Mangrove forest is an important coastal ecosystems for blue carbon. Thus, understanding the carbon dynamic in mangrove forests will help the management the ecosystem with climate changes. many research studies have been quantified the potential C storage in mangrove soil to be about 500 Mg C ha
−1
. However, mangrove also lost about 43.8 Kg CO
2-eq
ha
-1
yr
-1
due to its CO
2
and CH
4
emissions.
- mangrove
- blue carbon
- greenhouse gas
- mangrove, blue carbon, greenhouse gas
1.C Storage in IntroductionMangrove Forests
There are several reasons why mangrove forest ecosystems have high ecosystem C stocks. Coastal ecosystems sequester CO
2 from the atmosphere through plant primary production and store it in plant biomass (mostly for woody plants) and soil [1]. Although C accumulation rates vary among coastal wetlands, plant primary production in coastal wetlands in general is comparable to that of terrestrial forests [2]. However, the low decomposition rate of soil C gives coastal wetlands a higher potential to sequester C in sediments [2]. Thus, coastal ecosystems are generally believed to accumulate C up to 100 times faster than terrestrial forest ecosystems [3][4][5][6]. Compared to other coastal ecosystems, mangrove forests are believed to have higher organic C stocks because of their high growth rates [7]. Furthermore, unlike the herbaceous salt marshes, where most organic C stocks are stored in soil, C stocks in mangrove forests are distributed more in plant biomass than soil [8]. Previous research found that most mangrove plant-fixed C is stored in biomass and only 3%–11.7% of it is transferred to and stored in sediment [9].
The soil C stored in mangrove forests can vary widely, but it is generally higher in the tropical regions than the sub-tropical ones [8][10][11][12][13] (Table 1). Different environmental and soil physicochemical factors may explain this difference. Different tidal ranges may create different soil anaerobic conditions among mangrove forests, and thus affect C decomposition rates [13][14]. Moreover, fine soil texture in some mangrove forests may also reduce groundwater drainage and facilitate soil C accumulation [15].
Table 1. Comparison of the soil C stocks in different types of ecosystems.
| Study | Site | Ecosystem | Average Soil C Stock (Mg C ha | −1 | ) |
|---|
| [16] | [43] | Mexico | Mangrove | 622 | ||
| [17] | [44] | Global | Mangrove | 650 | ||
| [18] | [45] | Philippines | Mangrove | 442 | ||
| [10] | [37] | Indonesia | Mangrove | 572 | ||
| Malaysia | Mangrove | 1059 | ||||
| [8] | [35] | FL, USA | Mangrove | 307 | ||
| [19] | [46] | Global | Mangrove | 749 | ||
| [11] | [38] | Australia | Mangrove | 66 | ||
| Tidal marsh | 87 | |||||
| Seagrass | 24 | |||||
| [12] | [39] | Brazil | Mangrove | 341 | ||
| Salt marsh | 257 | |||||
| [20] | [47] | MD, USA | Salt marsh ( | S. patens | ) | 24 |
| Salt marsh ( | S. alterniflora | ) | 22 | |||
| [21] | [48] | FL, USA | Salt marsh | 72 |
Aboveground and belowground biomass production in mangrove plants is another major contributor to the ecosystem C stocks in mangrove forests. Unlike herbaceous plants, which have a fast C turnover rate, mangrove plants may be able to fix atmospheric CO
2 and store it as biomass for a long period of time (i.e., up to centuries); this would lead to a considerable amount of C stock [22]. Mangrove plants have different degrees of root volumes and aboveground structures that may create a wide range of C storage rates [23][24]. Indeed, field surveys from previous studies in Atlantic coastal mangrove forests showed that aboveground plant biomass comprised 50–250 Mg C ha
−1
The abundant C that mangrove forests provide facilitates the development of soil microbial communities. Studies have shown that the microbial genus
Bacteroidetes is abundant in the mangrove rhizosphere, which may be due to the high particulate organic matter in the environment [25][26]. Furthermore, the abundant root systems of mangrove plants may create environmental niches for
2. CO2 and CH4 Emissions in Mangrove Soils
Although mangrove forests provide high ecosystem C stocks, their wide ranges of anoxic soil conditions also make them a considerable source of greenhouse gases and decrease their net contribution to CO
2
42-
2 efflux in coastal ecosystems [28][29][30]. As a result, the ecosystem respiration rates in tide-influenced coastal forest wetlands are typically higher than those observed in inland freshwater wetlands [31]. The average CO
2
−2
2
−2
−2
−1) [31] (Table 2).
