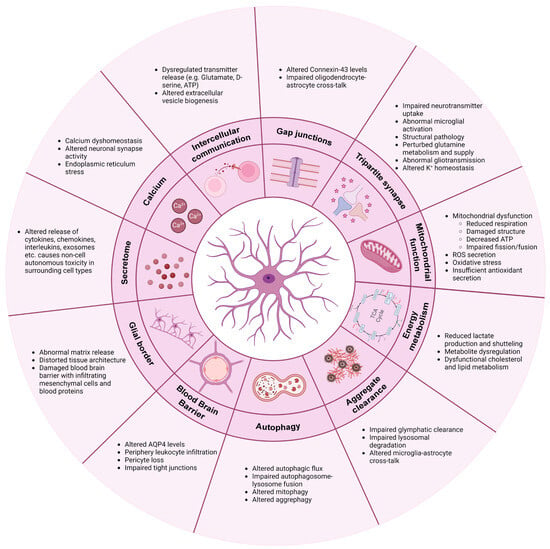
A constant stream of evidence has firmly documented that astrocytes not only contribute to neurodegenerative disease pathology and progression through non-cell autonomous mechanisms but in fact also display multiple intrinsic pathological phenotypes. In AD, PD, ALS, FTD,
multiple sclerosis (MS
) as well as prion disease, astrocytes harbor increased expression of GFAP and C3, which are thought to be important (albeit not exclusive) markers of astrocyte reactivity encompassing a neurotoxic phenotype
[1][2][3][4][5][3,37,60,78,86]. These protein expressions are part of a general observation of altered transcriptomic and proteomic levels, which largely result in a downregulation of astrocytic support functions such as ion and cholesterol homeostasis, glutamate uptake and synapse integrity and upregulation of pro-inflammatory pathways such as JAK-STAT3, NFAT and NFΚΒ with a resultant astrocytic toxic gain of functions
[6][7][8][9][10][11][12][13][14][15][16][17][18][19][38,40,67,69,70,73,87,88,89,90,91,92,93,94]. In the following section, the cell autonomous mechanisms in astrocytes, which are common across multiple neurodegenerative diseases, are highlighted (see also
Figure 1).
Figure 1.
Common mechanisms involved in astrocytic pathology in neurodegenerative diseases. Figure is created with
(accessed on 28 January 2024).
2. Communication
Unlike neurons, astrocytes are non-excitable and therefore have developed other means of communication. Besides cytokine, chemokine and interleukin releases, astrocytes use calcium waves, transmitter release and gap junction couplings as a means of communication to maintain network homeostasis
[20][49].
2.1. Calcium
Astrocytic calcium transients are highly dynamic. The “waves” are projected intra- and intercellularly through calcium-permeable ion channels and receptors, including various neurotransmitter receptors
[21][5]. Upon neurotransmitter binding, a receptor-specific intracellular calcium signal is evoked, which allows astrocytes to distinguish between, e.g., glutamatergic and cholinergic synaptic activity
[22][23][95,96]. Individual astrocytic subpopulations display unique calcium waves, which further contributes to their heterogeneity and complexity
[24][97].
In multiple neurodegenerative diseases including AD
[25][26][27][28][29][30][42,98–102], ALS
[31][32][103,104],
Huntington's disease (HD
) [33][34][35][105–107] and PD
[36][37][29,108], calcium dyshomeostasis is observed. The dysregulated calcium response is often detected early in disease progression and might therefore contribute to the pathologically altered neuronal synapse activity
[30][33][35][38][39][40][102,105,107,109–111]. For example, oligomeric forms of Aβ peptide were shown to drive astrocytic calcium hyperactivity early in AD disease progression, which consequently triggered glutamatergic hyperactivity in adjacent neurons
[38][39][109,110]. Calcium is primarily stored in the endoplasmic reticulum (ER) within astrocytes but can also be imported into the cells through AMPA and N-methyl-D-aspartate (NMDA) receptors or through voltage-gated channels
[20][49]. In ALS and PD, abnormal intracellular calcium dynamics with excess ER accumulation and storage release is observed, ultimately contributing to ER stress
[31][32][36][29,103,104]. Both genetic PD mutations and drug-induced depletion of dopamine transmission mimicking PD have shown to increase astrocytic calcium excitability
[36][37][29,108]. In the R6/2 transgenic mouse model of HD, spontaneous calcium signals and storage capacity are reduced, while evoked calcium responses are increased
[33][105].
