Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 3 by Sirius Huang and Version 2 by Sirius Huang.
Many mechanisms of cancer progression have been explained by principles of ecology, developmental biology, and evolutionary paradigms. Many authors have discussed ecological, developmental, and evolutionary strategies for more successful anti-cancer therapies, or for understanding the ecological, developmental, and evolutionary bases of breast cancer (BC) exploitable vulnerabilities. Researchers use the integrated framework of three well known ecological theories: the Bronfenbrenner’s theory of human development, the Vannote’s River Continuum Concept (RCC), and the Ecological Evolutionary Developmental Biology (Eco-Evo-Devo) theory, to explain and understand several eco-evo-devo-based principles that govern BC progression.
- breast cancer (BC)
- onco-breastomics
- eco-evo-devo theories
- tumorigenesis
- progression
1. BC Is a Hyphenated Eco-Evo-Devo Disease
It is now known that the eco-evolutionary theories are essential for understanding cancer development at species level, as long as a species evolves by mutation and selection that acts into a population akin to a tumor that develops by somatic mutations and clonal selection of cells within a tissue [1]. If we consider the evolutionary genomics focused on genetic variation between individuals and among populations as an essential determinant of the evolutionary features of a species [2], the somatic evolutionary genomics is focused on genetic mosaicism that occurs during an organism’s development, providing new opportunities to study the origin and progression of cancer [3]. Recently, the evolutionary proteomics integrates more and more the analysis of proteins and their functions in evolutionary biology [4]. In this context, parallels between cancer and ecological systems have been studied and reviewed by many researchers [5][6][7][8][9][10]. Furthermore, Boddy (2022) suggested that evolution and ecology may unify cancer research that requires more than a cellular or molecular perspective [10].
In nature, ecosystems are thermodynamic systems, open to energy and matter that self-organize or self-regulate towards higher complexity and organization, create order, and self-maintain far from thermodynamic equilibrium [11]. Considering the structure and behavior of cancer cells, a tumor may also be viewed as an open and non-linear dynamic system in the non-equilibrium thermodynamic state, self-organizing in time and space, exhibiting high complexity, robustness, and adaptability [12]. In this context, the eco-oncology is focused on tumors as complex, adaptive, and evolving systems, the eco-evolutionary approach helping to understand cancer dynamics, biology, and management [5].
Many mechanisms of cancer progression may be explained by the principles of ecology [13], developmental biology [14][15], and evolutionary paradigms [16][17]. Many authors have discussed the ecological, developmental, or evolutionary strategies for more successful anti-cancer therapies [7], or to understand the ecological and evolutionary bases of cancer vulnerabilities [17]. In this context, cancers can be viewed as pseudo-organs [8][18], as “living organisms” able to build “self-sustainable tumor ecosystems” [7], as well as both genetically and phenotypically heterogeneous populations within individuals and different species between individuals [17], or like local ecological communities that possess limited number of species, according to local conditions [19]. Cancer is considered a by-product of multicellularity, so that many authors have emphasized that cancer progression is a complex eco-evolutionary process, suggesting that understanding cancer’s evolutionary history may effectively help to manage and treat cancer [20].
Tot (2005–2008) launched the theory of “the sick breast lobe” [21][22][23], integrating genomic, embryologic, and ecological factors that drive the BC development. According to this hypothesis, the majority of breast malignant tumors are considered as “lobar diseases”, because the BC development, as a “long-life” process, arises into a single sick breast lobe that emphasizes a certain degree of genetic instability acquired through mutations during embryonic development, followed by successive accumulation of genetic and epigenetic changes that occur during postnatal lifespan due to exposure to environmental xenobiotics/noxes [24]. Moreover, Tan and Tot (2018) showed that both the ductal carcinoma in situ (DCIS), as well as the lobular carcinoma in situ (LCIS), follow the catchment patterns of breast ductal theories [24]. Thus, BC development as a lobar disease could be compared with the integrated catchment-lake ecological models, which postulate that a natural headwater catchment is the fundamental unit that connect the land to the ocean [25], taking into account that altered patterns into a lake catchment lead to modifications in the lake ecosystem itself [26].
Phylogenetic biogeography is usually used to reconstruct the evolutionary relationships of species and to trace their origin and past geographic distribution, but, recently, tumor biogeography has been introduced to reconstruct evolutionary relationships, genetic divergences, extinction of cancer cells events, and cancer cell migration strategies [27]. Chroni and Kumar (2021) proposed that tumors can be considered evolutionary island-like ecosystems that undergo evolutionary and spatio-temporal dynamic processes that shape tumor microenvironment (TME) and drive cancer cell migration [28]. Gatenby et al. (2020) hypothesized that large, diverse, and disseminated cancer cell populations could be eradicated using similar eco-evolutionary dynamics of Anthropocene species extinctions as an alternative model for cancer treatment [29]. Drawing parallels and contrasts between dormancy in cancer and other ecosystems in nature, Miller et al. (2021) emphasized the potential for studies in cancer to provide insights into the evolutionary ecology of dormancy [30]. Gatenbee et al. (2020) used species distribution modeling (SDM) or histoecology-based models to identify critical environmental factors that drive tumor development and predict response to therapy [31]. Additionally, cancers have been compared with invasive species in terms of invasion, growth, spread, treatment, and outcome, targeting the development of novel paradigms to cure cancer [32][33]. The concept of invasion is widely used in tumor biology, as well as in ecosystems science, providing opportunities to study the mechanisms of invasion at the molecular level [33]. Thus, high-throughput proteomics alone, or in association with complementary transcriptomics and genomics-based approaches, has the ability to identify multiple biomarkers of tumor invasiveness [34]. Multi-omics data, including gene expression, protein expression, miRNA, DNA methylation, and somatic mutation, may provide a comprehensive characterization of invasiveness-related molecular features across multiple cancer types [35]. Moreover, accumulating evidence showed that tumors act like parasites that fight to survive and optimize their fitness inside the host’s body ecosystem [6]. Experimental evidence suggests that some periods of the day are better that other for cancer proliferation and speed, because the neoplastic cells could exploit or manipulate the host biological rhythms [36]. Thus, tumor development has been compared to infectious disease progression [37].
Also, among other cancer types, BC develops through a process of somatic evolution and arises through natural selection at the cell level [38]. Consequently, cancer has been described as a process of Darwinian evolution [17], the differences and similarities between evolution among somatic cells versus evolution among organisms is a rising discussion [39]. As a clonal evolutionary process, BC may be caused by successive accumulation of genetic alterations followed by clone expansion that leads to tumorigenesis, progression, dissemination, and treatment resistance [40]. Moreover, the mutated tumor cells seem to adapt to microenvironmental niches called a tumor microenvironment (TME) better than normal cells, following different gene–TME interaction patterns that involve the history of genetic and epigenetic changes of the cells and the challenging TME characteristics [41]. Like many species in nature and according to the evolutionary fitness paradigm, cancer cells are constrained by their environments to adopt ecologically driven convergent phenotypes, known as cancer hallmarks, which ensure their fitness within the ecological conditions from body tissues [8]. Like in nature, TME condition includes both abiotic (i.e., local temperature, oxygen partial pressure, nutrient supply, blood flow, pH, intercellular fluid biochemistry) and biotic factors (tumor cell phenotypes, stromal cells, immune cells, vasculature) that interact with heritable features of cancer cells. Evolution by natural selection favors some clonal “species” over others, favoring certain subpopulations of cells that possess genetic and epigenetic traits, which allow for their high proliferation and immune evasion [42]. Advances in new spatial genomics-, transcriptomics- and proteomics-based methods offer new opportunities to study cancer evolution with molecular and spatial detail, for defining the main patterns in which cancer and its TME co-evolve [43].
Consequently, the eco-evolutionary theories are essential for understanding cancer development. In the following sections, the integrated framework of three well known ecological theories: the Bronfenbrenner’s theory of human development, the Vannote’s River Continuum Concept (RCC), and the Ecological Evolutionary Developmental Biology (Eco–Evo–Devo) theory, are used to explain and understand several eco-evo-devo-based principles that govern BC progression.
2. Can Ecological Evolutionary Developmental Biology (Eco-Evo-Devo) Theory Be Applied to BC Development?
The Ecological Evolutionary Developmental Biology (Eco-Evo-Devo) theory has been conceived to explain the complex interaction between an organism’s environment, genes, and developmental processes [44][45]. Initially, this theory had a major goal to demonstrate how the developmental pathways integrate environmental inputs, generating environment-dependent phenotypes [46]. Cancer development within an individual shapes the species evolution and, in turn, species evolution shapes the neoplastic profile of the species as well [1]. Plowman and Plowman (2021) showed that onco-ontogeny recapitulates phylogeny, such that the Haeckel’s controversial biogenetic law emphasizes a certain relevance to cancer behavior [47]. More recently, Liu et al. (2021) presented a framework of a novel theory titled Cancer Evolution-Development (Cancer Evo-Dev), referring to the inflammation-related hepatocarcinogenesis that is sustained by the interaction between genetic predispositions and environmental exposures [48]. Kozlov (2023), considering three major types of biological development—individual, evolutionary, and neoplastic—showed that the Carcino-Evo-Devo theory sustains the evolutionary role of heritable tumors in the evolution of multicellular organisms [49]. Thomas et al. (2017) hypothesized that, apart from related microbiota and parasites, multicellular organisms have a long evolutionary history with communities of cancer cells, known as oncobiota, that influences host life-history traits and survival strategies [50]. Herein, researchers used the integrated framework of two well-known ecological theories: the Bronfenbrenner’s theory of human development and the Vannote’s River Continuum Concept (RCC), to better understand several eco-evo-devo-based principles that govern BC progression.