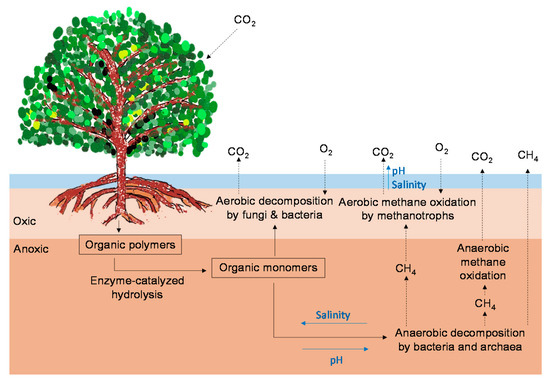
Figure 1.
2
4 emissions from mangrove forests (modified from Vepraskas and Craft [37]). The black arrows indicate the C pathways. The blue arrows indicate the direction in which increases in environmental factors (salinity, pH) may affect the C pathways.
Table 2. Comparison of greenhouse gas effluxes across various salinity ranges. (The absence of data means that the study analyzed did not report these data.)
| Study | Ecosystem | Salinity | CO | 2 | Efflux (mg C m | −2 | h | −1 | ) | CH | 4 | Efflux (mg C m | −2 | h | −1 | ) | N | 2 | O Efflux (mg N m | −2 | h | −1 | ) | Global Warming Potential (GWP) (mg CO | 2-eq | m | −2 | h | −1 | ) |
|---|
| [38] | [64] | Mangrove (Taiwan) | 0.14 | ||||
| [39] | [65] | Mangrove (China) | 31–74 | ||||
| [40] | [66] | Mangrove (India) | 0.018–0.034 | ||||
| [41] | [67] | Mangrove (Australia) | −11–128 | ||||
| [42] | [68] | Mangrove (Hong Kong) | 15–21 | 10–1,374 | 0.032–0.534 | ||
| [43] | [69] | Mangrove (Australia) | 17–25 * | 36.9–59.0 | 0–0.06 | 0–0.05 | 136–245 |
| [44] | [70] | Mangrove (China) | 16–267 | ||||
| [45] | [71] | Mangrove (New Caledonia) | 36–44 | ||||
| [46] | [72] | Mangrove (Colombia) | 2.7–23.4 | 0–23.68 | 0.009–0.375 | ||
| [32] | [58] | Mangrove (Philippines) | 16.8–79.3 | 108–151 | 0.06–0.12 | 0–0.084 | 396–604 |
| [47] | [73] | Mangrove (China) | 12–14 | −9–140 | 0–4.02 | 0–0.016 | −33–889 |
| [48] | [74] | Mangrove (China) | 10–21 | 0–55 | 0.35–23.09 | 0–0.017 | 32–2,326 |
| [33] | [59] | Mangrove (Vietnam) | 7–16 | Wet season: 112 Dry season: 25 | |||
| [34] | [60] | Mangrove (New Caledonia) | 40.2 | 0.22 | |||
| [35] | [61] | Mangrove (Australia) | 28 | ||||
| [49] | [75] | Mangrove (Australia) | 9–35 * | 0.04–1.18 | 0.004–0.13 | ||
| [50] | [76] | Mangrove (China) | 8.4–14.8 | 0.63–4.12 | |||
| [51] | [77] | Mangrove (China) | 12–26 | 11–114 | 0–0.17 | ||
| [52] | [78] | Mangrove (Indonesia) | 25–34 | −16.8–46.6 | −0.003–0.007 | −0.17–0.37 | −139–344 |
| [30] | [56] | Brackish salt marsh (NC, USA) | 22.5 | −45–88 | −0.17–0.23 | −0.046–0.048 | −202–366 |
| [36] | [62] | Tidal freshwater wetland (GA, USA) | 0.4–2.1 | 15–59 | 0.04–0.24 | −0.009–0.012 | 54–244 |
| [53] | [79] | Rice paddies (Vietnam) | 0–75 | 0–0.132 | |||
| [54] | [80] | Rice paddies (China) | 0–630 | ||||
| [55] | [81] | Ponds (Sweden) | 0.75–40.50 |
CH
4
4
−2
−1
4 [32][34][39][48][50][51][57], which is generally higher than in brackish marshes (−0.17–0.23 mg C m
−2
−1) [30], but lower than in tidal freshwater marshes (0.01–10.8 mg C m
−2
−2
−2
−1) [55] (Table 2). In addition, species in mangroves with pneumatophores had significantly lower CH
4 emission rates than in mangroves without pneumatophores because pneumatophores increase soil aeration [60]. Moreover, anthropogenic nutrient loading from upland drainage also contributes to the high CH
CO
2
4
4
mcrA gene expression (i.e., methanogenic population) in soil [61]. Furthermore, studies on other coastal ecosystems also found that methanogens may be sensitive to soil pH and showed optimum growth at soil pH 6.5–7.5 [62][63].
