1. TEX Contribute to the Differentiation of Macrophages into Highly Immunosuppressive M2-Polarised Phenotype
Innate immunity involves various immune cells. Macrophages, in particular, constitute the majority of the inflammatory infiltrate in tumour microenvironment (TME), both in terms of abundance and functionality
[1][35]. Given the functional plasticity of macrophages, they polarise into different functional phenotypes in response to different environmental stimuli—M1 (anti-tumour)/M2 (pro-tumour)
[2][36]. In most cancers, macrophage-infiltrates within the TME, termed tumour-associated macrophages (TAMs), are skewed towards the M2 phenotype and are associated with the promotion of tumour progression and survival
[3][37]. They are known for stimulating angiogenesis, fostering an inflammatory milieu, and suppressing anti-tumour immunity in the TME via altered cytokine profiles
[4][5][38,39].
Macrophage migration inhibitory factor (MIF) was suggested to play a predominant role in M2 macrophage polarisation, and MIF depletion spontaneously reverts TAMs to an M1-like polarisation, as found in oral squamous cell carcinoma
[6][40]. Chen et al. subsequently reported that MIF was upregulated in NPC and was related to a lower survival rate
[7][41]. More importantly, they have located MIF in exosomes derived from NPC cells. These MIF-rich TEX will not only affect the polarisation of M2 macrophages through the AMP-activated protein kinase (AMPK) pathway, but also prolong the survival of tumourigenic TAMs by reducing their ferroptosis through the upregulation of glutathione peroxidase 4 (GPX4)
[7][41]. Following, another study has connected NPC-TEX-mediated macrophage polarisation to the degradation of phosphatase and tensin homolog (PTEN) in TAMs
[8][42]. PTEN instability has also been previously reported to cause bladder cancer progression
[9][43]. The ring finger protein 126 (RNF126) found in NPC-TEX was shown to promote PTEN ubiquitination and, thus, degradation, in the exosome-receiving TAMs
[8][42]. Consequently, the PI3K/AKT pathway will be activated, and autophagy will be inhibited, resulting in increased polarisation, migration, and invasion of TAMs
[8][42]. Autophagy was reported to be an important defence against M2 polarisation
[10][11][12][44,45,46]. Therefore, it is conceivable that RNF126 in NPC-TEX contributes to the immunosuppressive TME by suppressing autophagy and promoting M2 polarisation
[8][42]. In addition, long non-coding RNAs (lncRNAs), such as the tumour protein P73 antisense RNA 1 (TP73-AS1), have also been shown to take part in TEX-mediated M2 polarisation
[13][47]. Yao et al. reported that tumour protein P73 antisense RNA 1 (TP73-AS1) is upregulated in NPC and associated with a poorer prognosis
[13][47]. Their further investigation showed that TP73-AS1 delivered by TEX could lead to M2 polarisation of macrophages and alter their cellular functions. To explain the phenomenon, one possible target of this lncRNA is the tumour-suppressor miR-342-3p. TP73-AS1 has been demonstrated to have a negative correlation with miR-342-3p, possibly due to its ability to directly bind to and sponge miR-342-3p
[13][47].
Functional changes in TAMs, in terms of phagocytosis and cytokine production, can also be affected by NPC-TEX
[14][15][48,49]. A recent study on scavenger receptor class B member 1 (SCARB1) reported its enrichment in NPC-TEX, and these TEX can be taken up by macrophages to confer tumourigenicity by targeting the transcription factor krueppel-like factor 9 (KLF-9)
[14][48]. In M1 macrophages, SCARB1 upregulates the expression of 3-hydroxyanthranilate 3,4-dioxygenase (HAAO), leading to ferroptosis of the macrophages
[14][48]. This reduces the protective effects from the M1 subtypes. Likewise, in M2 TAMs, SCARB1 downregulates the CYP1B1 gene of the cytochrome P450 family, inhibiting their phagocytic capacity
[14][48]. As a result, the remnant anti-tumour effects of M2 TAMs will also be hampered. Wang et al., on the other hand, found increased IL-6 secretion in macrophages treated with NPC-TEX, which is potentially linked to tumour progression and immunosuppression
[15][49]. Chronic inflammation in the TME is known to promote tumour progression
[16][50]. The linkage between EBV infection, immunity, inflammation, and NPC is largely contributed by the cytokines produced from the activated innate immune cells. TNF-α, TGF-β, and IL-6, -10, -12, -17, are some commonly studied examples
[17][51]. IL-6, in this case, has shown capability to induce NPC growth via STAT3 activation and IL-6R overexpression in the EBV-infected epithelial cells
[18][52]. In addition, IL-6 can also lead to immunosuppression such as by reducing the abundance of CD8+ tumour-infiltrating lymphocytes (TILs) in the TME while expanding the Tregs population, as demonstrated in other cancers
[19][53].
