Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Nana-Maria Grüning and Version 3 by Catherine Yang.
Oxidative stress, characterized by an imbalance between the production of reactive oxygen species (ROS) and the cellular anti-oxidant defense mechanisms, plays a critical role in the pathogenesis of various human diseases. ROS are either produced through cellular processes or environmental factors. Of note, oxidative stress has been described as a secondary effect within the pathology of several rare monogenic diseases and sometimes been called a common denominator.
- oxidative stress
- reactive oxygen species (ROS)
- cellular redox balance
1. The Mitochondrial Respiratory Chain
The mitochondrial respiratory chain, or electron transport chain (ETC), is one of the biggest endogenous sources of ROS in eukaryotic cells which use aerobic metabolism (Figure 1) [1][2][18,19]. In humans, the enzyme complexes I (NADH:ubiquinone oxidoreductase), II (succinate:ubiquinone oxidoreductase), and III (ubiquinol:cytochrome c oxidoreductase) are somewhat “leaky”, leading to direct one-electron transfer to molecular oxygen and the perpetual production of the superoxide anion radical (O2·−) as side reactions [2][3][19,20].
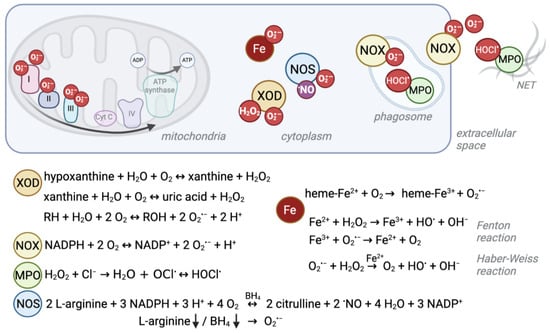
Figure 1. Schematic overview of ROS production sites and reactions. Several enzymatic reactions as well as interactions of iron with oxygen give rise to reactive oxygen species. Their localization and production level can differ between cell compartments and cell types. For example, red blood cells experience high levels of superoxide due to their heme-bound iron (Fe2+) and oxygen interaction, whereas the mitochondrial electron transport chain is a major source of ROS in other cells. Under ischemic conditions, xanthine oxidase (XOD) produces superoxide anions (O2·−) and hydrogen peroxide (H2O2) in the cytosol. NADPH oxidases (NOX) mainly produce O2·− to kill pathogens in the phagosome or extracellular space. Similarly, myeloperoxidase (MPO) produces hypochlorous acid (HOCl.) from H2O2 which derives from NOX-produced O2·− in phagosomes and neutrophil extracellular traps (NETs). Instead, nitric oxide synthases (NOS) normally produce nitric oxide (.NO) as a signaling molecule. Depletion of its substrate arginine or cofactor (6R)-5,6,7,8-tetrahydro-L-biopterin (BH4) can cause the enzyme to uncouple and produce O2·−.
Complex I is encoded by more than 40 genes [4][21], complex II by four genes [5][22] and complex III by 11 genes [6][23]. However, the specific sites and levels of ROS production within the different protein complexes depend on the respiration substrate, whether the cell experiences norm- or hypoxic conditions, and on the inhibitor used in experimental setups [7][24]. Furthermore, respiratory complexes can assemble into supercomplexes, which decreases ROS production [8][25]. Astrocytes contain high amounts of free complex I and thus potentially higher levels of ROS compared to neurons, which display complex I and III assembly [9][26]. Thus, the diversity of conditions as well as of cell type specificities make it often hard to pin down the specific set of genes of respiratory complex subunits which involve pathomechanisms related to ROS production in humans. This is also exemplified by the fact that some pathogenic mutations within subunits do not lead to ROS elevation, as shown for H2O2 in a mouse model for Leigh syndrome [10][27]. By contrast, in other experiments, mutations of genes involved in complex I, II, or III formation were indeed shown to create greater “leakiness”, and cause elevated ROS levels and oxidative stress [4][11][12][13][21,28,29,30].
Since the ETC generates a strong proton gradient used for oxidative phosphorylation, transport processes, and heat production, it can be difficult to disentangle whether the pathophysiological mechanism triggered by mutation of ETC genes is based on compromised primary functions, or elevated ROS levels. However, at least for complex I deficiency, elevated ROS production and its consequent oxidative damage were shown to induce apoptotic molecular pathways leading to neuron de.generation and hence neurological symptoms, the main features of the disorder [11][28].
Deficiencies in respiratory complexes have similar but wide-ranging symptoms from neonatal death, lactic acidosis, myopathy, hepatopathy, encephalopathy, Leigh syndrome [14][31], Leber hereditary optic neuropathy (LHON) [15][32], to adult-onset neurological symptoms such as some forms of Parkinson disease [16][17][33,34]. Isolated complex I deficiency is the most prevalent genetic disorder of oxidative phosphorylation [18][35].