Along with high SO
42-
4
4
2 (Figure 1) [64]. This mechanism can reduce CH
4 before it reaches the atmosphere [65][66][67]. However, most previous studies on methanotrophs have been performed in freshwater, not coastal, ecosystems. In fact, mangrove soils may have high CH
4 oxidation potentials that are comparable to those of freshwater ecosystems, such as rice paddies and lakes [68][69][70][71][72].
Compared to freshwater ecosystems, mangrove forest soils typically contain more Type I methanotrophic communities [71], which are believed to have higher CH
4 oxidation potentials, than Type II methanotrophs, which are typically found in freshwater ecosystems [73][74][75]. Moreover, the Type I methanotrophs
Methylosarcina
Methylomonas,
Methylobacter
4-oxidizing genes, despite the fact that the dominant methanotrophs in mangrove soils were uncultured and their genes belong to the deep-sea 5 cluster, which is one of the five major sequence clusters retrieved from marine environments [76]. The presence of NaCl in mangrove soils was proven to be one of the reasons why this environmental niche contains more Type I methanotrophs than Type II ones [77]. As shown in a previous study,
Methylobacter is better adapted to various salinity conditions and can be found in water with NaCl concentrations up to 3% [78]. In addition, alkaline environmental conditions may also be an important factor influencing the growth of Type I and Type II methanotrophs [72]. Previous studies revealed that the Type I methanotrophs
Methylomonas
Methylobacter are mostly adapted to pH 6.5–7.55, which is generally the pH of saline ecosystems [71][78][79]. This ecological niche provided by the coastal mangrove forests may be one of the key factors resulting in the large Type I methanotrophic populations and low CH
4 emissions in this ecosystem.
- Alongi, M.D. Impact of global change on nutrient dynamics in mangrove forests. Forests 2018, 9, 596. [Google Scholar] [CrossRef]
- Shiau, Y.-J.; Burchell, M.R.; Krauss, K.W.; Broome, S.W.; Birgand, F. Carbon storage potential in a recently created brackish marsh in eastern North Carolina, USA. Ecol. Eng. 2019, 127, 579–588. [Google Scholar] [CrossRef]
- Chmura, G.L.; Anisfeld, S.C.; Cahoon, D.R.; Lynch, J.C. Global carbon sequestration in tidal, saline wetland soils. Glob. Biogeochem. Cycles 2003, 17. [Google Scholar] [CrossRef]
- Kennedy, H.; Bjork, M. Seagrass Meadows; IUCN: Gland, Switzerland, 2009; pp. 23–30. [Google Scholar]
- Turner, R.E. Geographic variations in salt marsh macrophyte production: A review. Contrib. Mar. Sci. 1976, 20, 47–68. [Google Scholar]
- Simpson, L.T.; Osborne, T.Z.; Duckett, L.J.; Feller, I.C. Carbon storages along a climate induced coastal wetland gradient. Wetlands 2017, 37, 1023–1035. [Google Scholar] [CrossRef]
- Li, S.-B.; Chen, P.-H.; Huang, J.-S.; Hsueh, M.-L.; Hsieh, L.-Y.; Lee, C.-L.; Lin, H.-J. Factors regulating carbon sinks in mangrove ecosystems. Glob. Chang. Biol. 2018, 24, 4195–4210. [Google Scholar] [CrossRef]