In short, NPC-TEX alter TAMs phenotypically and functionally, resulting in an immunosuppressive TME. This highlights the significance of TEX in NPC immune evasion. Future studies may consider investigating the possible effects of TEX on other innate immune cells more in depth as well. On the other hand, the mechanisms underlying the cytokine changes in TAMs, such as that of IL-6, can also be further studied
[15][49]. As such, this will help us to map out how TEX orchestrate the inflammatory milieu in NPC. By understanding the pathogenic effects of this network, potential therapeutic targets can also be identified for controlling it.
2. Exosome-Mediated Inhibition of T Cells in NPC TME
Another typical feature of the NPC is the high abundance of TILs in its TME, with a rich CD8+ profile
[20][21][54,55]. Paradoxically, the NPC TME is also known to be immunosuppressive, with poor effector cell function, characterised by exhaustive markers
[22][56]. The presence of abnormal quantities of Tregs within the tumour site and the peripheral blood is both an explanation and a clear indication of it
[23][57]. Exosomes as insidious regulators in the TME may be implicated in this exhaustive T cell profile.
2.1. TEX-Mediated T Cell Inhibition Mechanisms Associated with EBV
Since NPC is associated with EBV infection, the interplay between EBV+ NPC cells and TILs has always been an area of focus. Exosomes derived from EBV+ tumour cells might therefore have implications for T cell exhaustion. Galectin-9 is a highly abundant protein in NPC, and is associated with exosomes
[24][58]. It is a known agonist for the T-cell immunoglobulin and mucin domain (Tim-3), which is a death-inducing receptor expressed on mature Th1 cells
[25][59]. Researchers specifically studied the effects of galectin-9 in exosomes derived from EBV+ NPC cells and validated their capability in inducing Th1 apoptosis through galectin-9/Tim-3 interaction
[26][60]. However, it should be noted that Th1 cells in the in vitro study were derived from healthy EBV carriers instead of NPC patients. Whether Th1 in the actual NPC TME would express the same level of Tim-3 and undergo apoptosis is yet to be further confirmed in future studies
[26][60]. Following, cancer-associated fibroblasts (CAFs), are also linked to exosomes
[27][61]. CAF is a population of cells involved in extracellular matrix (ECM) remodelling, and potentially, immunosuppression
[28][29][62,63]. Reported in a study, TEX containing EBV products, such as LMP-1, were found to activate the yes-associated protein (YAP1) signalling pathways in fibroblasts, leading to the formation of CAFs
[27][61]. These CAFs had an increased secretion of immunosuppressive cytokines IL6, IL8, and CCL2
[27][61]. Through 3D spheroid model studies, it was confirmed that these TEX-induced CAFs would reduce the abundance of CD8+ T lymphocytes in TME
[27][61]. While EBV was mentioned in these studies, the comparison with EBV- NPC cells-derived TEX was inadequate, and the evidence may not be conclusive. A panoramic understanding of the role of EBV in TEX-mediated immune suppression is yet to be drawn.
