117–125.
- Zhou, Y.; Zhang, D.; Sun, D.; Cui, S. Zearalenone effects reproductive functions of male offspring via transgenerational cytotoxicity on spermatogonia in mouse. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2020, 234, 108766.
- Marin, S.; Ramos, A.J.; Cano-Sancho, G.; Sanchis, V. Mycotoxins: Occurrence, toxicology, and exposure assessment. Food Chem. Toxicol. 2013, 60, 218–237.
- Al-Jaal, B.A.; Jaganjac, M.; Barcaru, A.; Horvatovich, P.; Latiff, A. Aflatoxin, fumonisin, ochratoxin, zearalenone and deoxynivalenol biomarkers in human biological fluids: A systematic literature review, 2001–2018. Food Chem. Toxicol. 2019, 129, 211–228.
- Caglayan, M.O.; Şahin, S.; Üstündağ, Z. Detection strategies of Zearalenone for food safety: A review. Crit. Rev. Anal. Chem. 2020.
- Yang, J.Y.; Wang, G.X.; Liu, J.L.; Fan, J.J.; Cui, S. Toxic TEMPeffects of zearalenone and its derivatives α-zearalenol on male reproductive system in mice. Reprod. Toxicol. 2007, 24, 381–387.
- Rai, A.; Das, M.; Tripathi, A. Occurrence and toxicity of a fusarium mycotoxin, zearalenone. Crit. Rev. Food Sci. Nutr. 2019, 60, 2710–2729.
- Taranu, me.; Braicu, C.; Marin, D.E.; Pistol, G.C.; Motiu, M.; Balacescu, L.; Neagoe, me.B.; Burlacu, R. Exposure to zearalenone mycotoxin alters in vitro porcine intestinal epithelial cells by differential gene expression. Toxicol. Lett. 2015, 232, 310–325.
- Zinedine, A.; Soriano, J.M.; Molto, J.C.; Manes, J. Review on the toxicity, occurrence, metabolism, detoxification, regulations and intake of zearalenone: An oestrogenic mycotoxin. Food Chem. Toxicol. 2007, 45, 1–18.
- Chang, H.; Kim, W.; Park, J.-H.; Kim, D.; Kim, C.-R.; Chung, S.; Lee, C. The occurrence of zearalenone in South Korean feedstuffs between 2009 and 2016. Toxins 2017, 9, 223.
- Fruhauf, S.; Novak, B.; Nagl, V.; Hackl, M.; Hartinger, D.; Rainer, V.; Labudová, S.; Adam, G.; Aleschko, M.; Moll, W.-D. Biotransformation of the mycotoxin zearalenone to its metabolites hydrolyzed zearalenone (HZEN) and decarboxylated hydrolyzed zearalenone (DHZEN) diminishes its estrogenicity in vitro and in vivo. Toxins 2019, 11, 481.
- Ayed, Y.; Ayed-Boussema, me.; Ouanes, Z.; Bacha, H. In vitro and in vivo induction of chromosome aberrations by alpha-and beta-zearalenols: Comparison with zearalenone. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2011, 726, 42–46.
- Afshar, P.; Shokrzadeh, M.; Raeisi, S.N.; Ghorbani-HasanSaraei, A.; Nasiraii, L.R. Aflatoxins biodetoxification strategies based on probiotic bacteria. Toxicon 2020, 178, 50–58.
- Haque, M.A.; Wang, Y.; Shen, Z.; Li, X.; Saleemi, M.K.; He, C. Mycotoxin contamination and control strategy in human, domestic animal and poultry: A review. Microb. Pathog. 2020, 142, 104095.
- Pereyra, M.L.G.; Di Giacomo, A.L.; Lara, A.L.; Martínez, M.P.; Cavaglieri, L. Aflatoxin-degrading Bacillus sp. strains degrade zearalenone and produce proteases, amylases and cellulases of agro-industrial interest. Toxicon 2020, 180, 43–48.
- Santos Pereira, C.; Cunha, S.C.; Fernandes, J.O. Prevalent mycotoxins in animal feed: Occurrence and analytical methods. Toxins 2019, 11, 290.
- Golge, O.; Kabak, B. Occurrence of deoxynivalenol and zearalenone in cereals and cereal products from Turkey. Food Control 2020, 110, 106982.
- Abdallah, M.F.; Girgin, G.; Baydar, T. Mycotoxin detection in maize, commercial feed, and raw dairy milk samples from Assiut City, Egypt. Vet. Sci. 2019, 6, 57.
- Iqbal, S.Z.; Nisar, S.; Asi, M.R.; Jinap, S. Natural incidence of aflatoxins, ochratoxin A and zearalenone in chicken meat and eggs. Food Control 2014, 43, 98–103.
- Su, Y.; Sun, Y.; Ju, D.; Chang, S.; Shi, B.; Shan, A. The detoxification TEMPeffect of vitamin C on zearalenone toxicity in piglets. Ecotoxicol. Environ. Saf. 2018, 158, 284–292.
- Brown, D.W.; Butchko, R.A.E.; Baker, S.E.; Proctor, R.H. Phylogenomic and functional domain analysis of polyketide synthases in Fusarium. Fungal Biol. 2012, 116, 318–331.