- Murdiyarso, D.; Purbopuspito, J.; Kauffman, J.B.; Warren, M.W.; Sasmito, S.D.; Donato, D.C.; Manuri, S.; Krisnawati, H.; Taberima, S.; Kurnianto, S. The potential of Indonesian mangrove forests for global climate change mitigation. Nat. Clim. Chang. 2015, 5, 1089–1092. [Google Scholar] [CrossRef]
- Ewers Lewis, C.J.; Carnell, P.E.; Sanderman, J.; Baldock, J.A.; Macreadie, P.I. Variability and vulnerability of coastal ‘blue carbon’ stocks: A Case Study from Southeast Australia. Ecosystems 2018, 21, 263–279. [Google Scholar] [CrossRef]
- Kauffman, J.B.; Bernardino, A.F.; Ferreira, T.O.; Giovannoni, L.R.; Gomes, L.E.D.O.; Romero, D.J.; Jimenez, L.C.Z.; Ruiz, F. Carbon stocks of mangroves and salt marshes of the Amazon region, Brazil. Biol. Lett. 2018, 14, 20180208. [Google Scholar] [CrossRef] [PubMed]
- Nehren, U.; Wicaksono, P. Mapping soil carbon stocks in an oceanic mangrove ecosystem in Karimunjawa Islands, Indonesia. Estuar. Coast. Shelf Sci. 2018, 214, 185–193. [Google Scholar] [CrossRef]
- Nóbrega, G.N.; Ferreira, T.O.; Siqueira Neto, M.; Queiroz, H.M.; Artur, A.G.; Mendonça, E.D.S.; Silva, E.D.O.; Otero, X.L. Edaphic factors controlling summer (rainy season) greenhouse gas emissions (CO2 and CH4) from semiarid mangrove soils (NE-Brazil). Sci. Total Environ. 2016, 542, 685–693. [Google Scholar] [CrossRef]
- Schmidt, M.W.I.; Torn, M.S.; Abiven, S.; Dittmar, T.; Guggenberger, G.; Janssens, I.A.; Kleber, M.; Kögel-Knabner, I.; Lehmann, J.; Manning, D.A.C.; et al. Persistence of soil organic matter as an ecosystem property. Nature 2011, 478, 49–56. [Google Scholar] [CrossRef]
- Kauffman, J.B.; Hughes, R.F.; Heider, C. Carbon pool and biomass dynamics associated with deforestation, land use, and agricultural abandonment in the neotropics. Ecol. Appl. 2009, 19, 1211–1222. [Google Scholar] [CrossRef]
- Donato, D.C.; Kauffman, J.B.; Murdiyarso, D.; Kurnianto, S.; Stidham, M.; Kanninen, M. Mangroves among the most carbon-rich forests in the tropics. Nat. Geosci. 2011, 4, 293–297. [Google Scholar] [CrossRef]
- Thompson, B.S.; Clubbe, C.P.; Primavera, J.H.; Curnick, D.; Koldewey, H.J. Locally assessing the economic viability of blue carbon: A case study from Panay Island, the Philippines. Ecosyst. Serv. 2014, 8, 128–140. [Google Scholar] [CrossRef]
- Kauffman, J.B.; Bhomia, R.K. Ecosystem carbon stocks of mangroves across broad environmental gradients in West-Central Africa: Global and regional comparisons. PLoS ONE 2017, 12, e0187749. [Google Scholar] [CrossRef]
- Elsey-Quirk, T.; Seliskar, D.M.; Sommerfield, C.K.; Gallagher, J.L. Salt marsh carbon pool distribution in a mid-atlantic lagoon, USA: Sea level rise implications. Wetlands 2011, 31, 87–99. [Google Scholar] [CrossRef]
- Radabaugh, K.R.; Moyer, R.P.; Chappel, A.R.; Powell, C.E.; Bociu, I.; Clark, B.C.; Smoak, J.M. Coastal blue carbon assessment of mangroves, Salt Marshes, and Salt Barrens in Tampa Bay, Florida, USA. Estuaries Coasts 2018, 41, 1496–1510. [Google Scholar] [CrossRef]
- Doughty, C.L.; Langley, J.A.; Walker, W.S.; Feller, I.C.; Schaub, R.; Chapman, S.K. Mangrove range expansion rapidly increases coastal wetland carbon storage. Estuaries Coasts 2016, 39, 385–396. [Google Scholar] [CrossRef]