2.2. miRNAs in NPC-TEX Alters the Differentiation Patterns and Secretome of T Cells
The suppression of NPC-TEX on the activation and proliferation of both CD4+ and CD8+ T cells has also been attributed to several miRNAs—“miR-24-3p, miR-20a-5p, miR-891a, miR-106a-5p, and miR-1908”
[30]. miR-24-3p, in further investigations, was found to target fibroblast growth factor 11 (FGF11)
[31][64]. miR-20a, denoted by another study, also affects STAT3 signalling
[32][65]. FGF11 was recognised as the central regulator in the mitogen-activated protein kinase (MAPK) pathway
[31][64]. The suppression of FGF11 by miR-24-3p, alongside the effects of other miRNAs in the TEX, disrupted the MAPK signalling in T cells. This resulted in altered phosphorylation of ERK and STAT proteins in the downstream
[31][64]. Consequently, the differentiation of CD4+ T cells into Th1 and Th17 cells was inhibited, while their differentiation into Tregs was promoted
[30]. This might, in turn, hamper the activity of CD8+ T cells. IFN-γ, as indicated by inflammatory studies, is not only an important activator of effector T cell, but also an antagonist to Tregs’ proliferation and function. The depletion of IFN-γ-producing Th1 cells could explain the diminished CD8+ effector functions and the expansion of Tregs within the TIME of NPC
[33][66]. Th1 differentiation itself has also demonstrated inhibitory effects on Tregs’ generation in specific contexts
[34][67]. The function is, however, likely compromised in NPC. Subsequent to phenotypical changes, the cytokine profile of T cells would also be affected. CD4+ T cells were shown to produce more IL-6 but less IL-2, IL-12, IL-17, and IFN-γ
[30]. Likewise, CD8+ T cells also showed increased production of IL-6 and IL-10 but reduced TNFα
[30]. The roles of IL-6 in tumour progression and immunosuppression have been discussed
[18][19][52,53], and the immunostimulatory roles of cytokines like IFN-γ, IL-2, and TNFα in cancer biology are also known
[35][36][68,69]. NPC-TEX, in short, increases immunosuppressive cytokines and reduces immunostimulatory cytokines within the TME, further contributing to immune evasion.
2.3. NPC-TEX Directly Recruit Tregs to Promote Immune Evasion
The expansion of Tregs in the NPC TME could not only be secondary to the phenotypical and secretome changes in T cells, but also occur under the direct effects of TEX. Mrizak et al. unveiled that NPC-TEX could directly recruit Tregs into the TME and promote their proliferation via CC motif chemokine ligand and receptor (CCL20-CCR6) interaction
[37][33]. In further investigations, NPC-TEX were also found to recruit conventional CD4+ CD25- T cells into the TME and promote their differentiation into CD4+ CD25
high Tregs
[37][33]. These TEX-induced Tregs presented with an altered cytokine profile in which they tend to secrete more IL-10 and transforming growth factor beta 1 (TGFB1), further contributing to an immunosuppressive TME
[37][33].
2.4. NPC-TEX Surface Proteins Contribute to T Cell Exhaustion
The latest studies on NPC-TEX tend to focus more on improving tumour sensitivity to immune checkpoint inhibitors (ICI), which is highly relevant to T cells
[38][70]. It is known that tumour cells and their exosomes express surface PD-L1 (Exo-PD-L1), contributing to immune evasion
[39][71]. Yang et al. validated in NPC that Exo-PD-L1 could substantially inhibit the proliferation, cytokine production, and tumour infiltration of CD8+ cytotoxic T lymphocytes (CTLs)
[40][31]. Such immunosuppression could be reversed by anti-PD-L1 antibodies, or exosome secretion inhibitors, presenting potential avenues in immunotherapy in addition to the current use of anti-PD-1 antibodies such as pembrolizumab and nivolumab
[41][72]. Moreover, a recent study on melanoma discovered that the loading of PD-L1 into TEX is mediated by the phosphorylation of hepatocyte-growth-factor-regulated tyrosine kinase substrate (HRS), a key component of the machinery which regulates the sorting of cargos into exosomes, suggesting that specifically inhibiting the phosphorylation of HRS in tumour cells could enhance the efficacy of ICI treatments
[42][73].
