1. The Gut Microbiota
The human gut microbiota, a complex ecosystem consisting of up to 100 trillion microorganisms, engages in a largely symbiotic relationship with its host
[1][25]. This diverse microbial community, collectively termed the microbiome, harbors at least 150 times more genes than the human genome itself
[1][25]. Analyses based on 16S rRNA-targeted sequencing reveal that fecal samples from healthy individuals predominantly harbor two major bacterial phyla, namely the Gram-negative Bacteroidetes consisting of the genera
Bacteroides,
Prevotella,
Parabacteroides, and
Alistipes and the Gram-positive Firmicutes made up of the
Faecalibacterium prausnitzii,
Eubacterium rectale, and
Eubacterium hallii species, in addition to numerous low-abundance species
[2][3][26,27].
Notably, the gut microbiota exhibits remarkable microbial and genetic diversity, with specific bacterial species associated with distinct gastrointestinal regions. Bacterial density steadily increases from the proximal to the distal portions of the gastrointestinal tract
[3][27]. The stomach harbors the lowest abundance, with approximately 10
1 microbial cells per gram of content. This number rises to 10
3 in the duodenum, 10
4 in the jejunum, and 10
7 in the ileum, culminating at 10
12 cells per gram in the colon
[4][28]. Consequently, the large intestine houses over 70% of the body’s microorganisms, which have a symbiotic relationship with the host and exert a substantial influence on the overall well-being of the host
[5][29].
Healthy aging comprises limiting age-related health problems in older people by avoiding or delaying chronic diseases, even though the chance of various ailments grows with age due to the loss of tissue structure and physiological function
[6][7][30,31]. This underscores the critical role that these variables play in age-related morbidity
[6][30]. Aging and health status affect biological parameters, with the human microbiota being a dynamic indication that is modified by factors such as nutrition, lifestyle, and immunological response
[8][32]. The varying composition of the gut microbiota in older adults reflects their current and previous health situations
[6][30].
The gut microbiota is essential for development and adult homeostasis, and alterations have been associated with inflammatory and metabolic problems in adults, including inflammatory bowel disease and obesity
[9][10][11][12][20,33,34,35]. While the gut microbiota remains stable and individual-specific in healthy adults, there is a significant fluctuation in older individuals compared to younger people
[13][14][36,37]. Immunosenescence, which is characterized by persistent NF-kB-mediated inflammation and the loss of naïve CD41 T cells, is one facet of the aging process
[15][38]. Intestinal homeostasis is significantly influenced by the gut microbiota, and immunosenescence is linked to the persistent activation of the innate and adaptive immune systems
[16][17][39,40].
Commensal bacteria control an innate immune response and accelerate the host’s response to enteric infections
[18][41] by increasing baseline pro-inflammatory IL-1β production, which exerts a protective effect in assisting gut pathogen clearance and neutrophil recruitment by stimulating the expression of endothelial adhesion molecules
[19][42]. Additionally, natural killer (NK) T cells are regulated by commensal bacteria that can express both T cell receptors and NK cell receptors, which help to maintain homeostasis, promote the release of inflammatory cytokines, including TNF-α, IFN-γ, IL-2, IL-4, IL-13, IL-17A, IL-21, and inhibit excessive inflammatory responses
[20][21][43,44].
Correlations between certain microbiota elements and an increase in pro-inflammatory cytokines, such as serum TNF-α, IL-6, IL-8, and C-reactive protein (CRP), have thus demonstrated this relationship in the elderly
[12][35]. Additionally, mitogens and LPSs cause macrophages to secrete more IL-6 and IL-8, which alters macrophage activity
[6][30]. Age-related dysregulation brought on by gut microbiota dysbiosis may weaken the intestinal barrier and cause the release of microbial products that raise pro-inflammatory factors like TNF-α, interferons, IL-6, and IL-1
[22][45]. This, in turn, may contribute to a chronic low-grade inflammatory state linked to chronic diseases
[23][24][25][26][18,46,47,48].
2. Obesity and Related Diseases
Obesity, defined as an abnormal or excessive accumulation of fat, has reached global pandemic proportions
[27][49]. The current clinical practice for identifying overweight and obese individuals relies primarily on body mass index (BMI)
[28][50]. Accordingly, based on established BMI classifications, values between 25 and 29.9 kg/m
2 classify individuals as overweight, while a value exceeding 30 kg/m
2 indicates obesity
[29][51]. Obesity primarily arises from an energy imbalance, where caloric intake exceeds expenditure, which leads to the storage of excess energy, such as fat and glycogen, in subcutaneous adipose tissue (SAT) and organs
[30][31][52,53]. However, adipose tissue itself exhibits functional heterogeneity, comprising distinct depots with specialized roles
[32][54].