3. Bronfenbrenner’s Theory
Bronfenbrenner’s theory of human development was first proposed in the 1970s and has been modified over time, evolving from an ecological to a bio-ecological approach [51]. In the BC field, adaptations of Bronfenbrenner’s ecological systems theory have been used in order to assess the role of personal characteristics and needs, dynamic influences of time, interaction with family and health professionals, palliative care services/systems and societal influences involved in BC monitoring and management at a social level [52]. Bronfenbrenner’s ecological systems theory sustains that individual system development is influenced by a series of interconnected environmental systems (Figure 1), such as the microsystem, mesosystem, exosystem, macrosystem, and chronosystem [51].
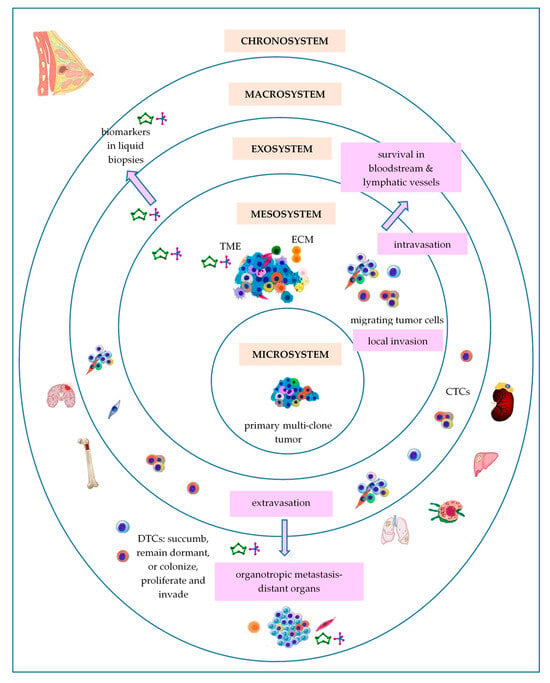
Figure 1. Bronfenbrenner’s theory applied to BC development. Abbreviations: CTCs—circulating tumor cells; DTCs—disseminated tumor cells; ECM—extracellular matrix; TME—tumor microenvironment.
3.1. Primary Breast Tumor as a Microsystem
Considering BC cells of origin as a central ring of Bronfenbrenner’s model, the microsystem is the most proximal setting, with particular physical characteristics, in which these cells are situated. Like all living systems, BC cells of origin are able to self-renew [53]. According to the original theory of cancer stem cells (CSCs), BC, as well as most solid tumors, may originate in a unique cell named a breast cancer stem cell (BCSC), due to an aberrant genetic event that has occurred into a mammary stem cell (MaSC) or within a progenitor cell, even long before the cancer diagnosis. Further, BCSCs retain the immortal proprieties of MaSCs throughout tumorigenesis. BCSCs, also called tumor-initiating cells (TICs), are capable of long-term self-renewal and differentiation, and may divide symmetrically or asymmetrically [54]. Finally, BCSCs account for only 0.1–1% of all tumor cells, representing a minor but significant subpopulation of undifferentiated cells in a tumor [55]. The clonal evolution model sustains that BCSCs might arise through the reprogramming of differentiated cells of mammary tissue, such as luminal and myoepithelial cells, which emphasize a specific mammary mutational profile, suffer a de-differentiation process, and regain stem-like proprieties that lead to new generations of BCSCs. Apart from the auto/self-renewal and stemness capacity, BCSCs have clonal tumor initiation capacity and clonal long-term repopulation potential sustained by specific self-renewal signaling pathways (SRPs) that are commonly dysregulated due to genomic and epigenomic changes [56][57]. Heyde et al. (2019) showed that intra-tumoral heterogeneity emerged as an inevitable consequence of cancer development so that many clones coexist in primary tumor at the time of cancer diagnosis [58]. Additionally, distinct from this classical linear pattern for the evolution of a unique cancer founder, a branching model of evolution of multiple cancer founders may be possible during cancer development [59]. Then, these related clones dissipate and invade the local environment and expand along mammary ducts and occupy a large area in the breast [59].
All living systems have, as a general feature, a hierarchical organization [60]. The theory of CSCs suggests that there is a hierarchy in cancer cells, in which CSCs lie at the top level [54]. Cole et al. (2020) showed that many heterogenic cancers, including BC, are organized into hierarchical structures, based on differentiation capacity of CSCs, the differentiation hierarchy model of cancer being dependent on the gain and loss of the different biomarkers used to characterize the different CSCs populations [61]. Moreover, a continuum of states from stem to differentiated cells exists, enhancing their adaptability and emerging in difficulties for cancer therapy [54].
Competition is a critical evolutionary mechanism that influences the spatio-temporal and dynamic pattern and persistence of species [62]. Tissues and organs may be considered as “social groups” governed by “societal rules” that allow for normal cells to cooperate, while mutant cells compete and expand their territory with the biomolecular aid of their neighboring non-tumor cells [63]. Some authors have claimed that tumors can be viewed as an “invasive species” [32], which express intelligent behavior [64]. As a species is composed by multiple populations, a tumor is composed by multiple clones. Some of these cells move from primary to distant metastatic sites and develop multiple adaptation strategies to survive in hostile environmental conditions. Madan et al. (2022) showed that cell–cell and cancer–microenvironment competition may be a key mediator of clonal dynamics throughout the neoplastic process, being involved in tumor initiation, field cancerization, tumor outgrowth at the expense of normal tissue, and development of malignant cancers from premalignant lesions [65]. Desjardins-Lecavalier et al. (2023) used a method based on single-cell magneto-optical capture (scMOCa) to isolate fast cells with a highly migratory phenotype from heterogeneous human BC cell populations, exploiting their migratory ability alone [66]. Moreover, the fast cell subpopulations expressed genes associated with cell migration, generated a high number of CTCs and soft tissue metastases in mouse models, and retain their high migration speed and focal adhesion dynamic over many generations due to their motility-related transcriptomic profile [66].
As any other living systems, BC microsystems emphasize the capacity to adapt and cope, according to adaptive capacity of complex ecosystems [67][68]. Tumor cells have the ability to reprogram their phenotype, in terms of shape, structure, metabolism, and behavior, in order to cope/adapt with/to environmental challenges, such as local hypoxia, acidity, local temperature, low nutrient supply, or chemotherapy [69]. For example, the oxygen content of primary tumor tissue is an inductor for metastatic cascade [70]. The mean partial pressure of oxygen (PO2) inside the breast tumors ranges from 2.5 to 28 mm of mercury (Hg) (usually < 0.1–5% in solid tumors [71]), with a median value of 10 mm Hg, as compared with 65 mm Hg in normal breast tissue [70]. A starvation-pseudostarvation model suggests that metastasis may be induced by starvation due to oxygen or nutrient limitations that cannot sustain the cell growth and proliferation, or by pseudostarvation imposed by oncogenic activation or microenvironmental signals that converge in translation reprogramming [72]. Lozano et al. (2020) developed a computational thermal model of BC, showing that the metabolic heat generation rates reached as high as 20,000 W/m3 for normal breast tissue and ranged between 100,000–1,200,000 W/m3 for cancerous breast tissue in the case of triple-negative BC (TNBC) [73]. The contact thermography applied to women with primary invasive ductal carcinoma (IDC) showed that the temperature of tumor was 1.79 ± 0.88 °C higher that of the surrounding tissue, in correlation to the microvessels density [74]. Moreover, tumor acidity affects most steps in the metastatic cascade [75]. Cancer cells have increased intracellular pH (pHi) and decreased extracellular pH (pHe) compared to normal cells that also affects cell proliferation, metabolic adaptation, and tumorigenesis by altering the structure and function of pH-sensor proteins [76]. Thus, the pHi ranges from 7.3 to 7.6, while the pHe of solid tumors ranges from 6.5 to 6.9, whereas pH ranges between 7.2 to 7.4 in normal tissues, compelling cancer cells to develop interrelated mechanisms of adaptation to acidity for survival, such as autophagy, increased lysosomal turnover and redistribution, or increased intracellular lipid droplets density [77].
3.2. Breast Tumor as a Mesosystem
According to Bronfenbrenner’s ecological systems theory, the mesosystem is the level of complex relationships among two or more microsystems, allowing the microsystem’s development [51].
It is known that tissues provide the context for cancer cell development and progression [78]. Thus, the primary/local tumor microenvironment (TME) may be considered as a mesosystem, in which heterogeneous populations of tumor cells that form the microsystem of the primary tumor interact with other microsystems represented by different types of non-tumor cells, such as infiltrating inflammatory cells (tumor infiltrating lymphocytes (TILs), dendritic cells (DCs), macrophages, and neutrophils) and stromal cells (fibroblasts, myoepithelial cells, adipocytes, endothelial cells, and pericytes), as well as the extracellular matrix (ECM) that includes various soluble and physical factors, such as enzymes, cytokines, growth factors, pH, or oxygen levels, to facilitate tumorigenesis. This interaction plays crucial roles in tumor proliferation, invasion, and therapies response [79]. Hence, the tumor cells modify the niche that they occupy to offer them a selected advantage over heathy cells [6]. Mayer et al. (2023), using single-cell RNA-sequencing (scRNA-seq), demonstrated that the human BC TME shows a hierarchical network structure of cell–cell interactions, dominated by cancer-associated fibroblasts (CAFs) [80], the most abundant and heterogeneous cellular components of the TME [81]. Local normal fibroblasts and mesenchymal cells adapt to the mesosystem local conditions, becoming activated CAFs and myofibroblasts, which reshape the ECM by “dermoplastic reaction” [81]. Moreover, CAFs may be derived from other multiple cell types, such as epithelial cells, endothelial cells, bone marrow-derived mesenchymal cells, transdifferentiated adipocytes, transdifferentiated smooth muscle cells, and resident stem cells, that can form distinct populations in the same stroma [82]. It was also demonstrated that differential oxygenation in TME modulates macrophages, the most abundant immune cell population in the majority of solid tumors, and cancer cell crosstalk [71].