2.5. Potential Pathways of TEX-Mediated T Cell Inhibition Yet to Be Found in NPC
Exosome-mediated suppression of T cells can also take place via indirect routes. CTLs’ activation and proliferation depend largely on antigen-presenting cells (APC) MHC I. Therefore, alterations in DCs’ maturation and antigen presentation are plausible causes of CTLs inhibition. Early in 2013, researchers pointed out that TEX can block the differentiation of myeloid precursors and dendritic cell precursors, resulting in a reduction in DCs and expansion of MDSCs
[43][74]. Likewise, S100A9 in melanoma TEX reduced the expression of CD83, CD86, IL-12, and IL-15 in DCs, inhibiting their maturation
[44][75]. Moreover, TEX galectin-9 in glioblastoma inhibited the maturation of DCs and prevented antigen presentation, hampering the activation of CTLs in the cerebrospinal fluid
[45][76]. TEX in a prostate cancer model also exhibited a predominant role in inducing a non-native CD73+ phenotype in DCs, promoting an adenosine-mediated suppression of CD8+ T cell activation via impaired DC antigen presentation
[46][32]. Similar exosome-mediated mechanisms that indirectly result in the inhibition of T cells or other immune cells are, however, poorly understood in NPC. Given the central role of CTLs in immunotherapy, it is suggested that related pathways should be uncovered.
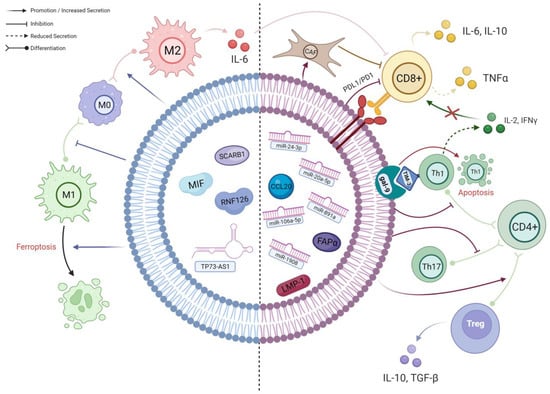
To summarise, TEX impede the anti-tumour response of T cells through numerous mechanisms, which can potentially diminish the efficacy of immunotherapies. By counteracting TEX, we could potentially enhance the success of immunotherapy in treating NPC (Figure 12).
Figure 12. A schematic diagram of the known interactions between TEX and different immune cells in NPC. The left side of the TEX (BLUE) shows cargos which predominantly affect macrophages; their effects are mainly on macrophage polarisation and viability. The right side of the TEX (RED) demonstrates the cargos involved in the complex interplay between different subpopulations of T cells. As delineated, CD4+ T cells preferentially differentiate into immunosuppressive Tregs under the influence of TEX, resulting in an altered cytokine secretion profile. Along with the effects of other TEX cargos, the infiltration and activation of CD8+ T cells will ultimately be hampered, leading to further immune evasion (created with BioRender.com, accessed on 22 October 2023).
3. Involvement of Exosomes in the Disease Progression and Metastasis of NPC
Tumour progression is the summed outcome of numerous processes. Apart from immune evasion, events such as angiogenesis, epithelial-to-mesenchymal transition (EMT), therapeutic resistance, etc., also play indispensable roles
[47][77]. Through these processes, the cancer becomes more aggressive in clinical behaviour over time
[48][78], resulting in increased tumour growth, invasiveness, and ultimately metastasis
[49][79]. Accumulating evidence shows that TEX might be involved in facilitating these processes. Here, the researchers aim to provide several examples to further highlight the importance of exosome studies in NPC (
Table 1).
Table 1.
