Antibiotic resistance is an increasing global problem for public health, and focusing on biofilms has provided further insights into resistance evolution in bacteria.poorly treated bacterial infections can lead to failures of different organ systems, particularly in fragile patients, and excessive immune-mediated inflammatory responses during sepsis may occur more frequently. Is it possible that nature can help control AMR diffusion? IAn this review, an anaanalysis of resistance mechanisms is summarized, and an excursus of the different approaches to challenging resistance spread based on natural processes is presented as “lessons from Nature”. On the “host side”, immunotherapy strategies for bacterial infections have a long history before antibiotics, but continuous new inputs through biotechnology advances are enlarging their applications, efficacy, and safety. Antimicrobial peptides and monoclonal antibodies are considered for controlling antibiotic resistance. Understanding the biology of natural predators is providing new, effective, and safe ways to combat resistant bacteria. As natural enemies, bacteriophages were used to treat severe infections before the discovery of antibiotics, marginalized during the antibiotic era, and revitalized upon the diffusion of multi-resistance. Finally, sociopolitical aspects such as education, global action, and climate change control are also considered as important tools for tackling antibiotic resistance from the One Health perspective.
- antibiotic resistance
- biofilm and quorum sensing
- antimicrobial peptides
1. Introduction
2. Antibiotics’ Primary Targets and Resistance Mechanisms
The discovery of penicillin in 1928 represented one of the great milestones in human medicine after centuries of periodically deadly epidemics due to bacterial infections amplified by wars, poverty, and poor hygienic conditions. Over time, antibiotics became very popular for their efficacy, specificity, and relatively low cost. Although some resistance