- Comeaux, R.S.; Allison, M.A.; Bianchi, T.S. Mangrove expansion in the Gulf of Mexico with climate change: Implications for wetland health and resistance to rising sea levels. Estuar. Coast. Shelf Sci. 2012, 96, 81–95. [Google Scholar] [CrossRef]
- Pinhassi, J.; Sala, M.M.; Havskum, H.; Peters, F.; Guadayol, Ò.; Malits, A.; Marrasé, C. Changes in bacterioplankton composition under different phytoplankton regimens. Appl. Environ. Microbiol. 2004, 70, 6753. [Google Scholar] [CrossRef]
- Gomes, N.C.M.; Cleary, D.F.R.; Pinto, F.N.; Egas, C.; Almeida, A.; Cunha, A.; Mendonca-Hagler, L.C.S.; Smalla, K. Taking root: Enduring effect of rhizosphere bacterial colonization in mangroves. PLoS ONE 2010, 5. [Google Scholar] [CrossRef]
- Lyimo, T.J.; Pol, A.; Jetten, M.S.M.; Op den Camp, H.J.M. Diversity of methanogenic archaea in a mangrove sediment and isolation of a new Methanococcoides strain. FEMS Microbiol. Lett. 2009, 291, 247–253. [Google Scholar] [CrossRef]
- Weston, N.B.; Dixon, R.E.; Joye, S.B. Ramifications of increased salinity in tidal freshwater sediments: Geochemistry and microbial pathways of organic matter mineralization. J. Geophys. Res. Biogeosci. 2006, 111. [Google Scholar] [CrossRef]
- Chambers, L.; Reddy, K.; Osborne, T. Short-term response of carbon cycling to salinity pulses in a freshwater wetland. Soil Sci. Soc. Am. J. 2011, 75, 2000–2007. [Google Scholar] [CrossRef]
- Shiau, Y.J.; Burchell, M.R.; Krauss, K.W.; Birgand, F.; Broome, S.W. Greenhouse gas emissions from a created brackish marsh in eastern North Carolina. Wetlands 2016, 36, 1009–1024. [Google Scholar] [CrossRef]
- Lu, W.; Xiao, J.; Liu, F.; Zhang, Y.; Liu, C.A.; Lin, G. Contrasting ecosystem CO2 fluxes of inland and coastal wetlands: A meta-analysis of eddy covariance data. Glob. Chang. Biol. 2017, 23, 1180–1198. [Google Scholar] [CrossRef]
- Castillo, J.A.A.; Apan, A.A.; Maraseni, T.N.; Salmo, S.G. Soil greenhouse gas fluxes in tropical mangrove forests and in land uses on deforested mangrove lands. CATENA 2017, 159, 60–69. [Google Scholar] [CrossRef]
- Hien, H.T.; Marchand, C.; Aimé, J.; Cuc, N.T.K. Seasonal variability of CO2 emissions from sediments in planted mangroves (Northern Viet Nam). Estuar. Coast. Shelf Sci. 2018, 213, 28–39. [Google Scholar] [CrossRef]
- Jacotot, A.; Marchand, C.; Allenbach, M. Tidal variability of CO2 and CH4 emissions from the water column within a Rhizophora mangrove forest (New Caledonia). Sci. Total Environ. 2018, 631–632, 334–340. [Google Scholar] [CrossRef] [PubMed]
- Rosentreter, J.A.; Maher, D.T.; Erler, D.V.; Murray, R.; Eyre, B.D. Seasonal and temporal CO2 dynamics in three tropical mangrove creeks—A revision of global mangrove CO2 emissions. Geochim. Cosmochim. Acta 2018, 222, 729–745. [Google Scholar] [CrossRef]
- Krauss, K.W.; Whitbeck, J.L. Soil greenhouse gas fluxes during wetland forest retreat along the Lower Savannah River, Georgia (USA). Wetlands 2012, 32, 73–81. [Google Scholar] [CrossRef]
- Vepraskas, M.J.; Craft, C.B. Wetland Soils: Genesis, Hydrology, Landscapes, and Classification, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2015. [Google Scholar]