The tumor mesosystem is the place of cell–cell competition, prey–predator and/or host–parasite interactions, and cell migration. Thus, tumor-immune cells interactions may be imperfectly compared with predatory-prey competition in nature [83]. In the tumor mesosystem there is a high competition for space and low resource supply (i.e., oxygen, glucose or other nutrients) that selects for tumor cells to adopt a dispersal-like behavior from the primary site and to colonize distant organs, while spreading slows when the resource supply is high [84]. Chang et al. (2025) showed that glucose consumption by tumor cells metabolically restricts and suppresses T cells functionality and, consequently, the nutrient competition could determine cancer progression [85]. Moreover, according to Taylor et al. (2017), the starved MCF7 BC cells may suffer epigenetic changes that persist across many generations [84]. The Warburg effect leads to an acidic TME that breaks down the ECM and promotes angiogenesis [86]. The interspecific relationships between human and its symbionts and parasites, which have shaped and are still shaping the human genome, are largely involved in evolution of living organisms [87]. Within this framework, some evidence identified a unique microbial community in breast tissue, as well as in breast tumors, that have their own distinct microbial community that may lead to the occurrence and BC development, for example, by different bacterial toxins that cause DNA damage or regulation of local and systemic immune response [88].
3.3. Breast Tumor as an Exosystem
The Exosystem represents the third ring of Bronfenbrenner’s ecological model. In tumor progression, the intravasation and circulation of tumor cells as circulating tumor cells (CTCs) into the bloodstream or lymphatic vessels towards colonization of pre-niches in preferentially distant sites after extravasation may be considered as main metastatic processes that define the tumor exosystem functionality. Generally, solid tumors give rise to heterogeneous populations of CTCs that evade and contribute to tumor dissemination and progression. Some authors suggest that only 0.2–2% of the tumor cells could form micrometastasis at distant organs [89], or approximately 0.01% of CTCs infiltrate and eventually colonize distant organs [90].
In nature, species migration requires eco-morphological, eco-physiological and behavioral adaptations and affects ecosystem processes [91]. Malignant cell migration, dissemination and invasion, through individual (single) cell migration (amoeboid and mesenchymal invasion), and collective cell migration, when multiple cells retain cell–cell connections and migrate coordinately, are essential for metastatic disease [92]. Moreover, cancer cells may adapt their motility patterns by reversible mesenchymal-amoeboid/amoeboid-mesenchymal transitions (MAT/AMT) and individual-collective/collective-individual transitions, such as amoeboid-collective/collective-amoeboid transitions (ACT/CAT) and mesenchymal-collective transition/collective-mesenchymal transitions (MCT/CMT) [92]. Once detached from the primary site, CTCs encounter the bloodstream and lymphatic vessels microenvironments, hostile habitats that stimulate them to adopt the best strategies to survive, such as phenotypic adaptation based on their phenotypic plasticity and metabolic reprogramming, adapting to supportive niches, and collective adaptation [64]. Thus, EMT is involved in genesis of CTCs [93]. EMT is also known as an important embryological step in the gastrulation, heart, musculoskeletal system, or peripheral nervous system organogenesis [93]. Into the metastatic cascade, there are two opposite cellular strategies that alter cancer cell shape: the epithelial-to-mesenchymal transition (EMT) and mesenchymal-to-epithelial transition (MET). During EMT, cancer cells lose epithelial specific biomarkers (i.e., E-cadherin, cluster of differentiation CD44v variant isoform), while acquiring mesenchymal biomarkers (i.e., N-cadherin, fibronectin (FNT), vimentin (VIM), smooth muscle actin (α-SMA), β-catenin (CTNNB), CD44s standard isoform) [94]. Inversely, during MET in distant organs, CTCs abandon their mesenchymal phenotype. Moreover, EMT is also associated with a complex metabolic reprogramming based on mutations in metabolic genes, which support the energy requirements of increased motility and growth in harmful environmental conditions [95]. Consequently, the shift between these very different environments is costly, as in the case of the salmon species that spend their first months in freshwater, and then migrate to saltwater for several years before returning to natal rivers to spawn [96]. Nicolazzo et al. (2023) compared the EMT of tumor cells-to-CTCs with shape-shifter birds and their metabolic reprogramming with naked mole-rats, the most hypoxia-tolerant mammal, as “metabolism switchers” [64].
3.4. Breast Tumor as a Macrosystem
BC cells preferentially metastasize to several organs, such as bone (30–60%), lung (21–32%), liver (15–32%), brain (4–10%), known as organotropic metastasis [97]. Thus, metastatic BC cells may be considered as “seeds”, while the microenvironment of the metastatic niche may be viewed as the “soil”, so that the metastatic niche formation depends on “seeds-soil” interactions [98]. It is known that organ-specific stromal cells release signaling proteins that induce chemotaxis, the ability to move in the direction of a chemical gradient, causing organotropic metastasis [99]. The metastatic organotropism depends on the subtype of BC, host organ microenvironment, and cell–cell and cell–matrix interactions. Thus, invasive lobular carcinomas (ILCs) has three times more metastases in the peritoneum, gastrointestinal tract, and ovaries comparative to invasive ductal carcinoma (IDC), which “prefers” to metastasize to the lungs, distant lymph nodes, and central nervous system [100]. The “seeds-soil” interactions facilitate: (i) pre-metastatic niche formation under the influence of factors released by cancer cells before their arrival at distant organs; (ii) metastatic niche formation, and (iii) interaction between disseminated tumor cells (DTCs) and local resident cells, assuring the cancer cell survival and formation of metastatic lesions [101]. Heyde et al. (2019) showed that 10 to 150 cells may seed each metastatic pre-niche and only a fraction of genetic diversity into a primary tumor is passed on to metastases [58]. Schrijver et al. (2018) showed that the main BC gene-drivers have been altered in both the primary tumors and their metastases, but they also identified genetic alterations restricted either to the primary tumor or within metastases to metastasis [102]. In the metastatic niche, DTCs form a metastasis or enter in the dormant period, when cancer cells exit the cell cycle, arrest their growth, and become immune to drugs that usually target cells in mitotic division [103]. Dormancy is known as a reversible state of reduced metabolic activity, known as a dynamic mechanism by which the organisms/cells respond to periods of unfavorable environmental/microenvironmental stress, including temperature, low nutrients, or toxins [104]. For example, the bone is considered a particularly hypoxic environment, where the oxygen levels range from <1% to 6% (approximately 7–43 mm Hg), compared with most normal tissues, where oxygen levels ranges between 2% and 9% (14–65 mm Hg) [105]. Moreover, in the bone marrow, the oxygen level ranges between 1% and 4% (7–29 mmHg) [105]. Smoking promotes lung metastasis of BC, because it generates a neutrophil-dependent pulmonary inflammatory microenvironment [98]. In the brain, BC cells which pass through the blood-brain barrier (BBB), interact with astrocytes, and release matrix metalloproteinases (MMPs) that destroy the collagen network, promoting tumor cells growth and development [100].
Dormant DTCs may reactivate after a long period of dormancy and become a source of BC recurrence [106]. Usually, stromal inflammation might reactivate these cells, inducing growth and a mesenchymal phenotype to dormant ER+ cells with an epithelial phenotype [107].
3.5. Breast Tumor as a Chronosystem
The chronosystem is the fifth outermost ring in the Bronfenbrenner’s ecological model that contains elements of time. Cancers begin when the first cell undergoes malignant transformation [108]. Various mathematical models have been used to explore the natural progression of BC [109]. Cancers may evolve over variable time frames that range from 1 to 50 years [78]. It is known that cancer cells double 20 to 30 times to reach 1 mm3 to 1 cm3 [108]. Thomas et al. (2017) showed that, during the organism’s lifespan, the duration of the interaction between the host and its oncobiota can vary from months to years, sometimes decades [50]. Relative to the time of the first cancer-drive mutation, metastasis has been described as a late event in the natural molecular history of cancer [110]. In BC, metastasis can occur early when the primary tumor is <1 mm in diameter, even with 2–4 years prior to diagnosis of the primary tumor [110]. Menes et al. (2015) have reported that the 10-year incidence of a second primary BC was highest in BRCA1 mutation carriers [111]. Moreover, according to both “the sick breast lobe” hypothesis and the “theory of biological timing”, BC development is subjected to biological timing of transformation of a large number of epithelial cells or stem cells of the sick lobe in tumor cell clones after several cell generations in large carcinomas [22]. Thus, the time needed for this transformation, which may be several decades, the number and location of transformed cells, and the differences in their transformation patterns determine the individual morphology and behavior of BC.