TEX-mediated pathways leading to tumour progression and metastasis in NPC.
| Pathogenic Mechanism |
Exosome Cargo(s) |
Target Gene(s)/Pathway(s) |
Effects of Exosome Cargo(s) in Recipient Cells |
Ref. |
| 1a. Angiogenesis |
VEGF Upregulation in
ECs |
ICAM-1 |
MAPK |
Mediates neovascularisation through Src kinase, ERK1/2, p38 MAPK, RhoA/ROCK and eNOS. |
[50][80] |
| miR-17-5p |
BAMBI |
miR-17-5p downregulate BAMBI, leading to disinhibition of Akt, and thus increased downstream VEGF-A expression. |
[51][81] |
| miR-144 |
FBXW7/HIF-1 α Axis |
miR-144 upregulates VEGF-A via the FBXW7/HIF-1α axis. |
[52][82] |
| Other Mechanisms to Alter ECs’ Properties |
STIM1/LMP-1 |
Akt/ERK |
EBV LMP-1 promotes proliferation, migration, tubulogenesis and permeability in ECs by activating the Akt/ERK pathway. STIM1 was found to promote LMP-1 enrichment in NPC-TEX. |
[53][83] |
| miR-23a |
TSGA-10 |
miR-23a represses TSGA10 and positively regulates ERK signalling, resulting in proliferation, migration, and formation of tube-like structures in ECs. |
[54][84] |
| miR-205-5p |
DSC2 |
miR-205-5p downregulates tumour suppressor DSC2 to promote EGFR/ERK signalling and enhance ECs’ proliferation. |
[55][85] |
| CCAT2 |
Unknown |
The lncRNA boosts the proliferation and migration ability of ECs. |
[56][86] |
| lincROR |
p-AKT, p-VEGFR2 |
Accelerate the growth of blood vessels, contributing to proliferation, migration and tube formation ability. |
[57][87] |
| PFKFB3 |
ERK, p-AKT |
PFKFB3 improve vessel sprouting by regulating cytoskeleton remodelling, migration and tip cell competitiveness. |
[58][88] |
| HAX-1 |
ITGB6 |
By upregulating ITGB6, the FAK pathway is activated, leading to higher cell junction permeability and neovascularisation. |
[59][89] |
| miR-455 |
ZO-1 |
Under hypoxia, the exosomal miR-455 level is increased, which increases vascular permeability via the inhibition of ZO-1, a protein for endothelial tight junctions. |
[60][90] |
| ECM Modulation |
CD44v5 |
Adhesive Proteins |
Mediates endothelial cell adhesive proteins and their interactions with ECM components. |
[50][80] |
| EBERs |
VCAM-1 |
EBERs delivered into ECs are recognised by cytoplasmic TLR-3/RIG-I, activating the downstream ERK1/2/AP1 axis, thus stimulating VCAM-1 adhesive protein expression. |
[61][91] |
| miR-205-5p |
DSC2 |
Inhibition of DSC2 by miR-205-5p enhances EGFR/ERK signalling and MMP-2, -9 expression such that extracellular matrix proteins are degraded and remodelled. |
[55][85] |
| 1b. Metastasis |
| Promoting Tumour Intravasation |
HMGA2 |
Snail |
HMGA2 is overexpressed in EBV-infected NPC and their TEX. It upregulates Snail in ECs, promoting mesenchymal transition and degrading tight junctions, increasing vascular permeability. |
[62][92] |
| Inducing EMT in NPC Cells |
MMP-13 |
AKT1,
ERK1/2 |
NPC-TEX is often enriched in MMP-13. Overexpression of HIF-1α in NPC cells under hypoxia is a probable cause. Exosomal MMP-13 could induce EMT in normoxic tumour cells possibly through AKT1 and ERK1/2 signalling. |
[63][93] |
| miR-106a-5p |
FBXW7- TRIM24- SRGN Axis |
Exosomal miR-106a 5p suppresses FBXW7 to downregulate FBXW7-mediated ubiquitin degradation of TRIM24. Thus, more TRIM24 binds to SRGN promoter region, and SRGN then promotes metastasis through EMT. |
[64][94] |
| miR-18a-5p |
BTG3 |
EMT markers are induced by TEX miR-18a-5p in NPC cells by suppressing BTG3 and activating the Wnt/β-catenin pathway. |
[65][95] |
| miR-301a-3p |
BTG1 |
Aberrant expression of miR-301a-3p promotes the proliferation, migration, invasion and EMT of NPC cells by suppressing BTG1 mRNA, a tumour suppressor gene. |
[66][96] |