As outlined above, it is challenging to disentangle the underlying factors, but the numerous potential ROS production sites and levels might be part of the explanation of the broad range of symptoms and disorder severities.
2. Heme in Red Blood Cells
Another ROS formation hotspot, despite being devoid of mitochondria, is the cytosol of red blood cells (RBCs). RBCs transport oxygen from the lung to peripheral tissues and superoxide can be formed when O2 interacts with the iron (Fe2+) of the heme group of hemoglobin (Hb) [19][36]. Spontaneous autoxidation results in O2·− and methemoglobin (Hb-Fe3+, MetHb, Hb M) (Figure 1) [18][20][35,37], and ca. 1% of all Hb is present as Hb M in healthy individuals [21][38].
This normally occurring rate of superoxide production can be exceeded by pathogenic genetic variants that cause conformational changes to the globin chain proteins that contain heme as a prosthetic group. This phenomenon is well described for, e.g., autosomal recessive sickle cell disease (SCD) (Table 1). SCD is the most common severe hemoglobinopathy worldwide and is caused by a missense pathogenic variant in the globin beta-chain (HBB), resulting in Hb S (NM_000518.5(HBB):c.20A>T (p.Glu7Val); E6V) [22][39], which is unstable, prone to polymerization and autoxidation [23][40]. Besides other ROS sources like iron release, this autoxidation is the primary source for oxidative stress in sickle cells, and it leads to loss of membrane structure and function and consequent multisystem disease [24][25][26][27][41,42,43,44]. When iron is released, it impacts the oxidative balance through transfer of single electrons via the Fenton reaction (Fe2+ + H2O2 → Fe3+ + HO· + OH−) and the Haber–Weiss reaction (O2·− + H2O2 → O2 + HO· + OH− catalyzed by iron) (Figure 1) [28][45].
Table 1. Summary of discussed genes and phenotypes. AR, autosomal recessive, AD, autosomal dominant, XLR, X-linked recessive, OMIM®, Online Mendelian Inheritance in Man database.
| Protein | Gene (OMIM® no.) | Monogenic Disease (OMIM®no.) | Inheritance | Described by |
|---|---|---|---|---|
| ROS production | ||||
| Hemoglobin beta-locus | HBB (141900) | Sickle cell disease (603903) | AR | [22][39] |
| Methemoglobinemia, beta type (617971) | AD | [21][38] | ||
| Cytochrome b(-245), beta subunit, p91-phox | CYBB (300481) | Chronic granulomatous disease (306400) | XLR | [29][53] |
| Immunodeficiency 34, mycobacteriosis (300645) | XLR | [30][54] | ||
| Cytochrome b(-245), alpha subunit, p22-phox | CYBA (608508) | Chronic granulomatous disease 4 (233690) | AR | [31][55] |
| Neutrophil cytosolic factor-1, p47-phox | NCF1 (608512) | Chronic granulomatous disease 1 (233700) | AR | [32][56] |
| Neutrophil cytosolic factor-2, p67-phox | NCF2 (608515) | Chronic granulomatous disease 2 (233710) | AR | [33][57] |
| Neutrophil cytosolic factor-4, p40-phox | NCF4 (601488) | Chronic granulomatous disease 3 (613960) | AR | [34][58] |
| Myeloperoxidase | MPO (606989) | Myeloperoxidase deficiency (254600) | AR | [35][70] |
| Xanthine dehydrogenase | XDH (607633) | Xanthinuria, type I (278300) | AR | [36][75] |
| Nitric oxide synthase 1 | NOS1 (163731) | - | ||
| Nitric oxide synthase 2 | NOS2 (163730) | - | ||
| Nitric oxide synthase 3 | NOS3 (163729) | - | ||
| enzymatic ROS clearance | ||||
| superoxide dismutase 1 | SOD1 (147450) | Amyotrophic lateral sclerosis 1 (105400) | AD, AR | [37][93] |
| Spastic tetraplegia and axial hypotonia, progressive (618598) | AR | [38][99] | ||
| superoxide dismutase 2 | SOD2 (147460) | - | ||
| superoxide dismutase 3 | SOD3 (185490) | - | ||
| catalase | CAT (115500) | Acatalasemia (614097) | AR | [39][102] |
| glutathione peroxidase 1 | GPX1 (138320) | Hemolytic anemia due to glutathione peroxidase deficiency (614164) | AR | [40][108] |