2.2. Intercellular Communication
In addition to receiving neurotransmitter information, calcium transients can release transmitters such as glutamate, GABA, D-serine and ATP from astrocytic processes, which contribute to the regulation of neuronal synapse excitability and plasticity
[20][22][49,95]. In pathological conditions, the release of these transmitters from both neurons and astrocytes is dysregulated, and studies show that excess release causes a continuous self-activation, leading to cytotoxicity
[25][27][41][42,99,112]. This phenomenon is observed in several models of PD and AD and is triggered by excessive astrocytic calcium-dependent release of glutamate and D-serine
[39][40][42][110,111,113]. Similarly, astrocytic release of ATP can bind purinergic P2Y receptors on microglia, thereby modulating their phagocytic functions and release of cytokines further driving inflammation
[43][44][114,115]. Astrocytic self-activation is also enhanced by the release of astrocytic TNF-α and prostaglandin, which, through calcium signaling, can promote additional “gliotransmitter” release
[41][112].
The astrocyte–neuron communication can further be impaired through other means such as extracellular vesicle (EV) production. In
LRKK2-mutant astrocytes, the EV biogenesis is altered, causing an accumulation of PD-related proteins within multivesicular bodies
[45][116]. These astrocyte-secreted EVs are internalized by dopaminergic neurons and are linked to an astrocytic failure of providing neurotrophic support
[45][116]. Furthermore, C3-containing exosomes are released by AD astrocytes, consequently driving an inflammatory response and being linked to reduced neurite outgrowth
[46][47][55,117].
2.3. Gap Junctions
Astrocytes not only communicate via neurotransmitters but also through gap junction couplings. Connexin-43 (Cx43) is the predominant connexin protein, which constitutes hemichannels and gap junctions in astrocytes
[48][49][50][118,119,120]. Through these, diffusion of ions, metabolites, miRNAs and second messengers is facilitated, in addition to the important mitigation of calcium waves
[51][121]. Therefore, gap junctions are key components in astrocyte networks, contributing to synapse activity modulation and homeostatic buffering. In ALS
[9][52][53][69,122,123], AD
[25][54][42,124] and PD
[37][108], Cx43 is abnormally elevated, which causes increased gap junction coupling and hemichannel activity, resulting in calcium hyperactivity, neuronal excitability and cell death. Astrocytes also form collaborative glial networks with oligodendrocytes through their Cx43/Cx47 gap junction connections, which are important for their coordinated cross-talk
[55][125]. In MS, oligodendrocytes and astrocytes lose their communication through decreased Cx43/Cx47 gap junction expression, consequently promoting demyelination and inflammation
[56][57][58][126–128]. Lack of Cx43/Cx47 gap junction couplings is likewise found in AD
[59][129].
3. Tripartite Synapse: Neurotransmitter Regulation and Synapse Function
Astrocytes are highly involved in neuronal synapse plasticity and function through their perisynaptic process ensheathment of neuronal synapses
[60][130]. This tripartite synapse collaboration between pre- and postsynaptic neurons and astrocytes ensures optimal neuronal firing by continued astrocytic removal and recycle of excess neurotransmitters from the intersynaptic space
[61][62][131,132]. In neurodegeneration, this function is impaired. Due to the downregulation of key astrocyte receptors, excitatory amino acid transporters 1 and 2 (EAAT1 and EAAT2), astrocytes fail to properly manage the uptake of the neurotransmitter glutamate. Glutamate is the main excitatory neurotransmitter in the brain, and a large part of its uptake from the synaptic cleft appears through EAAT1/2 receptors on astrocytes
[63][64][65][133,134,135]. Persistent glutamate exposure is believed to cause excessive neuronal firing and abnormal neuronal calcium influx, which ultimately results in severe neuronal excitotoxicity
[61][64][66][67][131,134,136,137]. Lack of EAAT1/2 receptors and consequent glutamate excitotoxicity is a common phenomenon in AD
[68][69][70][71][72][138,139,140,141,142], PD
[73][74][143,144], ALS
[75][76][77][78][48,83,145,146] and HD
[33][79][80][27,28,105]. Additionally, downregulation of astrocytic glutamate receptors drives abnormal microglial pruning and phagocytosis of hippocampal glutamatergic synapses in AD
[81][147]. In HD, structural pathology, mediated by lack of astrocytic engagement of neuronal synapses, enables the hyper-excitability
[82][148]. AD astrocytes likewise have perturbed glutamine metabolism and supply, which affects the GABA synthesis
[83][84][85][86][149,150,151,152]. GABA is the main inhibitory neurotransmitter in the central nervous system, and lack thereof contributes to the excitatory imbalance
[21][5]. In addition to neurotransmitter uptake, astrocytic release of various transmitters mentioned above is additionally important for synapse activity regulation. Abnormal gliotransmission has thus been shown to affect the synaptic transmission in PD and AD, consequently contributing to synapse loss and excitotoxicity
[39][42][67][87][88][89][110,113,137,153,154,155]. Similarly, astrocytic release of C3 can bind to neuronal C3 receptors and hamper their synaptic density and dendritic morphology
[90][156].