4. Vannote’s River Continuum Concept (RCC)
Chroni and Kumar (2021) categorized tumor ecosystems into islets, islands, and archipelagos-like ecosystems that may transform from one type into another [28]. Here, researchers aim to discuss cancer progression as a watercourse. The River Continuum Concept (RCC) ecological theory, first developed by Robin L. Vannote et al. in 1980 [112], is considered a “milestone in stream ecology”, due to its comprehensive description and evaluation of the structure and function of river systems [113]. This theory is based on the concept of dynamic equilibrium of a watercourse as an open ecosystem in constant interaction with the bank, moving from headwaters to mouth/sea, and links physical variables with patterns in biodiversity, functionality, and metabolism dynamics, that results in downstream gradients in communities composition and ecosystem processes [113]. Vannote et al. (1980) showed that in natural stream systems, biological communities form a temporal continuum of synchronized species replacements [112]. Comparatively, BC tumorigenesis and metastasis represent a complex cascade/continuum of cells and biomarkers that comprises successively integrated populations of heterogeneous tumor cells and cancer-associated cells, interacting in a dynamic spatio-temporal fashion under specific microenvironmental conditions (Figure 2 and Table 1). Thus, akin to the species of biological communities that form a spatial continuum in a watercourse or in other ecosystem types, cancer cells should have the ability to: (i) move to more hospitable environments when local conditions in primary tumor sites become unfavorable, like in case of hypoxia, nutritional and thermoregulatory stress or acidic pHe, known to promote cell invasion [114]; (ii) move to minimize the cell–cell competition for local resources; (iii) move in biomolecular gradients, following directional migration patterns that may be cell type specific [115] and adjusted by different growth factors and chemokines from the primary TME [116]; (iv) don’t waste effort moving in wrong directions before drifting in the correct direction; and (v) should arrive at a specific distant site in a timely manner based on chemotactic-based behavior that requires sensing, polarizations, and directional motility, emphasizing accuracy, speed and persistence [117]. Moreover, metastasis is a selective process that favors cells with higher deformability and motility [118]. Migrating animals possess ecomorfological adaptations and can often move together, so that collective factors and sensory cues could shape migration [119]. Similarly, increasing evidence suggests that clustered CTCs (microemboli) resist better in circulatory system and emphasize higher metastatic capacities that single CTCs [120]. Migratory animals can adopt adaptive strategies, modifying their behavior, life-history, and physiology through phenotypic plasticity [121]. Similarly, CTCs enter the bloodstream, where they are exposed to immunological insults from leukocytes, to collision with erythrocytes, and to interaction with activated platelets and macrophages, so that only a small fraction of these cells are able to complete the metastatic process, colonizing pre-metastatic niches from distant organs [122]. Kareva (2015) showed that only a small fraction of cells from primary tumors are successful in establishing distant metastasis, a similar process occurs in nature and is known as the ecological succession [6].
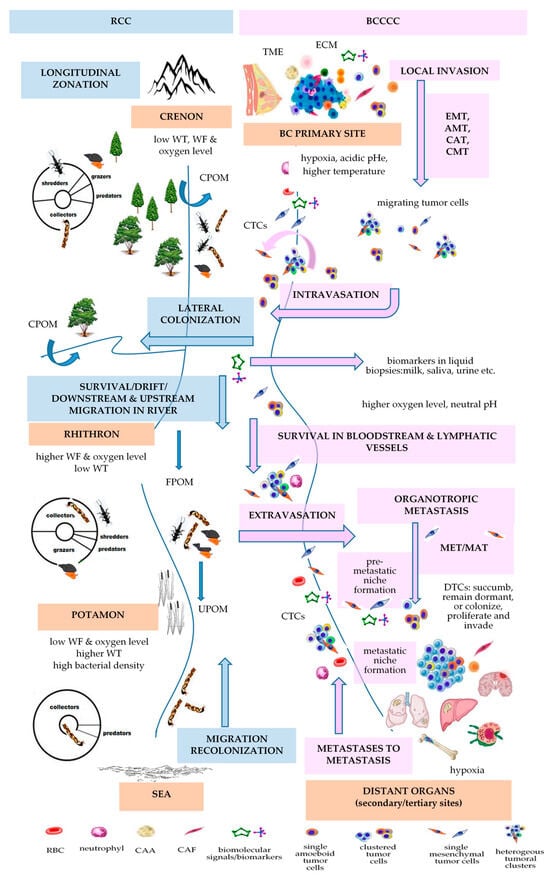
Figure 2. Vannote’s River Continuum Concept (RCC) principles applied to BC development (BCCCC). Abbreviations: AMT—amoeboid-mesenchymal transition; BCCCC—Breast Cancer Cell Continuum Concept; CAA—cancer-associated adipocyte; CAF—cancer-associated fibroblast; CAT—collective-amoeboid transition; CMT—collective-mesenchymal transition; CPOM—coarse particulate organic matter; CTCs—circulating tumor cells; DTCs—disseminated tumor cells; ECM—extracellular matrix; EMT—epithelial-mesenchymal transition; FPOM—fine particulate organic matter; MAT—mesenchymal-amoeboid transition; MET-mesenchymal-to-epithelial transition; pHe—extracellular pH; RBC—red blood cell; RCC—River Continuum Concept; TME—tumor microenvironment; UPOM—ultrafine particulate organic matter; WT—water temperature; WF—water flow.
Genomics, transcriptomics, proteomics, secretomics or exosomics-based approaches are essential to define the BC continuum (BCC) from primary to metastatic sites. During this metastatic continuum, dissemination of CTCs is a crucial step. CTCs may even assure the “tumor self-seeding”, also colonizing their tumors of origin, where they actively contribute to tumorigenesis [123]. These cells are extremely heterogeneous and form phenotypically and genotypically distinct rare subpopulations of highly active tumor cells released into the bloodstream from primary cancers and metastases that reflect the status of tumor genotypes, as long as mutations in known driver genes found in primary tumor and metastases were also detected in corresponding CTCs together with mutations exclusively observed in CTCs [124][125]. Evidence demonstrates that CTCs undergo modification in response to the dynamic biophysical environment in the bloodstream, partially due to fluid shear stress that generates reactive oxygen species (ROS), which affect mitochondria, causing aberrant energy metabolism, oxidative stress (OS), and cell death pathways that sustain cancer invasiveness [126]. In RCC theory, Vannote showed that the longitudinal distribution and lateral colonization of biotic communities primarily depends on abiotic factor gradients. Similarly, during the metastatic cascade, CTC subpopulations migrate from the hypoxic primary TME to hypoxic pre-metastatic niches (PMN) of metastatic distant sites, through the bloodstream, also considered as a hostile environment for CTCs, where the oxygen level is much higher than in most solid tumors. Proteomics-based techniques have been used to distinguish the proteome landscape of distant metastasis derived from primary breast tumors [127], to identify circulating proteins from serum or plasma, taking into account that proteins can act as the primary “bio-effectors” of metastasis [128], or to identify the epithelial, mesenchymal, and stemness biomarkers to characterize the CTC subpopulations [93]. Secretomics and mass spectrometry (MS)-based proteomics combinatorial analysis identified the stromal proteome of BC, including the protein–protein interaction (PPI) network, according to luminal-like and basal-like phenotypes, emphasizing secreted proteins that are increased by hypoxia [129]. The genomic landscape of cancer and the evolution during treatment may be non-invasively assessed by biomolecular characterization of CTCs [123]. Next generation sequencing (NGS)-based strategies allowed sequencing cancer genomes to reveal the subclonal diversification of primary BC [130]. The development of whole genome amplification (WGA) followed by NGS, microarray-based comparative genome hybridization (array-CGH), and single-circulating tumor cell sequencing techniques are able to profile single CTC [123]. Moreover, genomics-based approaches showed that plasma circulating cell-free DNA (cfDNA) and CTCs from the same blood sample provided complementary mutation information [131].
On the other hand, BC cells, primary tumor stroma and specific stromal components in distant organs intercommunicate and dictate the continuum of metastatic process through exosomes. These nano-vesicles are crucial mediators that transfer molecular signals/bioactive molecules, such as lipids, proteins, and different types of RNAs, such as microRNAs (miRNAs), mRNAs, transfer RNA, ribosomal RNA, small nuclear RNA, small nucleolar RNA, piwi-interacting RNA and long-non coding RNA, which are essential for intercellular communication [132]. Thus, tumor-derived exososmes contribute to generation of pre-metastatic niches, sustain the cancer dissemination, colonization, survival, proliferation or dormancy of incoming metastatic tumor cells, like in the case of BC cell-derived exosomes that transfer miR-21 to osteoclasts, promoting BC bone metastatic lesions [133]. Miller et al. (2021) sustained that even dormancy is a continuum of dormancy phenotypes characterized by hypometabolism, reduced feeding, reproduction or proliferation [30]. Thus, cancer cells exhibit a continuum of states from quiescence to long-term dormancy characterized by lack of cell division.
The researchers previously reviewed and defined a Breast Cancer Cell Continuum Concept (BCCCC) comprised of successively integrated and interactive populations of heterogeneous tumor and CACs and sustained by a Breast Cancer Proteomics Continuum Concept (BCPCC), where each phenotype of neoplastic and CACs is characterized by a changing and adaptive proteomics profile [9]. Some authors delineated an epigenetic monoclonal progression continuum from normal to benign to invasive BC, based on DNA hypermethylation profiles [134]. Sanati et al. (2019) sustained that ductal carcinoma in situ (DCIS) has a continuum of histologically diverse proliferations that range from very well to very poorly differentiated [135]. Evidence emphasized the transition between an epithelial to a more mesenchymal cell state as an EMT continuum based on a continuum of multiple intermediate phenotypes of EMT transformation [136][137][138][139]. Thus, single-cell spatial transcriptomics based on RNA-sequencing has been preferentially used to characterize the EMT continuum [139][140].