was always possible, antibiotics were considered innocuous for humans and animals because their specific targets are present in bacteria and not in eucaryotes (Figure 1). In light of recent evidence of the great importance that the internal microbiota has for its respective host (animals, humans, and plants) this concept should be revisited, and a rational plan for the usage of antibiotics is important not only to control the resistance of pathogens but also to preserve the “good bacteria” to benefit the ecosystem and individuals’ health.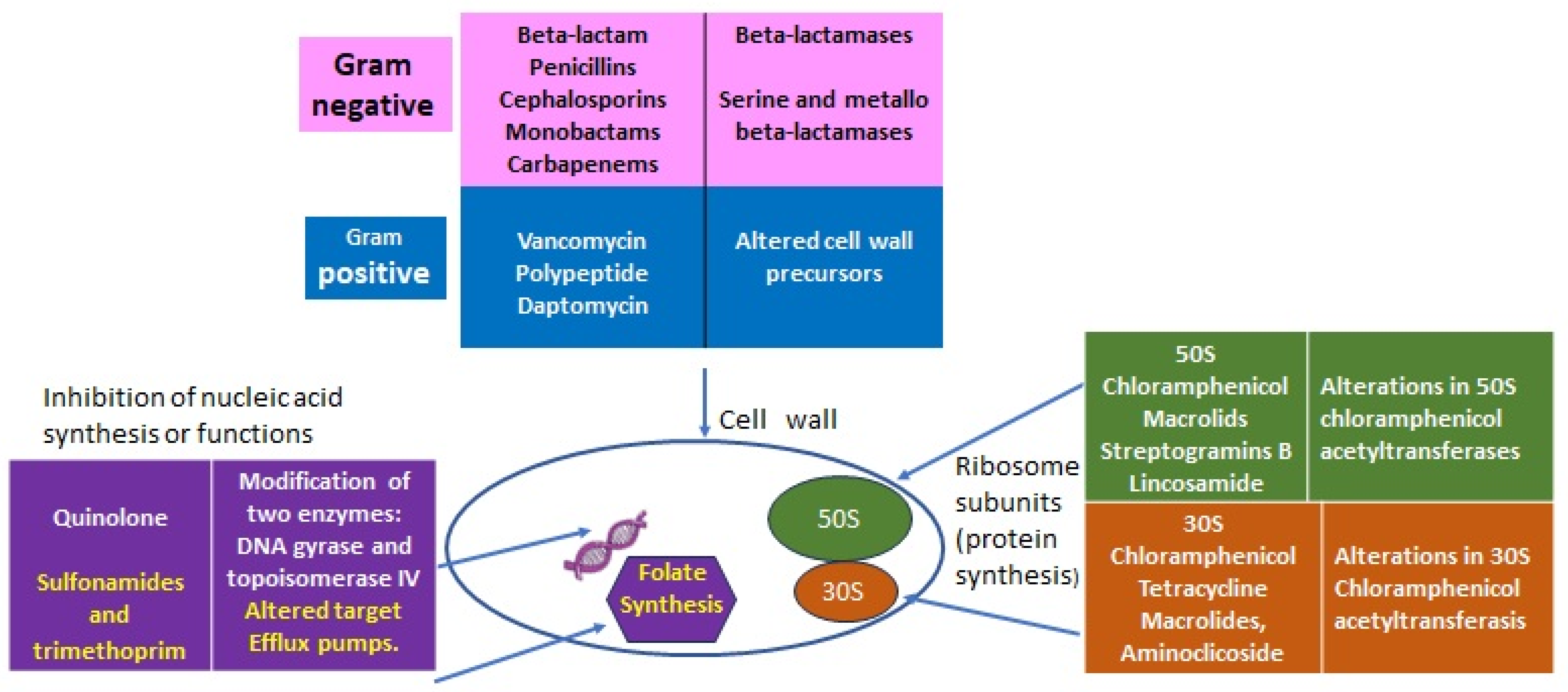
2.1. Lessons from Nature: From Intrinsic to Acquired Resistance
Antibiotic resistance is a natural and general phenomenon in many bacteria which carrying chromosomally located resistance genes (intrinsic resistance) to compete with other species in their environment and to be successful. Highly effective antibiotics have been derived from environmental microorganisms such as Streptomyces, Actinomycetes, and Fungi that produce antibiotics as secondary metabolites. The main molecular mechanisms of bacterial antibiotic resistance can be divided into (I) modifications to the target sites of antibiotics; (II) the alteration or degradation of the antibiotic; (III) antibiotic efflux via transporters; and (IV) reduced antibiotic penetration into bacteria through decreased membrane permeability (Figure 2).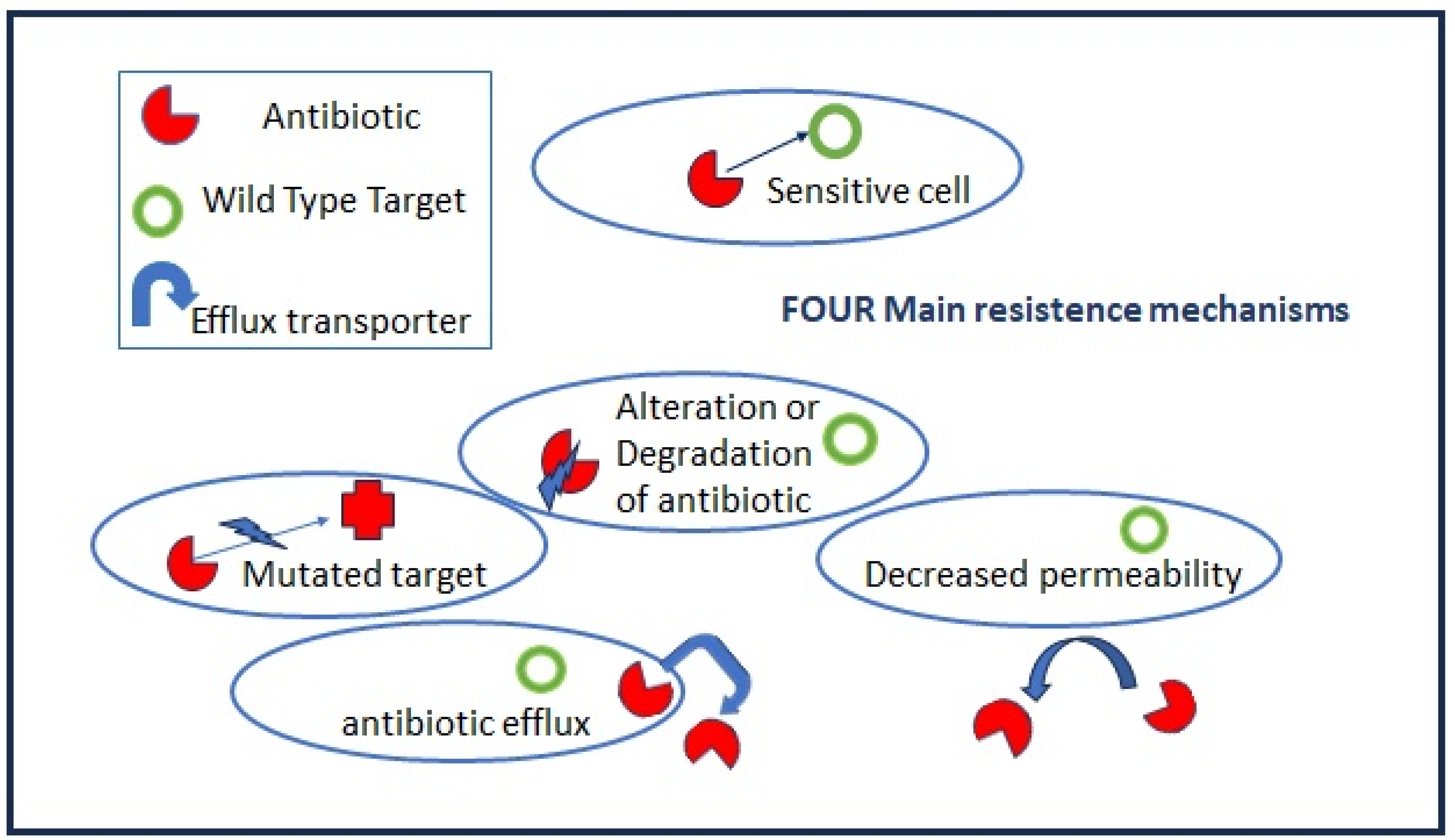
2.1.1. Intrinsic Resistance
All microorganisms are trained through evolution to become the fittest and best adapted to a particular environment, but they are also capable of innovating when alterations to their natural habitats occur [2]. Intrinsic resistance (IR) is an intraspecies genetic and phenotypic resistance which is transmitted vertically and has been used for decades to prepare selective culture media for specific enrichment and to isolate bacteria from complex matrices such as food, soil, and tissue samples. IR is present in the majority of the most relevant