Astrocytes control the ion homeostasis through multiple ion channels, which is crucial for maintaining synapse functionality
[91][157]. Potassium (K
+) buffering is a key mechanism affected in neurodegeneration. Under physiological conditions, astrocytes regulate K
+ levels in the extracellular space through clearance via K
+ channels such as the main astrocytic Kir4.1 sub-type
[20][49]. Through these mechanisms, astrocytes can modulate neuronal depolarization and thereby excitability
[92][158]. In HD
[93][26] and ALS
[94][159], astrocytic Kir channels are downregulated, which hampers the K
+ buffering and clearing, consequently causing increased extracellular K
+ levels, overall leading to neuronal excitotoxicity. Besides through Kir.4.1 channels, K
+ is released through EAAT2 and as mentioned previously, the astrocytic release and uptake of transmitters is modulated by calcium transients. This interconnected relationship between ion homeostasis, calcium dynamics and gliotransmission is therefore crucial for optimal neuronal function, and any dysregulation of one mechanism might consequently disrupt the others.
4. Mitochondrial Function
Mitochondrial dysfunction,
reactive oxygen species (ROS
) secretion and oxidative stress are common findings in neurodegeneration. In AD, mitochondrial dysfunctions are predicted as an early astrocytic phenotype consequently driving astrocyte reactivity
[95][23], and in PD, maintenance of mitochondrial DNA is correlated with reduced mitochondrial respiration
[36][29]. Astrocytic accumulations of α-synuclein in PD have been linked to damage within the mitochondrial structure, consequently lowering the total ATP level as well as causing disruption of the fission and fusion dynamics
[96][97][160,161]. In ALS, mitochondria display decreased membrane potential and a compromised oxygen consumption, possibly correlated with a lower secretion of antioxidants
[98][99][85,162]. Astrocytes express and release antioxidants as a part of their function in regulating the redox balance through removal of ROS in order to prevent oxidative damage of neurons
[100][163]. Astrocytic nuclear factor erythroid 2-related factor 2 (Nrf2) transcription factor is a key regulator of antioxidant, detoxification and proteostasis pathways and might therefore be an important mediator in neurodegeneration
[100][163]. In amyloid and tau pathology models of AD, pre-emptive activation of astrocytic Nrf2 was shown to be neuroprotective by attenuating the aggregation burden and slowing disease progression
[101][33]. Similar observations were seen by overexpressing Nrf2 in astrocytes in
SOD1-ALS mice
[102][164] and in α-synuclein-mutant mice in PD
[103][165], which advocates for a common therapeutic mechanism in neurodegeneration. Interestingly,
Nrf2 target gene levels are in fact increased in AD but might appear too limited or too late in the disease progression to make a considerable neuroprotective difference
[101][104][33,166].
5. Energy Metabolism
Neurons rely on astrocytes for nutritional support to meet their optimal energy consumption. Through key mechanisms such as the lactate shuttle, astrocytes convert glucose or glycogen to lactate via aerobic glycolysis and provide the lactate to adjacent neurons via monocarboxylate transporters
[105][167]. In the neurons, the lactate is incorporated into the oxidative cycle for ATP production
[105][167]. Lactate is also shuttled directly to neurons from the bloodstream via the astrocytes’ perisynaptic process engagement with the vasculature
[106][168]. In ALS, astrocytic intra- and extracellular levels of lactate are decreased, possibly due to diminished lactate production and/or transport
[107][108][169,170]. Similarly, AD astrocytes have decreased glycolysis and secretion of lactate to the environment
[29][101]. Lack of lactate production causes metabolic dysfunction and failure of adequate neuronal energy supply. Additionally, lactate production is coupled to glutamate transporter activity, and their shared impairment might therefore further hamper the energy metabolism in neurodegenerative disorders
[109][12].
A widespread dysregulation of metabolites is generally observed in neurodegenerative diseases. Astrocyte metabolic proteomics is increasingly enriched during the commencement and disease progression of AD, with markers expressing both neuroprotective and neurotoxic phenotypes
[110][24]. More specifically, AD astrocytes display augmented glycolytic flux and reduced glycogen storage
[72][142]. In PD, astrocytes have altered polyamine and phospholipid levels
[36][29], and in ALS, astrocytic metabolic dysfunction with compromised adenosine, fructose and glycogen metabolism is observed
[111][112][171,172].