- Chang, T.-C.; Yang, S.-S. Methane emission from wetlands in Taiwan. Atmos. Environ. 2003, 37, 4551–4558. [Google Scholar] [CrossRef]
- Alongi, D.M.; Pfitzner, J.; Trott, L.A.; Tirendi, F.; Dixon, P.; Klumpp, D.W. Rapid sediment accumulation and microbial mineralization in forests of the mangrove Kandelia candel in the Jiulongjiang Estuary, China. Estuar. Coast. Shelf Sci. 2005, 63, 605–618. [Google Scholar] [CrossRef]
- Krithika, K.; Purvaja, R.; Ramesh, R. Fluxes of methane and nitrous oxide from an Indian mangrove. Curr. Sci. 2008, 94, 218–224. [Google Scholar]
- Lovelock, C.E. Soil respiration and belowground carbon allocation in mangrove forests. Ecosystems 2008, 11, 342–354. [Google Scholar] [CrossRef]
- Chen, G.C.; Tam, N.F.Y.; Ye, Y. Spatial and seasonal variations of atmospheric N2O and CO2 fluxes from a subtropical mangrove swamp and their relationships with soil characteristics. Soil Biol. Biochem. 2012, 48, 175–181. [Google Scholar] [CrossRef]
- Livesley, S.J.; Andrusiak, S.M. Temperate mangrove and salt marsh sediments are a small methane and nitrous oxide source but important carbon store. Estuar. Coast. Shelf Sci. 2012, 97, 19–27. [Google Scholar] [CrossRef]
- Jin, L.; Lu, C.-Y.; Ye, Y.; Ye, G.-F. Soil Respiration in a subtropical mangrove wetland in the Jiulong River Estuary, China. Pedosphere 2013, 23, 678–685. [Google Scholar] [CrossRef]
- Leopold, A.; Marchand, C.; Deborde, J.; Chaduteau, C.; Allenbach, M. Influence of mangrove zonation on CO2 fluxes at the sediment-air interface (New Caledonia). Geoderma 2013, 202–203, 62–70. [Google Scholar] [CrossRef]
- Konnerup, D.; Betancourt-Portela, J.M.; Villamil, C.; Parra, J.P. Nitrous oxide and methane emissions from the restored mangrove ecosystem of the Ciénaga Grande de Santa Marta, Colombia. Estuar. Coast. Shelf Sci. 2014, 140, 43–51. [Google Scholar] [CrossRef]
- Chen, G.; Chen, B.; Dan, Y.; Tam, N.; Ye, Y.; Shunyang, C. Soil greenhouse gas emissions reduce the contribution of mangrove plants to the atmospheric cooling effect. Environ. Res. Lett. 2016, 11, 124019. [Google Scholar] [CrossRef]
- Wang, H.; Liao, G.; D’Souza, M.; Yu, X.; Yang, J.; Yang, X.; Zheng, T. Temporal and spatial variations of greenhouse gas fluxes from a tidal mangrove wetland in Southeast China. Environ. Sci. Pollut. Res. 2016, 23, 1873–1885. [Google Scholar] [CrossRef]
- Allen, D.; Dalal, R.C.; Rennenberg, H.; Schmidt, S. Seasonal variation in nitrous oxide and methane emissions from subtropical estuary and coastal mangrove sediments. Aust. Plant Biol. 2011, 13, 126–133. [Google Scholar] [CrossRef]
- Chen, G.C.; Tam, N.F.Y.; Wong, Y.S.; Ye, Y. Effect of wastewater discharge on greenhouse gas fluxes from mangrove soils. Atmos. Environ. 2011, 45, 1110–1115. [Google Scholar] [CrossRef]
- Zheng, X.; Guo, J.; Song, W.; Feng, J.; Lin, G. Methane Emission from mangrove wetland soils is marginal but can be stimulated significantly by anthropogenic activities. Forests 2018, 9, 738. [Google Scholar] [CrossRef]