Table 21. Vannote’s River Continuum Concept (RCC) applied to BC development (Breast Cancer Cell Continuum Concept (BCCCC) and Breast Cancer Proteomic Continuum Concept (BCPCC)).
| RCC | [112] | BCCCC and BCPCC | [9] | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Longitudinal Changes in the Benthic Communities in Temperate Rivers | Longitudinal Changes in Kinetics of Metastasis | ||||||||||||||
| ecological zonation | gradient of physical variables [141] | gradient of biological communities | gradient of energy input | BC progression | gradient of physical variables | BCCCC | BCPCC | ||||||||
| headwaters or crenon | water temperature, flow, and oxygen level are low | shredders, collectors, grazers, predators |
CPOM | primary breast tumor | PO2: 2.5–28 mm Hg (mv 10 mm Hg) [70]/<0.1–5% [71]; MR: 100,000–1,200,000 W/m3 [73]; temperature with 1.79 ± 0.88 °C higher that of the surrounding tissue [74]; pHi: 7.3–7.6 & pHe: 6.5–6.9 [77] |
tumor cells; stromal cells (CAFs, TECs, TAPs, CAAs); immune cells (TAMs, TAMCs, TANs, TALs, TAPs, MDSCs); surrounding normal cells (luminal and myoepithelial cells); ECM |
stem-like markers (CD44high/CD24low, EpCAM, PI3K, ALDH1+) | ||||||||
| MIGRATION/DRIFT/COLONIZATION | EMT/INTRAVASATION | ||||||||||||||
| rhithron | high water current and dissolved oxygen; low temperature |
collectors, grazers, shredders, predators | FPOM, UPOM | bloodstream or lymphatic vessels | blood: oxygen level 12% [71] | CTCs | epithelial markers (EpCAM, E-cadherin, CKs, ZO, ESPR1); mesenchymal-like markers (N-cadherin, VIM, Twist1, AKT and PI3K, ZEB1); stemness-like markers (ALDH1, CD44, gangliosides, ABC proteins) [93] |
||||||||
| MIGRATION/DRIFT/COLONIZATION | ETRAVASATION/MET/COLONIZATION | ||||||||||||||
| potamon | low speed; low oxygen content; sandy river bed; higher water temperature; higher bacterial density |
collectors, predators | FPOM, UPOM | preferred BC distant metastatic sites | bone: oxygen levels <1–6% (7–43 mmHg) [105] | homing and dormant DTCs | overexpression of epithelial markers (E-cadherin, occludin, crumbs3); downregulation of mesenchymal markers [142] | ||||||||
Abbreviations: ALDH1—aldehyde dehydrogenase-1; AKT—protein kinase B; BCCCC—Breast Cancer Cell Continuum Concept; BCPCC—Breast Cancer Proteomic Continuum Concept; CAAs—cancer-associated adipocytes; CAFs—cancer-associated fibroblasts; CKs—cytokeratins; CPOM—coarse particulate organic material; CTCs—circulating tumor cells; DTCs—disseminated tumor cells; ECM—extracellular matrix; EMT-epithelial-mesenchymal transition; EpCAM—epithelial cellular adhesion molecule; ESPR1-epithelial splicing regulator 1; FPOM—fine particulate organic material; pHe—extracellular pH; pHi—intracellular pH; MDSCs—myeloid-derived suppressor cells; MET—mesenchymal-epithelial transition; MR—metabolic rate; mv—median value; PI3K –phosphoinositide 3-kinase; PO2—oxygen partial pressure; RCC—River Continuum Concept; TALs—tumor-associated lymphocytes; TAMCs—tumor-associated mast cells; TAMs—tumor-associated macrophages; TANs—tumor-associated neutrophils; TAPs –tumor-associated pericytes; TECs—tumor endothelial cells; TICs—tumor-initiating cells; TME—tumor microenvironment; UPOM—ultra-fine particulate organic material; VIM—vimentin; ZEB1—zinc finger E-box binding homeobox 1; ZO—zonula occludens.
References
- Casás-Selves, M.; Degregori, J. How cancer shapes evolution and how evolution shapes cancer. Evol. Educ. Outreach 2011, 4, 624–634.
- Maruki, T.; Ye, Z.; Lynch, M. Evolutionary Genomics of a Subdivided Species. Mol. Biol. Evol. 2022, 39, msac152.
- Frank, S.A. Somatic evolutionary genomics: Mutations during development cause highly variable genetic mosaicism with risk of cancer and neurodegeneration. Proc. Natl. Acad. Sci. USA 2010, 107 (Suppl. S1), 1725–1730.
- Baer, B.; Millar, A.H. Proteomics in evolutionary ecology. J. Proteom. 2016, 135, 4–11.
- Reynolds, B.A.; Oli, M.W.; Oli, M.K. Eco-oncology: Applying ecological principles to understand and manage cancer. Ecol. Evol. 2020, 10, 8538–8553.
- Kareva, I. Cancer Ecology: Niche Construction, Keystone Species, Ecological Succession, and Ergodic Theory. Biol. Theory 2015, 10, 283–288.
- Chen, X.; Song, E. The theory of tumor ecosystem. Cancer Commun. 2022, 42, 587–608.
- Somarelli, J. The Hallmarks of Cancer as Ecologically Driven Phenotypes. Front. Ecol. Evol. 2021, 9, 661583.
- Neagu, A.-N.; Whitham, D.; Buonanno, E.; Jenkins, A.; Alexa-Stratulat, T.; Tamba, B.I.; Darie, C.C. Proteomics and its applications in breast cancer. Am. J. Cancer Res. 2021, 11, 4006–4049.
- Boddy, A.M. The need for evolutionary theory in cancer research. Eur. J. Epidemiol. 2022, 38, 1259–1264.
- Coscieme, L.; Pulselli, F.M.; Jørgensen, S.E.; Bastianoni, S.; Marchettini, N. Thermodynamics-based categorization of ecosystems in a socio-ecological context. Ecol. Model. 2013, 258, 1–8.
- Modaresi Movahed, T.; Jalaly Bidgoly, H.; Khoshgoftar Manesh, M.H.; Mirzaei, H.R. Predicting cancer cells progression via entropy generation based on AR and ARMA models. Int. Commun. Heat Mass Transf. 2021, 127, 105565.
- Myers, K.V.; Pienta, K.J.; Amend, S.R. Cancer Cells and M2 Macrophages: Cooperative Invasive Ecosystem Engineers. Cancer Control J. Moffitt Cancer Cent. 2020, 27, 1073274820911058.
- Sell, S.; Nicolini, A.; Ferrari, P.; Biava, P.M. Cancer: A Problem of Developmental Biology; Scientific Evidence for Reprogramming and Differentiation Therapy. Curr. Drug Targets 2016, 17, 1103–1110.
- Edwards, P.A.W. The Impact of Developmental Biology on Cancer Research: An Overview. Cancer Metastasis Rev. 1999, 18, 175–180.
- Zhu, X.; Li, S.; Xu, B.; Luo, H. Cancer evolution: A means by which tumors evade treatment. Biomed. Pharmacother. 2021, 133, 111016.
- Thomas, F.; Fisher, D.; Fort, P.; Marie, J.-P.; Daoust, S.; Roche, B.; Grunau, C.; Cosseau, C.; Mitta, G.; Baghdiguian, S.; et al. Applying ecological and evolutionary theory to cancer: A long and winding road. Evol. Appl. 2013, 6, 1–10.
- Chouaib, S.; Lorens, J. Editorial: Targeting the Tumor Microenvironment for a More Effective and Efficient Cancer Immunotherapy. Front. Immunol. 2020, 11, 933.
- Kotler, B.P.; Brown, J.S. Cancer Community Ecology. Cancer Control J. Moffitt Cancer Cent. 2020, 27, 1073274820951776.
- Dujon, A.M.; Aktipis, A.; Alix-Panabières, C.; Amend, S.R.; Boddy, A.M.; Brown, J.S.; Capp, J.-P.; DeGregori, J.; Ewald, P.; Gatenby, R.; et al. Identifying key questions in the ecology and evolution of cancer. Evol. Appl. 2021, 14, 877–892.
- Tot, T. Breast Cancer: A Lobar Disease; Springer: London, UK, 2008; pp. 1–216.
- Tot, T. The Theory of the Sick Breast Lobe and the Possible Consequences. Int. J. Surg. Pathol. 2007, 15, 369–375.
- Tot, T. DCIS, cytokeratins, and the theory of the sick lobe. Virchows Arch. 2005, 447, 1–8.
- Tan, M.; Tot, T. The sick lobe hypothesis, field cancerisation and the new era of precision breast surgery. Gland. Surg. 2018, 7, 611–618.
- Li, L.; Sullivan, P.L.; Benettin, P.; Cirpka, O.A.; Bishop, K.; Brantley, S.L.; Knapp, J.L.A.; van Meerveld, I.; Rinaldo, A.; Seibert, J.; et al. Toward catchment hydro-biogeochemical theories. WIREs Water 2021, 8, e1495.
- Jiménez-Navarro, I.C.; Mesman, J.P.; Pierson, D.; Trolle, D.; Nielsen, A.; Senent-Aparicio, J. Application of an integrated catchment-lake model approach for simulating effects of climate change on lake inputs and biogeochemistry. Sci. Total Environ. 2023, 885, 163946.
- Chroni, A.; Miura, S.; Oladeinde, O.; Aly, V.; Kumar, S. Migrations of cancer cells through the lens of phylogenetic biogeography. Sci. Rep. 2021, 11, 17184.
- Chroni, A.; Kumar, S. Tumors Are Evolutionary Island-Like Ecosystems. Genome Biol. Evol. 2021, 13, evab276.
- Gatenby, R.A.; Artzy-Randrup, Y.; Epstein, T.; Reed, D.R.; Brown, J.S. Eradicating Metastatic Cancer and the Eco-Evolutionary Dynamics of Anthropocene Extinctions. Cancer Res. 2020, 80, 613–623.