| glutathione peroxidase 4 | GPX4 (138322) | Spondylometaphyseal dysplasia, Sedaghatian type (250220) | AR | [41][110] |
| glutathione peroxidase 2, 3, 5-8 | GPX2 (138319) GPX3 (138321) GPX5 (603435) GPX6 (607913) GPX7 (615784) GPX8 (617172) | - | ||
| peroxiredoxin 3 | PRDX3 (604769) | Corneal dystrophy, punctiform, and polychromatic pre-Descemet (619871) | AD | [42][120] |
| Spinocerebellar ataxia, autosomal recessive 32 (619862) | AR | [43][116] | ||
| peroxiredoxin 1, 2, 4, 5, 6 | PRDX1 (176763) PRDX2 (600538) PRDX4 (300927) PRDX5 (606583) PRDX6 (602316) | - | ||
| thiols and thiol production | ||||
| gamma-glutamylcysteine synthetase, catalytic subunit | GCLC (606857) | Hemolytic anemia due to gamma-glutamylcysteine synthetase deficiency (230450) | AR | [44][124] |
| glutathione synthetase | GSS (601002) | Glutathione synthetase deficiency (266130) | AR | [45][127] |
| Hemolytic anemia due to glutathione synthetase deficiency (231900) | AR | [46][126] | ||
| thioredoxin 2 | TXN2 (609063) | Combined oxidative phosphorylation deficiency 29 (616811) | AR | [47][135] |
| thioredoxin | TXN (187700) | - | ||
| thiol recycling | ||||
| glutathione reductase | GSR (138300) | Hemolytic anemia due to glutathione reductase deficiency (618660) | AR | [48][138] |
| thioredoxin reductase 1, 2, 3 | TXNRD1 (601112) TXNRD2 (606448) TXNRD3 (606235) | - | ||
| NADPH production | ||||
| glucose 6P-dehydrogenase | G6PD (305900) | Hemolytic anemia, G6PD deficient (favism) (611162) | XL | [49][147] |
| 6-phosphogluconate dehydrogenase | PGD (172200) | Phosphogluconate dehydrogenase deficiency (619199) | AD | [50][151] |
| malic enzyme 1 | ME1 (154250) | - | ||
| malic enzyme 3 | ME3 (604626) | - | ||
| glutamate dehydrogenase 1 | GLUD1 (138130) | Hyperinsulinism-hyperammonemia syndrome (606762) | AD | [51][156] |
| glutamate dehydrogenase 2 | GLUD2 (300144) | - | ||
| isocitrate dehydrogenase 2 | IDH2 (147650) | D-2-hydroxyglutaric aciduria 2 (613657) | AD | [52][158] |
| isocitrate dehydrogenase 1 | IDH1 (147700) | - | ||
| methylenetetrahydrofolate dehydrogenase 1 | MTHFD1 (172460) | Combined immunodeficiency and megaloblastic anemia with or without hyperhomocysteinemia (617780) | AR | [53][166] |
| methylenetetrahydrofolate dehydrogenase 2 | MTHFD2 (604887) | - | ||
| 10-formyltetrahydrofolate dehydrogenase (mitochondrial) | ALDH1L2 (613584) | neuro-ichthyotic syndrome? | AR | [54][172] |
| 10-formyltetrahydrofolate dehydrogenase (cytosolic) | ALDH1L1 (600249) | - | ||
Also, other HBB variants have been described to cause methemoglobinemia. For example, the so-called Hb M-Hyde Park (Hb M-Akita) (NM_000518.4(HBB):c.277C>T (p.His93Tyr)) variant leads to conformational changes at the heme binding site and hence higher rates of methemoglobin formation. Inheritance is autosomal dominant [21][55][38,46].
3. NADPH Oxidases and Myeloperoxidase
Besides these cellular processes in which ROS are formed passively as byproducts, the cell can actively produce high levels of ROS through dedicated enzymes. NADPH oxidases (NOX) use reduced nicotinamide adenine dinucleotide phosphate (NADPH) to generate superoxide (Figure 1) [56][47]. NOX play an important role in the innate host defense. For the so-called “respiratory burst”, superoxide is released into the extracellular space or phagosomes to fight off pathogenic bacteria or fungi [57][48]. Furthermore, NOX enzymes were found to produce intracellular ROS at lower levels, which is believed to serve signaling functions [58][49] and to control the cellular redox balance by oxidizing NADPH and through ROS formation [59][50]. The importance of the ability to generate high ROS levels is exemplified by the detrimental consequences of NOX disruption.