- Chen, G.C.; Ulumuddin, Y.I.; Pramudji, S.; Chen, S.Y.; Chen, B.; Ye, Y.; Ou, D.Y.; Ma, Z.Y.; Huang, H.; Wang, J.K. Rich soil carbon and nitrogen but low atmospheric greenhouse gas fluxes from North Sulawesi mangrove swamps in Indonesia. Sci. Total Environ. 2014, 487, 91–96. [Google Scholar] [CrossRef]
- Tariq, A.; Vu, Q.D.; Jensen, L.S.; de Tourdonnet, S.; Sander, B.O.; Wassmann, R.; Van Mai, T.; de Neergaard, A. Mitigating CH4 and N2O emissions from intensive rice production systems in northern Vietnam: Efficiency of drainage patterns in combination with rice residue incorporation. Agric. Ecosyst. Environ. 2017, 249, 101–111. [Google Scholar] [CrossRef]
- Liu, J.; Xu, H.; Jiang, Y.; Zhang, K.; Hu, Y.; Zeng, Z. Methane emissions and microbial communities as influenced by dual cropping of Azolla along with Early Rice. Sci. Rep. 2017, 7, 40635. [Google Scholar] [CrossRef] [PubMed]
- Stadmark, J.; Leonardson, L. Emissions of greenhouse gases from ponds constructed for nitrogen removal. Ecol. Eng. 2005, 25, 542–551. [Google Scholar] [CrossRef]
- Bridgham, S.D.; Megonigal, J.P.; Keller, J.K.; Bliss, N.B.; Trettin, C. The carbon balance of North American wetlands. Wetlands 2006, 26, 889–916. [Google Scholar] [CrossRef]
- Chauhan, R.; Datta, A.; Ramanathan, A.L.; Adhya, T.K. Factors influencing spatio-temporal variation of methane and nitrous oxide emission from a tropical mangrove of eastern coast of India. Atmos. Environ. 2015, 107, 95–106. [Google Scholar] [CrossRef]
- Bartlett, K.B.; Bartlett, D.S.; Harriss, R.C.; Sebacher, D.I. Methane emissions along a salt-marsh salinity gradient. Biogeochemistry 1987, 4, 183–202. [Google Scholar] [CrossRef]
- Liu, H.; Wu, X.; Li, Z.; Wang, Q.; Liu, D.; Liu, G. Responses of soil methanogens, methanotrophs, and methane fluxes to land-use conversion and fertilization in a hilly red soil region of southern China. Environ. Sci. Pollut. Res. 2017, 24, 8731–8743. [Google Scholar] [CrossRef]
- He, Y.; Guan, W.; Xue, D.; Liu, L.; Peng, C.; Liao, B.; Hu, J.; Zhu, Q.A.; Yang, Y.; Wang, X.; et al. Comparison of methane emissions among invasive and native mangrove species in Dongzhaigang, Hainan Island. Sci. Total Environ. 2019, 697, 133945. [Google Scholar] [CrossRef]
- Arai, H.; Yoshioka, R.; Hanazawa, S.; Minh, V.Q.; Tuan, V.Q.; Tinh, T.K.; Phu, T.Q.; Jha, C.S.; Rodda, S.R.; Dadhwal, V.K.; et al. Function of the methanogenic community in mangrove soils as influenced by the chemical properties of the hydrosphere. Soil Sci. Plant Nutr. 2016, 62, 150–163. [Google Scholar] [CrossRef]
- Sowers, K.R.; Baron, S.F.; Ferry, J.G. Methanosarcina acetivorans sp. nov., an acetotrophic methane-producing bacterium isolated from marine sediments. Appl. Environ. Microbial. 1984, 47, 971–978. [Google Scholar] [CrossRef]
- Lyimo, T.J.; Pol, A.; Op den Camp, H.J.; Harhangi, H.R.; Vogels, G.D. Methanosarcina semesiae sp. nov., a dimethylsulfide-utilizing methanogen from mangrove sediment. Int. J. Syst. Evol. Microbiol. 2000, 50, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Coyne, M. Soil Microbiology: An Exploratory Approach; Delmar Publishers: New York, NY, USA, 1999. [Google Scholar]
- Roslev, P.; King, G.M. Regulation of methane oxidation in a freshwater wetland by water table changes and anoxia. FEMS Microbiol. Ecol. 1996, 19, 105–115. [Google Scholar] [CrossRef]