Adipose tissue is classified into WAT and brown adipose tissue (BAT), distinguished by variations in morphology, anatomical position, developmental patterns, and metabolic functions
[33][55]. WAT serves as a key endocrine organ, storing energy in the form of triglycerides and secreting adipokines, while BAT, characterized by multilocular adipocytes and UCP-1 expression, actively contributes to energy expenditure through non-shivering thermogenesis, playing a role in regulating body temperature and providing protection against obesity
[34][56]. WAT is categorized into two primary depots, visceral WAT (VAT) and SAT, both of which are extensively examined for their correlation with the development of related diseases
[35][57]. Despite accounting for just 1% to 2% of total fat, BAT is indispensable for maintaining homeostasis and has a beneficial impact on blood glucose levels
[36][58]. Obese and diabetic individuals have smaller BATs and less activity than those with a normal BMI
[37][59].
Adipose tissue in individuals exhibiting overweight or obesity is intricately associated with a chronic, low-grade inflammatory state, characterized by the heightened infiltration of macrophages of the M1 or ‘classically activated’ phenotype from the circulation into adipose tissue, leading to adipose tissue inflammation through the release of pro-inflammatory cytokines (TNF-α, IL-6, IL-8), while the balance of anti-inflammatory cytokines (IL-4, IL-10, IL-13, IL-19) from adipocytes tends to decrease with weight gain, favoring the heightened production of pro-inflammatory adipokines
[38][39][40][60,61,62]. Adipose tissue not only releases adipokines (leptin, adiponectin, visfatin, resistin) and constituents of the extracellular matrix to modulate interconnected pathways but also undergoes hyperplasia and hypertrophy due to excess fat accumulation, altering the secretome, releasing metabolites, and subsequently influencing the surrounding microenvironment
[41][42][63,64].
An elevated level of proinflammatory adipokine leptin, in conjunction with an increase in the levels of hepatocyte growth factor (HGF), plasminogen activator inhibitor-1 (PAI-1), resistin, TNF-α, IL-1β, IL-6, and monocyte chemoattractant protein-1 (MCP-1), accompanied by a simultaneous decrease in adiponectin, contributes to the metabolic syndrome, which is characterized by glucose intolerance, insulin resistance, central obesity, dyslipidemia, hypertension, heightened cardiovascular disease risk, and increased susceptibility to cancer
[43][44][45][8,9,10]. Increased serum levels of free fatty acids (FFAs) in obese individuals promote vascular endothelial growth factor A (VEGF-A) and vimentin expression through peroxisome proliferator-activated receptor gamma (PPARγ) upregulation, contributing to tumor growth, insulin resistance, and hepatic steatosis. At the same time, the concomitant overexpression of TNF-α and leptin inhibits insulin receptor activation, inducing resistance in the muscle, liver, islet α-cells, and adipose tissue, leading to T2DM
[46][47][65,66].
3. Low-Grade Chronic Inflammation: Linking Gut Microbiota and Obesity
Obesity is characterized by changes in the abundant ratios of the dominant phyla. While some research implies that obese individuals have a higher Firmicutes to Bacteroidetes ratio, the consistency of this observation and its reliability as a biomarker remain uncertain
[48][49][50][67,68,69]. Furthermore, obesity-linked low-grade inflammatory states may be aggravated by microbiota-associated inflammatory processes
[50][69]. Seven aging pillars that form an interconnected network that converge at inflammation have been identified
[51][70], with dysfunction in one pillar leading to inflammation and subsequently impacting other pillars
[23][18]; this event is now referred to as inflammaging, defined as a “chronic, sterile (occurring in the absence of infection and primarily driven by endogenous signals), low-grade inflammation that occurs during aging”
[23][18]. This chronic inflammatory state, characterized by the innate immune system via macrophage activation and regulated by the gut microbiota, results in the production of inflammatory products
[23][52][18,71].