Finally, astrocytes are the main synthesizers and suppliers of apolipoprotein E (ApoE), which ensures sufficient transport of cholesterol to neurons
[20][49]. Cholesterol is an essential lipid in cell membranes and presynaptic vesicle formation and is therefore crucial for the integrity and function of synapses. One of the main risk factors of late-onset AD is the human isoform
APOEε4 variant
[113][114][115][116][173,174,175,176]. Carriers are shown to have pathologically altered glucose metabolism, lactate production, cholesterol homeostasis and calcium dynamics, resulting in AD
[26][117][118][119][120][98,177,178,179,180]. Dysfunctional cholesterol and lipid metabolism are also found in HD
[121][122][123][35,181,182] and PD
[124][125][183,184] and appear to be a general mechanism of reactive astrocytes, potentially influencing many neurodegenerative diseases
[126][185].
6. Clearance of Protein Aggregates
Astrocytes have been shown to contribute to the insufficient clearance of protein aggregates in neurodegeneration due to failure of the glymphatic system. Under physiological conditions, astrocytes regulate water flux as well as removal of metabolic waste products from the brain interstitium to and from the perivascular space through their Aquaporin-4 (AQP4) water channels
[21][5]. Additionally, through the synergistic collaboration between AQP4 and Kir4.1 channels, osmohomeostasis is maintained in the brain
[127][128][129][186,187,188]. During disease progressions, astrocytes remove Aβ peptides and mutant huntingtin through the glymphatic system
[130][131][189,190]. However, due to the downregulations of Kir4.1 and AQP4, this clearing mechanism is impaired, consequently accelerating the aggregation burden
[93][132][133][26,191,192]. Additionally, astrocytic ApoE is normally involved in Aβ clearance, but in
APOEε4 carriers, this function appears to be compromised, which likewise contributes to the enhanced Aβ aggregation
[119][134][179,193].
Secondly, astrocytes contribute to clearance of proteins through their intracellular lysosomal pathways. This mechanism is affected in PD, where both α-synuclein oligomers as well as mutations in the
LRRK2 gene interfere with the astrocytic clearance of the aggregation burden
[96][135][160,194]. Neuronal α-synuclein is taken up by astrocytes from the extracellular space via endocytosis as well as transmitted directly from neurons
[18][136][93,195]. Similarly, aggregates are transferred between astrocytes via nanotubules
[97][161]. This increased inclusion burden aided by overload/stress-induced insufficient lysosomal degradation further triggers cellular toxicity and reactivity
[18][97][93,161]. More specifically, α-synuclein is shown to alter the lysosomal morphology, distribution and function by alkalinization and decrease activity of lysosomal proteases in neuronal cells and idiopathic PD brains
[137][138][196,197], which correlates with similar observations of disrupted lysosomal proteolysis recently observed in astrocytes in an early-onset PD model
[139][198]. Astrocytic engulfment of monomeric tau from the extracellular environment has been observed in tauopathies
[140][199], and the endo-lysosomal dysfunction in astrocytes is likewise predicted as an early disease phenotype in AD
[95][141][23,200].
Finally, microglial and astrocytic cross-communication is important for the optimal clearance of Aβ and α-synuclein aggregates
[142][201]. Intercellular miscommunication or toxic reactivity might therefore hamper this essential function, thereby accelerating disease progressions.
7. Autophagy
Autophagy is a vital cellular process, which facilitates the degradation and recycling of intracellular components such as damaged organelles and protein aggregates
[143][202]. Several conditions like starvation, stress and pharmacological treatment can modulate autophagy to facilitate a faster release of important nutrients in the reestablishment of cellular homeostasis
[144][203]. Autophagy is a multi-step process, where formation of large double-membrane vesicles termed autophagosomes bind and enclose cargo destined for degradation through ubiquitin-P62-LC3 protein interactions
[143][202]. The autophagosomes fuse with late endosomes and finally with lysosomes, and the lysosomal enzymes degrade the vesicular content for reuse
[145][204].
Mutations in
LRRK2 are a common cause of genetic PD
[146][205].