- Le Mer, J.; Roger, P. Production, oxidation, emission and consumption of methane by soils: A review. Eur. J. Soil Biol. 2001, 37, 25–50. [Google Scholar] [CrossRef]
- Megonigal, J.P.; Schlesinger, W.H. Methane-limited methanotrophy in tidal freshwater swamps. Glob. Biogeochem. Cycles 2002, 16. [Google Scholar] [CrossRef]
- Cai, Y.; Zheng, Y.; Bodelier, P.L.E.; Conrad, R.; Jia, Z. Conventional methanotrophs are responsible for atmospheric methane oxidation in paddy soils. Nat. Commun. 2016, 7. [Google Scholar] [CrossRef]
- Pandit, P.S.; Ranade, D.R.; Dhakephalkar, P.K.; Rahalkar, M.C. A pmoA-based study reveals dominance of yet uncultured Type I methanotrophs in rhizospheres of an organically fertilized rice field in India. 3 Biotech 2016, 6, 135. [Google Scholar] [CrossRef]
- Oswald, K.; Graf, J.S.; Littmann, S.; Tienken, D.; Brand, A.; Wehrli, B.; Albertsen, M.; Daims, H.; Wagner, M.; Kuypers, M.M.M.; et al. Crenothrix are major methane consumers in stratified lakes. ISME J. 2017, 11, 2124–2140. [Google Scholar] [CrossRef]
- Shiau, Y.J.; Cai, Y.F.; Lin, Y.T.; Jia, Z.; Chiu, C.Y. Community structure of active aerobic methanotrophs in Red Mangrove (Kandelia obovata) soils under different frequency of tides. Microb. Ecol. 2017. [Google Scholar] [CrossRef]
- Shiau, Y.J.; Cai, Y.F.; Jia, Z.J.; Chen, C.L.; Chiu, C.Y. Phylogenetically distinct methanotrophs modulate methane oxidation in rice paddies across Taiwan. Soil Biol. Biochem. 2018, 124, 59–69. [Google Scholar] [CrossRef]
- He, R.; Wooller, M.J.; Pohlman, J.W.; Catranis, C.; Quensen, J.; Tiedje, J.M.; Leigh, M.B. Identification of functionally active aerobic methanotrophs in sediments from an arctic lake using stable isotope probing. Environ. Microbiol. 2012, 14, 1403–1419. [Google Scholar] [CrossRef]
- Dumont, M.G.; Luke, C.; Deng, Y.; Frenzel, P. Classification of pmoA amplicon pyrosequences using BLAST and the lowest common ancestor method in MEGAN. Front. Microbiol. 2014, 5. [Google Scholar] [CrossRef]
- Zheng, Y.; Jia, Z. The application of biomarker genes for DNA/RNA-stable isotope probing of active methanotrophs responsible for aerobic methane oxidation in six paddy soils. Acta Pedol. Sin. 2016, 53, 490–501. [Google Scholar] [CrossRef]
- Lüke, C.; Frenzel, P. Potential of pmoA amplicon pyrosequencing for methanotroph diversity studies. Appl. Environ. Microbiol. 2011, 77, 6305. [Google Scholar] [CrossRef] [PubMed]
- Ho, A.; Mo, Y.L.; Lee, H.J.; Sauheitl, L.; Jia, Z.J.; Horn, M.A. Effect of salt stress on aerobic methane oxidation and associated methanotrophs; a microcosm study of a natural community from a non-saline environment. Soil Biol. Biochem. 2018, 125, 210–214. [Google Scholar] [CrossRef]
- Bowman, J.P. Methylobacter. In Bergey’s Manual of Systematics of Archaea and Bacteria; Whitman, W.B., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015. [Google Scholar]
- Bowman, J.P. Methylomonas. In Bergey’s Manual of Systematics of Archaea and Bacteria; Whitman, W.B., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015. [Google Scholar]