- Miller, A.K.; Brown, J.S.; Enderling, H.; Basanta, D.; Whelan, C.J. The Evolutionary Ecology of Dormancy in Nature and in Cancer. Front. Ecol. Evol. 2021, 9, 676802.
- Gatenbee, C.D.; Minor, E.S.; Slebos, R.J.C.; Chung, C.H.; Anderson, A.R.A. Histoecology: Applying Ecological Principles and Approaches to Describe and Predict Tumor Ecosystem Dynamics Across Space and Time. Cancer Control J. Moffitt Cancer Cent. 2020, 27, 1073274820946804.
- Noorbakhsh, J.; Zhao, Z.-M.; Russell, J.C.; Chuang, J.H. Treating Cancer as an Invasive Species. Mol. Cancer Res. MCR 2020, 18, 20–26.
- Neinavaie, F.; Ibrahim-Hashim, A.; Kramer, A.M.; Brown, J.S.; Richards, C.L. The Genomic Processes of Biological Invasions: From Invasive Species to Cancer Metastases and Back Again. Front. Ecol. Evol. 2021, 9, 681100.
- Pouliquen, D.L.; Boissard, A.; Coqueret, O.; Guette, C. Biomarkers of tumor invasiveness in proteomics (Review). Int. J. Oncol. 2020, 57, 409–432.
- Bi, G.; Liang, J.; Zheng, Y.; Li, R.; Zhao, M.; Huang, Y.; Zhan, C.; Xu, S.; Fan, H. Multi-omics characterization and validation of invasiveness-related molecular features across multiple cancer types. J. Transl. Med. 2021, 19, 124.
- Cortés-Hernández, L.E.; Eslami-S, Z.; Dujon, A.M.; Giraudeau, M.; Ujvari, B.; Thomas, F.; Alix-Panabières, C. Do malignant cells sleep at night? Genome Biol. 2020, 21, 276.
- Arnal, A.; Beckmann, C.; Biro, P.A.; Boidin-Wichlacz, C.; Massol, F.; Mery, F.; Misse, D.; Poulin, R.; Renaud, F.; Roche, B.; et al. Cancer and life-history traits: Lessons from host–parasite interactions. Parasitology 2016, 143, 533–541.
- Gerstung, M.; Jolly, C.; Leshchiner, I.; Dentro, S.C.; Gonzalez, S.; Rosebrock, D.; Mitchell, T.J.; Rubanova, Y.; Anur, P.; Yu, K.; et al. The evolutionary history of 2658 cancers. bioRxiv 2017, 161562.
- Fortunato, A.; Boddy, A.; Mallo, D.; Aktipis, A.; Maley, C.C.; Pepper, J.W. Natural Selection in Cancer Biology: From Molecular Snowflakes to Trait Hallmarks. Cold Spring Harb. Perspect. Med. 2017, 7, a029652.
- Wang, J.; Khiabanian, H.; Rossi, D.; Fabbri, G.; Gattei, V.; Forconi, F.; Laurenti, L.; Marasca, R.; Del Poeta, G.; Foà, R.; et al. Tumor evolutionary directed graphs and the history of chronic lymphocytic leukemia. eLife 2014, 3, e02869.
- Vineis, P. Cancer as an evolutionary process at the cell level: An epidemiological perspective. Carcinogenesis 2003, 24, 1–6.
- Alfarouk, K.O.; Ibrahim, M.E.; Gatenby, R.A.; Brown, J.S. Riparian ecosystems in human cancers. Evol. Appl. 2013, 6, 46–53.
- Seferbekova, Z.; Lomakin, A.; Yates, L.R.; Gerstung, M. Spatial biology of cancer evolution. Nat. Rev. Genet. 2023, 24, 295–313.
- Gilbert, S.F.; Bosch, T.C.G.; Ledón-Rettig, C. Eco-Evo-Devo: Developmental symbiosis and developmental plasticity as evolutionary agents. Nat. Rev. Genet. 2015, 16, 611–622.
- Abouheif, E.; Favé, M.-J.; Ibarrarán-Viniegra, A.S.; Lesoway, M.P.; Rafiqi, A.M.; Rajakumar, R. Eco-Evo-Devo: The Time Has Come. In Ecological Genomics: Ecology and the Evolution of Genes and Genomes; Landry, C.R., Aubin-Horth, N., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 107–125.
- Sultan, S.E. Eco-Evo-Devo. In Evolutionary Developmental Biology: A Reference Guide; Nuno de la Rosa, L., Müller, G., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 1–13.
- Plowman, P.N.; Plowman, C.E. Onco-ontogeny recapitulates phylogeny–A consideration. Oncogene 2021, 40, 1542–1550.
- Liu, W.; Deng, Y.; Li, Z.; Chen, Y.; Zhu, X.; Tan, X.; Cao, G. Cancer Evo–Dev: A Theory of Inflammation-Induced Oncogenesis. Front. Immunol. 2021, 12, 768098.
- Kozlov, A.P. Carcino-Evo-Devo, A Theory of the Evolutionary Role of Hereditary Tumors. Int. J. Mol. Sci. 2023, 24, 8611.
- Thomas, F.; Jacqueline, C.; Tissot, T.; Henard, M.; Blanchet, S.; Loot, G.; Dawson, E.; Mery, F.; Renaud, F.; Montagne, J.; et al. The importance of cancer cells for animal evolutionary ecology. Nat. Ecol. Evol. 2017, 1, 1592–1595.
- Rosa, E.M.; Tudge, J. Urie Bronfenbrenner’s Theory of Human Development: Its Evolution from Ecology to Bioecology. J. Fam. Theory Rev. 2013, 5, 243–258.
- Pask, S.; Pinto, C.; Bristowe, K.; van Vliet, L.; Nicholson, C.; Evans, C.J.; George, R.; Bailey, K.; Davies, J.M.; Guo, P.; et al. A framework for complexity in palliative care: A qualitative study with patients, family carers and professionals. Palliat. Med. 2018, 32, 1078–1090.
- Yeremeyev, I.; Dychko, A.; Remez, N.; Kraychuk, S.; Ostapchuk, N. Problems of sustainable development of ecosystems. IOP Conf. Ser. Earth Environ. Sci. 2021, 628, 012014.
- Majumdar, S.; Liu, S.-T. Cell division symmetry control and cancer stem cells. AIMS Mol. Sci. 2020, 7, 82–98.
- Zhang, X.; Powell, K.; Li, L. Breast Cancer Stem Cells: Biomarkers, Identification and Isolation Methods, Regulating Mechanisms, Cellular Origin, and Beyond. Cancers 2020, 12, 3765.
- Nalla, L.V.; Kalia, K.; Khairnar, A. Self-renewal signaling pathways in breast cancer stem cells. Int. J. Biochem. Cell Biol. 2019, 107, 140–153.
- Plaks, V.; Kong, N.; Werb, Z. The cancer stem cell niche: How essential is the niche in regulating stemness of tumor cells? Cell Stem Cell 2015, 16, 225–238.
- Heyde, A.; Reiter, J.G.; Naxerova, K.; Nowak, M.A. Consecutive seeding and transfer of genetic diversity in metastasis. Proc. Natl. Acad. Sci. USA 2019, 116, 14129–14137.
- Nishimura, T.; Kakiuchi, N.; Yoshida, K.; Sakurai, T.; Kataoka, T.R.; Kondoh, E.; Chigusa, Y.; Kawai, M.; Sawada, M.; Inoue, T.; et al. Evolutionary histories of breast cancer and related clones. Nature 2023, 620, 607–614.
- Miller, W. The Hierarchical Structure of Ecosystems: Connections to Evolution. Evol. Educ. Outreach 2008, 1, 16–24.
- Cole, A.J.; Fayomi, A.P.; Anyaeche, V.I.; Bai, S.; Buckanovich, R.J. An evolving paradigm of cancer stem cell hierarchies: Therapeutic implications. Theranostics 2020, 10, 3083–3098.
- Pelage, L.; Lucena-Frédou, F.; Eduardo, L.N.; Le Loc’h, F.; Bertrand, A.; Lira, A.S.; Frédou, T. Competing with each other: Fish isotopic niche in two resource availability contexts. Front. Mar. Sci. 2022, 9, 975091.
- Johnston, L.A. Socializing with MYC: Cell competition in development and as a model for premalignant cancer. Cold Spring Harb. Perspect. Med. 2014, 4, a014274.
- Nicolazzo, C.; Francescangeli, F.; Magri, V.; Giuliani, A.; Zeuner, A.; Gazzaniga, P. Is cancer an intelligent species? Cancer Metastasis Rev. 2023, 42, 1201–1218.
- Madan, E.; Palma, A.M.; Vudatha, V.; Trevino, J.G.; Natarajan, K.N.; Winn, R.A.; Won, K.J.; Graham, T.A.; Drapkin, R.; McDonald, S.A.C.; et al. Cell Competition in Carcinogenesis. Cancer Res. 2022, 82, 4487–4496.
- Desjardins-Lecavalier, N.; Annis, M.G.; Nowakowski, A.; Kiepas, A.; Binan, L.; Roy, J.; Modica, G.; Hébert, S.; Kleinman, C.L.; Siegel, P.M.; et al. Migration speed of captured breast cancer subpopulations correlates with metastatic fitness. J. Cell Sci. 2023, 136, jcs260835.
- Angeler, D.G.; Fried-Petersen, H.B.; Allen, C.R.; Garmestani, A.; Twidwell, D.; Chuang, W.-C.; Donovan, V.M.; Eason, T.; Roberts, C.P.; Sundstrom, S.M.; et al. Chapter One–Adaptive capacity in ecosystems. In Advances in Ecological Research; Bohan, D.A., Dumbrell, A.J., Eds.; Academic Press: New York, NY, USA, 2019; Volume 60, pp. 1–24.