Previous research demonstrated that a 4-week high-fat (HF) diet resulted in a two-to-threefold increase in plasma LPS levels, which is comparable to the effects observed during the subcutaneous infusion of LPSs in mice, leading to insulin resistance and obesity
[53][72]. LPSs, a powerful activator of Toll-like receptor 4 (TLR4), are found in gram-negative bacteria
[54][73]. Hence, changes in gut microbiota composition, known as intestinal dysbiosis, may contribute to a persistent low-grade inflammatory response in obesity. Since LPSs contain lipid A, they can translocate across the intestinal mucosa via tight junctions or with chylomicron facilitation. Given that lipoproteins play a crucial role in the absorption and transport of dietary triglycerides, this mechanism may serve as an initiating factor for inflammation, potentially contributing to the commonly observed insulin resistance in obesity
[53][55][72,74].
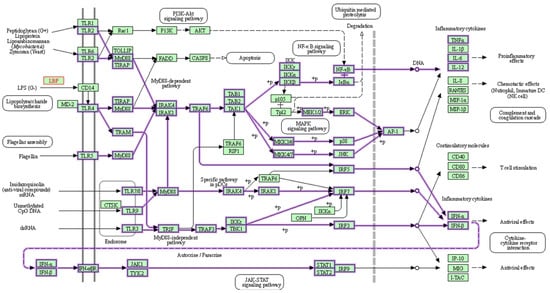
As a member of the TLR family, TLR4 is found in many different types of cells, including macrophages. It recognizes pathogen-associated molecular patterns (PAMPs) and initiates a complicated cell signaling pathway that, when bound by LPSs, activates an inflammatory response and triggers the release of cytokines provided by the KEGG pathway in
Figure 1 [56][57][58][13,75,76]. Additionally, TLR4 has been linked to the inflammatory response associated with increased intestinal permeability in circumstances such as diet-induced obesity (DIO), which leads to insulin resistance and metabolic imbalance
[56][13]. Furthermore, elevated LPS levels are associated with increased intestinal permeability, driven by the reduced expression of vital tight junction proteins like zonula occludens-1 (ZO-1), claudin, and occludin, leading to a compromised epithelial barrier that facilitates the entry of bacterial components from the intestinal lumen into the circulation, potentially initiating inflammation and insulin resistance in humans and animals
[59][60][77,78].
Figure 1. LPSs activate TLR4, signaling a chain reaction that releases inflammatory cytokines. TLRs, which are homologs of Toll in Drosophila, are present on the membranes of innate immune cells in humans (e.g., macrophages and dendritic cells) and are activated by membrane components from Gram-positive or Gram-negative bacteria. When TLRs detect pathogens, they immediately activate innate immunity, causing the generation of proinflammatory cytokines and increasing the expression of costimulatory molecules. As shown above, TLR signaling networks include a MyD88-dependent pathway that rapidly activates NF-kB and MAPK, resulting in the generation of proinflammatory cytokines, while the MyD88-independent pathway is linked to delayed NF-kB and MAPK activation, resulting in the stimulation of IFN-beta, IFN-inducible genes, and dendritic cell maturation. The green box represents organism-specific pathways; +p = phosphorylation
[58][76].
4. Short-Chain Fatty Acids (SCFAs)
The fermentation of non-digestible carbohydrates in the cecum and colon by the gut microbiota produces SCFAs, such as acetate, propionate, and butyrate, demonstrating metabolic cooperation among the bacterial community, where the collective role of the entire community is emphasized, and absorbed SCFAs in the intestine occur via passive diffusion via monocarboxylate transporter 1 (MCT1)
[61][79]. SCFA, particularly butyrate, is a primary source of energy for colonic epithelial cells, promoting cell proliferation and differentiation
[62][63][80,81], whereas acetate and propionate play separate roles in cholesterol/fatty acid precursor and gluconeogenesis, respectively
[64][82]. While other bacterial by-products, such as conjugated linoleic acids and bile acids, and gases, including methane and hydrogen sulfide, have metabolic regulatory activities, they play limited roles in mammalian physiology in comparison to SCFA’s dominant effect
[65][66][83,84].
Butyrate and acetate are essential for maintaining epithelial barrier function by influencing tight-junction protein expression (zonulin and occludin), increasing mucus production, and reducing intestinal permeability, with acetate having the most pronounced effects on epithelial protection and both SCFAs contributing to increased fatty acid oxidation and energy expenditure, potentially leading to weight loss, insulin sensitivity, and improved metabolic health
[67][68][85,86]. SCFAs block NF-kB activation in host immune cells via binding to the G-protein-coupled receptors 43 and 41 (GPR43 and GPR41), with GPR43 playing an important role in regulating the anti-inflammatory responses elicited by acetate
[69][70][87,88].