Several monogenic disorders are related to NOX subunits. The genes that encode the NOX2 complex in phagocytes are related to chronic granulomatous disease [60][61][51,52]: CYBB pathogenic variants cause X-linked recessive chronic granulomatous disease (CGDX, [29][53]) and immunodeficiency 34 [30][54]), CYBA pathogenic variants cause autosomal recessive (AR) CGD4 [31][55], NCF1 pathogenic variants cause AR CGD1 [32][56], NCF2 pathogenic variants cause AR CGD2 [33][57], and NCF4 pathogenic variants cause AR CGD3 [34][58] (Table 1). These rare primary immunodeficiencies increase susceptibility to life-threatening bacterial and fungal infections and lead to development of granulomas [62][59].
Pathogenic variants in other NADPH oxidases were shown to increase susceptibility to inflammatory bowel disease (NOX1 and DUOX2, [63][60]), or cause congenital hypothyroidism (DUOX2, Thyroid dyshormonogenesis 6, AR, [64][61]; DUOXA2, Thyroid dyshormonogenesis 5, AR, [65][62], Table 1). The latter one is the result of disrupted H2O2 production through mutated DUOX2, which would be required for organification of iodide for thyroid hormone synthesis catalyzed by thyroid peroxidase [61][66][52,63].
Another enzyme of the innate immune response is the heme-containing enzyme myeloperoxidase (MPO, Table 1, Figure 1). It is highly abundant in azurophilic granules of neutrophils [67][64] and is a crucial component in neutrophil extracellular nets [68][65]; it is also found—to a lesser extent—in monocytes [69][66]. To kill pathogens, MPO creates strongly reactive species, such as hypochlorous acid (HOCl.) from hydrogen peroxide (H2O2) and chloride (Cl−) [70][71][67,68]. Autosomal recessive MPO deficiency is the most common phagocyte defect but is asymptomatic in the majority of patients, suggesting compensatory mechanisms [72][69]. However, higher susceptibility to fungal infections, especially Candida albicans, has been described [35][70]. This vulnerability might become apparent only in combination with comorbidities like diabetes, which itself increases the risk of infections [71][68]. Similarly, a recent study described a case of partial DiGeorge syndrome together with MPO deficiency. DiGeorge syndrome is characterized by immunodeficiency among other symptoms. However, the patient had more frequent and severe infections than expected for partial DiGeorge alone, which the authors explained by co-occurrence of MPO deficiency [73][71]. On the other hand, a protective mechanism against cardiovascular disease by absence of the potentially oxidative stress causing MPO enzyme has also been discussed [72][69].
4. Other ROS Sources
In addition to the above-described reactions and enzymes, there are additional sources that can produce significant amounts of reactive species under certain physiological circumstances.
For example, xanthine oxidoreductase (XDH/XOD), encoded by the XDH gene (Table 1, Figure 1), exists in two interconvertible isoforms. Both forms utilize hypoxanthine or xanthine to produce uric acid. However, the predominant form has xanthine dehydrogenase (XDH) activity, and uses NAD+ as cofactor to produce NADH, whereas the xanthine oxidase (XOD) form uses oxygen to produce the superoxide anion and H2O2 [74][72]. XDH can be converted to XOD by irreversible limited proteolysis or reversibly by thiol oxidation (reviewed in [75][73]), e.g., in hypoxic/ischemic tissue [76][74]. Homozygous or compound heterozygous pathogenic XDH variants cause type I xanthinuria (XAN1) [36][75]. The symptoms, low serum and urine uric acid and xanthinuria leading to urolithiasis [77][76], result from XDH’s primary function.
Furthermore, nitric oxide synthases (NOS1-3) can become sources of O2·− (Figure 1). Normally, they use L-arginine to produce nitric oxide (·NO), which belongs to the group of reactive nitrogen species (RNS) and serves as an important signaling molecule, especially for the vascular tone [78][77]. However, persistent oxidative stress, and thus reduced levels of the cofactor (6R)-5,6,7,8-tetrahydro-L-biopterin (BH4), lead to uncoupling of endothelial NOS (eNOS, NOS3) so that the enzyme produces O2·− instead of ·NO [79][78]. Furthermore, O2·− is produced by NOS2 in arginine-depleted macrophages [80][79]. Although some NOS3 variants have been described to increase the risk for certain conditions like pregnancy- induced hypertension [81][80] or ischemic stroke [82][81], no clear monogenic disorder has been described for NOS1, NOS2, or NOS3.
At lower amounts, ROS can also be byproducts of other enzymes like cytochrome P450 or cyclooxygenase (reviewed in [83][82]).
Environmental factors that create elevated ROS levels are, e.g., UV radiation, ionizing radiation, smoking and air pollution, chemicals such as drugs, and certain types of food like the fava bean [84][85][2,83]. Often, genetic defects in the anti-oxidative machinery remain unnoticed until such environmental stresses hit and overwhelm the residual anti-oxidative capacity of the cell, e.g., in hemolytic anemia due to glucose 6-phosphate dehydrogenase (G6PDH) deficiency (see below).