- Dong, X.; Fisher, S.G. Ecosystem spatial self-organization: Free order for nothing? Ecol. Complex. 2019, 38, 24–30.
- Balcioglu, O.; Heinz, R.E.; Freeman, D.W.; Gates, B.L.; Hagos, B.M.; Booker, E.; Mirzaei Mehrabad, E.; Diesen, H.T.; Bhakta, K.; Ranganathan, S.; et al. CRIPTO antagonist ALK4L75A-Fc inhibits breast cancer cell plasticity and adaptation to stress. Breast Cancer Res. 2020, 22, 125.
- Gilkes, D.M.; Semenza, G.L. Role of hypoxia-inducible factors in breast cancer metastasis. Future Oncol. 2013, 9, 1623–1636.
- Campillo, N.; Falcones, B.; Otero, J.; Colina, R.; Gozal, D.; Navajas, D.; Farré, R.; Almendros, I. Differential Oxygenation in Tumor Microenvironment Modulates Macrophage and Cancer Cell Crosstalk: Novel Experimental Setting and Proof of Concept. Front. Oncol. 2019, 9, 43.
- García-Jiménez, C.; Goding, C.R. Starvation and Pseudo-Starvation as Drivers of Cancer Metastasis through Translation Reprogramming. Cell Metab. 2019, 29, 254–267.
- Lozano, A., 3rd; Hayes, J.C.; Compton, L.M.; Azarnoosh, J.; Hassanipour, F. Determining the thermal characteristics of breast cancer based on high-resolution infrared imaging, 3D breast scans, and magnetic resonance imaging. Sci. Rep. 2020, 10, 10105.
- Yahara, T.; Koga, T.; Yoshida, S.; Nakagawa, S.; Deguchi, H.; Shirouzu, K. Relationship between Microvessel Density and Thermographic Hot Areas in Breast Cancer. Surg. Today 2003, 33, 243–248.
- Rolver, M.G.; Pedersen, S.F. Putting Warburg to work: How imaging of tumour acidosis could help predict metastatic potential in breast cancer. Br. J. Cancer 2021, 124, 1–2.
- White, K.A.; Grillo-Hill, B.K.; Barber, D.L. Cancer cell behaviors mediated by dysregulated pH dynamics at a glance. J. Cell Sci. 2017, 130, 663–669.
- Ibrahim-Hashim, A.; Estrella, V. Acidosis and cancer: From mechanism to neutralization. Cancer Metastasis Rev. 2019, 38, 149–155.
- Greaves, M.; Maley, C.C. Clonal evolution in cancer. Nature 2012, 481, 306–313.
- Li, J.J.; Tsang, J.Y.; Tse, G.M. Tumor Microenvironment in Breast Cancer–Updates on Therapeutic Implications and Pathologic Assessment. Cancers 2021, 13, 4233.
- Mayer, S.; Milo, T.; Isaacson, A.; Halperin, C.; Miyara, S.; Stein, Y.; Lior, C.; Pevsner-Fischer, M.; Tzahor, E.; Mayo, A.; et al. The tumor microenvironment shows a hierarchy of cell-cell interactions dominated by fibroblasts. Nat. Commun. 2023, 14, 5810.
- Hu, D.; Li, Z.; Zheng, B.; Lin, X.; Pan, Y.; Gong, P.; Zhuo, W.; Hu, Y.; Chen, C.; Chen, L.; et al. Cancer-associated fibroblasts in breast cancer: Challenges and opportunities. Cancer Commun. 2022, 42, 401–434.
- Takebe, N.; Ivy, P.; Timmer, W.; Khan, M.; Schulz; Harris, P. Cancer-associated Fibroblasts and Therapies That Interfere with Their Activity. Tumor Microenviron. Ther. 2013, 1, 19–36.
- Kareva, I.; Luddy, K.A.; O’Farrelly, C.; Gatenby, R.A.; Brown, J.S. Predator-Prey in Tumor-Immune Interactions: A Wrong Model or Just an Incomplete One? Front. Immunol. 2021, 12, 668221.
- Taylor, T.B.; Wass, A.V.; Johnson, L.J.; Dash, P. Resource competition promotes tumour expansion in experimentally evolved cancer. BMC Evol. Biol. 2017, 17, 268.
- Chang, C.-H.; Qiu, J.; O’Sullivan, D.; Buck, M.D.; Noguchi, T.; Curtis, J.D.; Chen, Q.; Gindin, M.; Gubin, M.M.; van der Windt, G.J.W.; et al. Metabolic Competition in the Tumor Microenvironment Is a Driver of Cancer Progression. Cell 2015, 162, 1229–1241.
- Brown, R.A.M.; Richardson, K.L.; Kabir, T.D.; Trinder, D.; Ganss, R.; Leedman, P.J. Altered Iron Metabolism and Impact in Cancer Biology, Metastasis, and Immunology. Front. Oncol. 2020, 10, 476.
- Bañuls, A.-L.; Thomas, F.; Renaud, F. Of parasites and men. Infect. Genet. Evol. 2013, 20, 61–70.
- Song, X.; Wei, C.; Li, X. The Relationship between Microbial Community and Breast Cancer. Front. Cell Infect. Microbiol. 2022, 12, 849022.
- Liao, W.-T.; Ye, Y.-P.; Deng, Y.-J.; Bian, X.-W.; Ding, Y.-Q. Metastatic cancer stem cells: From the concept to therapeutics. Am. J. Stem Cells 2014, 3, 46–62.
- Baumann, Z.; Auf der Maur, P.; Bentires-Alj, M. Feed-forward loops between metastatic cancer cells and their microenvironment–The stage of escalation. EMBO Mol. Med. 2022, 14, e14283.
- Somveille, M.; Ellis-Soto, D. Linking animal migration and ecosystem processes: Data-driven simulation of propagule dispersal by migratory herbivores. Ecol. Evol. 2022, 12, e9383.
- Wu, J.-S.; Jiang, J.; Chen, B.-J.; Wang, K.; Tang, Y.-L.; Liang, X.-H. Plasticity of cancer cell invasion: Patterns and mechanisms. Transl. Oncol. 2021, 14, 100899.
- Barriere, G.; Fici, P.; Gallerani, G.; Fabbri, F.; Zoli, W.; Rigaud, M. Circulating tumor cells and epithelial, mesenchymal and stemness markers: Characterization of cell subpopulations. Ann. Transl. Med. 2014, 2, 109.
- Liu, F.; Gu, L.-N.; Shan, B.-E.; Geng, C.-Z.; Sang, M.-X. Biomarkers for EMT and MET in breast cancer: An update. Oncol. Lett. 2016, 12, 4869–4876.
- Sciacovelli, M.; Frezza, C. Metabolic reprogramming and epithelial-to-mesenchymal transition in cancer. FEBS J. 2017, 284, 3132–3144.
- Miller, K.M.; Schulze, A.D.; Ginther, N.; Li, S.; Patterson, D.A.; Farrell, A.P.; Hinch, S.G. Salmon spawning migration: Metabolic shifts and environmental triggers. Comp. Biochem. Physiol. Part D Genom. Proteom. 2009, 4, 75–89.
- Wu, Q.; Li, J.; Zhu, S.; Wu, J.; Chen, C.; Liu, Q.; Wei, W.; Zhang, Y.; Sun, S. Breast cancer subtypes predict the preferential site of distant metastases: A SEER based study. Oncotarget 2017, 8, 27990–27996.
- Wu, M.; Liang, Y.; Zhang, X. Changes in Pulmonary Microenvironment Aids Lung Metastasis of Breast Cancer. Front. Oncol. 2022, 12, 860932.
- Lim, S.; Nam, H.; Jeon, J.S. Chemotaxis Model for Breast Cancer Cells Based on Signal/Noise Ratio. Biophys. J. 2018, 115, 2034–2043.
- Farahani, M.K.; Gharibshahian, M.; Rezvani, A.; Vaez, A. Breast cancer brain metastasis: From etiology to state-of-the-art modeling. J. Biol. Eng. 2023, 17, 41.
- Chen, W.; Hoffmann, A.D.; Liu, H.; Liu, X. Organotropism: New insights into molecular mechanisms of breast cancer metastasis. npj Precis. Oncol. 2018, 2, 4.
- Schrijver, W.A.M.E.; Selenica, P.; Lee, J.Y.; Ng, C.K.Y.; Burke, K.A.; Piscuoglio, S.; Berman, S.H.; Reis-Filho, J.S.; Weigelt, B.; van Diest, P.J.; et al. Mutation Profiling of Key Cancer Genes in Primary Breast Cancers and Their Distant Metastases. Cancer Res. 2018, 78, 3112–3121.
- Yumoto, K.; Eber, M.R.; Berry, J.E.; Taichman, R.S.; Shiozawa, Y. Molecular pathways: Niches in metastatic dormancy. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2014, 20, 3384–3389.
- Măgălie, A.; Schwartz, D.A.; Lennon, J.T.; Weitz, J.S. Optimal dormancy strategies in fluctuating environments given delays in phenotypic switching. J. Theor. Biol. 2023, 561, 111413.
- Johnson, R.W.; Sowder, M.E.; Giaccia, A.J. Hypoxia and Bone Metastatic Disease. Curr. Osteoporos. Rep. 2017, 15, 231–238.
- Bushnell, G.G.; Deshmukh, A.P.; den Hollander, P.; Luo, M.; Soundararajan, R.; Jia, D.; Levine, H.; Mani, S.A.; Wicha, M.S. Breast cancer dormancy: Need for clinically relevant models to address current gaps in knowledge. npj Breast Cancer 2021, 7, 66.