LRRK2 contributes, among many functions, to the phosphorylation of various proteins from the Rab family as well as P62, thereby filling a prominent role in autophagy initiation and vesicle transport
[147][206]. Consequently,
LRRK2-mutant astrocytes display impaired autophagy, possibly linked to progressive accumulation of α-synuclein as mentioned above
[148][207]. Additionally, α-synuclein oligomers have been shown to interfere with the autophagosome–lysosome fusion, consequently halting the autophagic flux in astrocytes
[97][161]. This degradation impairment results in insufficient turnover of damaged mitochondria
[97][161]. Moreover, in ALS/FTD-relevant
C9ORF72-mutant models, accumulation of P62 is present in astrocytes
[149][208], and we previously showed how human induced pluripotent stem cell-derived FTD astrocytes displayed insufficient autophagy, consequently perturbing the mitochondrial turnover
[3][60]. This lack of mitophagy resulted in augmented mitochondrial fusion, impaired mitochondrial respiration and glycolysis, increased ROS and stress granules formation, which overall triggered astrocyte reactivity and cytokine secretion
[3][60].
8. Blood–Brain Barrier (BBB)
Through close interaction with capillary endothelial cells and pericytes, astrocytes function as efficient gatekeepers of the central nervous system through regulation of the integrity and permeability of the BBB
[150][6]. Additionally, neurons rely on astrocytes for shuttling of nutrients and metabolic substrates from the blood stream through the BBB
[150][6], and with calcium-dependent release of vasodilators or vasoconstrictors, astrocytes control the blood flow depending on the neuronal energy demand
[151][209]. As mentioned previously, AQP4 levels are changed in various neurodegenerative diseases, which also contribute to the compromised BBB
[152][153][13,210]. Therefore, cerebrovascular deterioration in AD could be correlated with astrocytic calcium hyperactivity
[25][42].
In ALS, astrocytes contribute to BBB disruption and pericyte loss, which allow periphery leukocyte infiltration and a disturbance of the homeostatic environment
[154][155][156][157][158][43,211,212,213,214]. In AD,
APOEε4 carriers display pericyte degeneration due to insufficient astrocyte suppression of proinflammatory pathways, consequently damaging the BBB
[159][160][215,216]. Additionally, the barrier function of BBB tight junctions is impaired when astrocytes carry the
APOEε4 isoform
[161][217], and astrocytes modulate leukocyte infiltration due to Aβ exposure
[162][218]. In HD, astrocytes trigger endothelial cell proliferation and pericyte damage, which disrupts the vascular function
[163][219]. Finally, in PD, reactive astrocytes fail to support vessel formation and barrier integrity
[164][220].
9. Glial Border Formation
Astrocytes are crucial for the integrity of the stroma in the central nervous system and provide important structural support
[21][5]. Through their secretion of molecules such as proteoglycans, astrocytes contribute to the extracellular matrix (ECM) structure surrounding synapses and within the synaptic cleft. This is important for capturing nutrients and growth factors as well as acting as a diffusion barrier for neurotransmitter concentration buffering
[20][49]. During neurodegeneration, subpopulations of astrocytes initiate a border formation, which primarily functions to contain the injury. However, emerging evidence point towards a rising role of abnormal scarring in these disorders
[165][221]. Here, ECM molecules promote a pro-inflammatory signal mediated by microglia and infiltrating macrophages, which further drives the astrocytic reactivity and matrix release
[165][221]. Conversely, glial border formation is shown to be beneficial in its formation of “glial bridges”, along which axonal regrowth is permitted if appropriately guided by growth factors
[166][222]. Lack of astrocytic neuro-supportive functions likely influences this mechanism in neurodegenerative diseases
[167][34].
Chondroitin sulfate proteoglycans (CSPGs) are central components of glial border formation and mediated by reactive astrocytes in MS lesions
[168][223]. Although this physical barrier aids in the containment of the lesion, it also prevents axonal outgrowth and remyelination
[168][223]. As a consequence, excessive glial border formation by astrocytes might distort the tissue architecture, thereby prolonging the disease course of MS. Similarly, in ALS, CSPG accumulation is present at the site of motor neuron degeneration, and excessive and abnormal CSPG receptors are also found on the surface of reactive astrocytes, further contributing to their self-activation and the overall impairment of the homeostatic environment
[169][170][224,225]. Additionally, chronic release of TGF-β promotes excessive fibrosis in ALS
[171][226] and acts as an upstream regulator of CSPG secretion
[172][173][227,228]. In AD, astrocytic proteoglycans interact with Aβ plaques, consequently promoting aggregation and inhibiting clearance
[174][229].
Finally, the STAT3 pathway in astrocytes is an important modulator of glial border formation after spinal cord injury
[175][230]. Abnormal expression of the STAT3 pathway in multiple neurodegenerative disease could likely contribute to border formation imbalances. Similarly, the damage to the BBB generally observed in neurodegeneration permits the infiltration of mesenchymal cells and blood proteins, which mediates excessive scarring
[165][221].