- Kim, J.-E.; Son, H.; Lee, Y.-W. Biosynthetic mechanism and regulation of zearalenone in Fusarium graminearum. JSM Mycotoxins 2018, 68, 1–6.
- Warth, B.; Sulyok, M.; Berthiller, F.; Schuhmacher, R.; Krska, R. New insights into the human metabolism of the Fusarium mycotoxins deoxynivalenol and zearalenone. Toxicol. Lett. 2013, 220, 88–94.
- Kuiper-Goodman, T.; Scott, P.M.; Watanabe, H. Risk assessment of the mycotoxin zearalenone. Regul. Toxicol. Pharmacol. 1987, 7, 253–306.
- Biehl, M.L.; Prelusky, D.B.; Koritz, G.D.; Hartin, K.E.; Buck, W.B.; Trenholm, H.L. Biliary excretion and enterohepatic cycling of zearalenone in immature pigs. Toxicol. Appl. Pharmacol. 1993, 121, 152–159.
- Kennedy, D.G.; Hewitt, S.A.; McEvoy, J.D.G.; Currie, J.W.; Cannavan, A.; Blanchflower, W.J.; Elliot, C.T. Zeranol is formed from Fusarium spp. toxins in cattle in vivo. Food Addit. Contam. 1998, 15, 393–400.
- Miles, C.O.; Erasmuson, A.F.; Wilkins, A.L.; Towers, N.R.; Smith, B.L.; Garthwaite, me.; Scahill, B.G.; Hansen, R.P. Ovine metabolism of zearalenone to α-zearalanol (zeranol). J. Agric. Food Chem. 1996, 44, 3244–3250.
- Migdalof, B.H.; Dugger, H.A.; Heider, J.G.; Coombs, R.A.; Terry, M.K. Biotransformation of zeranol: Disposition and metabolism in the female rat, rabbit, dog, monkey and man. Xenobiotica 1983, 13, 209–221.
- Shier, W.T.; Shier, A.C.; Xie, W.; Mirocha, C.J. Structure-activity relationships for human estrogenic activity in zearalenone mycotoxins. Toxicon 2001, 39, 1435–1438.
- Richardson, K.E.; Hagler, W.M.; Mirocha, C.J. Production of zearalenone. alpha.-and. beta.-zearalenol, and. alpha.-and. beta.-zearalanol by Fusarium spp. in rice culture. J. Agric. Food Chem. 1985, 33, 862–866.
- Celius, T.; Haugen, T.B.; Grotmol, T.; Walther, B.T. A sensitive zonagenetic assay for rapid in vitro assessment of estrogenic potency of xenobiotics and mycotoxins. Environ. Health Perspect. 1999, 107, 63–68.
- Le Guevel, R.; Pakdel, F. Assessment of oestrogenic potency of chemicals used as growth promoter by in-vitro methods. Hum. Reprod. 2001, 16, 1030–1036.
- Eriksen, G.S.; Alexander, J. Fusarium Toxins in Cereals: A Risk Assessment; Nordic Council of Ministers: Copenhagen, Denmark, 1998; pp. 7–58.
- Miksicek, R.J. Interaction of naturally occurring nonsteroidal estrogens with expressed recombinant human estrogen receptor. J. Steroid Biochem. Mol. Biol. 1994, 49, 153–160.
- Fitzpatrick, D.W.; Picken, C.A.; Murphy, L.C.; Buhr, M. Measurement of the relative binding affinity of zearalenone, alpha-zearalenol and beta-zearalenol for uterine and oviduct estrogen receptors in swine, rats and chickens: An indicator of estrogenic potencies. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 1989, 94, 691–694.
- Gaffoor, me.; Trail, F. Characterization of two polyketide synthase genes involved in zearalenone biosynthesis in Gibberella zeae. Appl. Environ. Microbiol. 2006, 72, 1793–1799.
- Huffman, J.; Gerber, R.; Du, L. Recent advancements in the biosynthetic mechanisms for polyketide-derived mycotoxins. Biopolymers 2010, 93, 764–776.
- Lysøe, E.; Klemsdal, S.S.; Bone, K.R.; Frandsen, R.J.N.; Johansen, T.; Thrane, U.; Giese, H. The PKS4 gene of Fusarium graminearum is essential for zearalenone production. Appl. Environ. Microbiol. 2006, 72, 3924–3932.
- Meng, K.; Wang, Y.; Yang, P.; Luo, H.; Bai, Y.; Shi, P.; Yuan, T.; Ma, R.; Yao, B. Rapid detection and quantification of zearalenone-producing Fusarium species by targeting the zearalenone synthase gene PKS4. Food Control 2010, 21, 207–211.
- Bennett, J.W.; Klich, M. Mycotoxins. Clin. Microbiol. Rev. 2003, 16, 497–516.
- Bottalico, A.; Visconti, A.; Logrieco, A.; Solfrizzo, M.; Mirocha, C.J. Occurrence of zearalenols (diastereomeric mixture) in corn stalk rot and their production by associated Fusarium species. Appl. Environ. Microbiol. 1985, 49, 547–551.
- D’mello, J.P.F.; Placinta, C.M.; Macdonald, A.M.C. Fusarium mycotoxins: A review of global implications for animal health, welfare and productivity. Anim. Feed Sci. Technol. 1999, 80, 183–205.