- Tivari, S.; Lu, H.; Dasgupta, T.; De Lorenzo, M.S.; Wieder, R. Reawakening of dormant estrogen-dependent human breast cancer cells by bone marrow stroma secretory senescence. Cell Commun. Signal. 2018, 16, 48.
- Bleicher, R.J. Timing and Delays in Breast Cancer Evaluation and Treatment. Ann. Surg. Oncol. 2018, 25, 2829–2838.
- Wu, J.; Liu, H.; Hu, T.; Long, M.; Zhou, X.; Wang, S. The natural history of breast cancer: A chronological analysis of breast cancer progression using data from the SEER database. Ann. Transl. Med. 2022, 10, 365.
- Hu, Z.; Curtis, C. Looking backward in time to define the chronology of metastasis. Nat. Commun. 2020, 11, 3213.
- Menes, T.S.; Terry, M.B.; Goldgar, D.; Andrulis, I.L.; Knight, J.A.; John, E.M.; Liao, Y.; Southey, M.; Miron, A.; Chung, W.; et al. Second primary breast cancer in BRCA1 and BRCA2 mutation carriers: 10-year cumulative incidence in the Breast Cancer Family Registry. Breast Cancer Res. Treat. 2015, 151, 653–660.
- Vannote, R.L.; Minshall, G.W.; Cummins, K.W.; Sedell, J.R.; Gushing, C.E. The River Continuum Concept. Can. J. Fish. Aquat. Sci. 1980, 37, 130–137.
- Doretto, A.; Piano, E.; Larson, C.E. The River Continuum Concept: Lessons from the past and perspectives for the future. Can. J. Fish. Aquat. Sci. 2020, 77, 1853–1864.
- Tinganelli, W.; Durante, M. Tumor Hypoxia and Circulating Tumor Cells. Int. J. Mol. Sci. 2020, 21, 9592.
- Rao, S.M.N.; Tata, U.; Lin, V.K.; Chiao, J.-C. The Migration of Cancer Cells in Gradually Varying Chemical Gradients and Mechanical Constraints. Micromachines 2014, 5, 13–26.
- Garg, A.A.; Jones, T.H.; Moss, S.M.; Mishra, S.; Kaul, K.; Ahirwar, D.K.; Ferree, J.; Kumar, P.; Subramaniam, D.; Ganju, R.K.; et al. Electromagnetic fields alter the motility of metastatic breast cancer cells. Commun. Biol. 2019, 2, 303.
- Varennes, J.; Moon, H.-R.; Saha, S.; Mugler, A.; Han, B. Physical constraints on accuracy and persistence during breast cancer cell chemotaxis. PLoS Comput. Biol. 2019, 15, e1006961.
- Liu, Z.; Lee, S.J.; Park, S.; Konstantopoulos, K.; Glunde, K.; Chen, Y.; Barman, I. Cancer cells display increased migration and deformability in pace with metastatic progression. FASEB J. 2020, 34, 9307–9315.
- Couzin, I.D. Collective animal migration. Curr. Biol. 2018, 28, R976–R980.
- Amintas, S.; Bedel, A.; Moreau-Gaudry, F.; Boutin, J.; Buscail, L.; Merlio, J.-P.; Vendrely, V.; Dabernat, S.; Buscail, E. Circulating Tumor Cell Clusters: United We Stand Divided We Fall. Int. J. Mol. Sci. 2020, 21, 2653.
- Kubelka, V.; Sandercock, B.K.; Székely, T.; Freckleton, R.P. Animal migration to northern latitudes: Environmental changes and increasing threats. Trends Ecol. Evol. 2022, 37, 30–41.
- Rejniak, K.A. Circulating Tumor Cells: When a Solid Tumor Meets a Fluid Microenvironment. In Systems Biology of Tumor Microenvironment; Advances in Experimental Medicine and Biology; Springer: Cham, Switzerland, 2016; Volume 936, pp. 93–106.
- Bulfoni, M.; Turetta, M.; Del Ben, F.; Di Loreto, C.; Beltrami, A.P.; Cesselli, D. Dissecting the Heterogeneity of Circulating Tumor Cells in Metastatic Breast Cancer: Going Far Beyond the Needle in the Haystack. Int. J. Mol. Sci. 2016, 17, 1775.
- Menyailo, M.E.; Tretyakova, M.S.; Denisov, E.V. Heterogeneity of Circulating Tumor Cells in Breast Cancer: Identifying Metastatic Seeds. Int. J. Mol. Sci. 2020, 21, 1696.
- Heitzer, E.; Auer, M.; Gasch, C.; Pichler, M.; Ulz, P.; Hoffmann, E.M.; Lax, S.; Waldispuehl-Geigl, J.; Mauermann, O.; Lackner, C.; et al. Complex Tumor Genomes Inferred from Single Circulating Tumor Cells by Array-CGH and Next-Generation Sequencing. Cancer Res. 2013, 73, 2965–2975.
- Park, H.-A.; Brown, S.R.; Kim, Y. Cellular Mechanisms of Circulating Tumor Cells During Breast Cancer Metastasis. Int. J. Mol. Sci. 2020, 21, 5040.
- Shin, D.; Park, J.; Han, D.; Moon, J.H.; Ryu, H.S.; Kim, Y. Identification of TUBB2A by quantitative proteomic analysis as a novel biomarker for the prediction of distant metastatic breast cancer. Clin. Proteom. 2020, 17, 16.
- Veyssière, H.; Bidet, Y.; Penault-Llorca, F.; Radosevic-Robin, N.; Durando, X. Circulating proteins as predictive and prognostic biomarkers in breast cancer. Clin. Proteom. 2022, 19, 25.
- Kjølle, S.; Finne, K.; Birkeland, E.; Ardawatia, V.; Winge, I.; Aziz, S.; Knutsvik, G.; Wik, E.; Paulo, J.A.; Vethe, H.; et al. Hypoxia induced responses are reflected in the stromal proteome of breast cancer. Nat. Commun. 2023, 14, 3724.
- Yates, L.R.; Gerstung, M.; Knappskog, S.; Desmedt, C.; Gundem, G.; Van Loo, P.; Aas, T.; Alexandrov, L.B.; Larsimont, D.; Davies, H.; et al. Subclonal diversification of primary breast cancer revealed by multiregion sequencing. Nat. Med. 2015, 21, 751–759.
- Fernandez-Garcia, D.; Nteliopoulos, G.; Hastings, R.K.; Rushton, A.; Page, K.; Allsopp, R.C.; Ambasager, B.; Gleason, K.; Guttery, D.S.; Ali, S.; et al. Shallow WGS of individual CTCs identifies actionable targets for informing treatment decisions in metastatic breast cancer. Br. J. Cancer 2022, 127, 1858–1864.
- Bhatia, R.; Chang, J.; Munoz, J.L.; Walker, N.D. Forging New Therapeutic Targets: Efforts of Tumor Derived Exosomes to Prepare the Pre-Metastatic Niche for Cancer Cell Dissemination and Dormancy. Biomedicines 2023, 11, 1614.
- Yuan, X.; Qian, N.; Ling, S.; Li, Y.; Sun, W.; Li, J.; Du, R.; Zhong, G.; Liu, C.; Yu, G.; et al. Breast cancer exosomes contribute to pre-metastatic niche formation and promote bone metastasis of tumor cells. Theranostics 2021, 11, 1429–1445.
- Chen, K.M.; Stephen, J.K.; Raju, U.; Worsham, M.J. Delineating an Epigenetic Continuum for Initiation, Transformation and Progression to Breast Cancer. Cancers 2011, 3, 1580–1592.
- Sanati, S. Morphologic and Molecular Features of Breast Ductal Carcinoma in Situ. Am. J. Pathol. 2019, 189, 946–955.
- Lüönd, F.; Sugiyama, N.; Bill, R.; Bornes, L.; Hager, C.; Tang, F.; Santacroce, N.; Beisel, C.; Ivanek, R.; Bürglin, T.; et al. Distinct contributions of partial and full EMT to breast cancer malignancy. Dev. Cell 2021, 56, 3203–3221.e11.
- Manfioletti, G.; Fedele, M. Epithelial-Mesenchymal Transition (EMT). Int. J. Mol. Sci. 2023, 24, 11386.
- Tomaskovic-Crook, E.; Thompson, E.W.; Thiery, J.P. Epithelial to mesenchymal transition and breast cancer. Breast Cancer Res. 2009, 11, 213.
- Malagoli Tagliazucchi, G.; Wiecek, A.J.; Withnell, E.; Secrier, M. Genomic and microenvironmental heterogeneity shaping epithelial-to-mesenchymal trajectories in cancer. Nat. Commun. 2023, 14, 789.
- McFaline-Figueroa, J.L.; Hill, A.J.; Qiu, X.; Jackson, D.; Shendure, J.; Trapnell, C. A pooled single-cell genetic screen identifies regulatory checkpoints in the continuum of the epithelial-to-mesenchymal transition. Nat. Genet. 2019, 51, 1389–1398.
- Kamboj, V.; Kamboj, N.; Sharma, A. A review on general characteristics, classification and degradation of river systems. In Environmental Degradation: Causes and Remediation Strategies; Agro Environ Media, Agriculture and Environmental Science Academy: Haridwar, India, 2020; pp. 47–62.
- Terceiro, L.E.L.; Edechi, C.A.; Ikeogu, N.M.; Nickel, B.E.; Hombach-Klonisch, S.; Sharif, T.; Leygue, E.; Myal, Y. The Breast Tumor Microenvironment: A Key Player in Metastatic Spread. Cancers 2021, 13, 4798.
More