opportunistic pathogenic and resistant bacteria in clinical settings, including some bacteria of the so-called ESKAPE group (Enterococcus faecium, Staphylococcus aureus, Klebsiella pneumoniae, Acinetobacter baumannii, Pseudomonas aeruginosa, and Enterobacter species), due to their capacity to escape or evade the action of antimicrobial agents [3]. Moreover, these opportunistic pathogens also have great capacity to acquire new resistance through horizontal transfer and to form biofilms. IR can exist for a single class of antibiotics, such as the resistance of Listeria monocytogenes to cephalosporins [4], but also for multiple drugs, such as the case of Enterococci (Gram-positive, monoderm), which have the potential for resistance to virtually all clinically useful antibiotics [5]. P. aeruginosa, among Gram-negative bacteria (diderm bacteria), represents well the phenomenon of intrinsic bacterial resistance since it carries practically all known mechanisms of antimicrobial resistance: the activation of chromosomal AmpC cephalosporinase; the production of plasmid- or integron-mediated b-lactamases from different molecular classes, diminished outer membrane permeability, and the overexpression of active efflux systems with wide substrate profiles, etc. [6] (Table 1).| Intrinsic Resistance (IR) | Determinants | |
|---|---|---|
| All Gram-negative diderm bacteria | Glycopeptides, lipopeptides, and antibiotic targeting the bacteria peptidoglycan wall | EPS (extra-polymeric substance) avoids the permeability of antibiotics. |
| P. aeruginosa | Sulfonamides, ampicillin, 1st- and 2nd-generation cephalosporins, chloramphenicol, and tetracycline | Constitutive expression of Amp C beta-lactamase and efflux pumps. Low permeability of the outer membrane [6]. |
| Enterococcus spp. | Aminoglycosides, cephalosporins, and lincosamides | Low cell wall permeability, aminoglycoside-modifying enzyme (AME), ribosome-modifying methyltransferase, altered cell wall, and ABC-efflux pump [5]. |
| L. monocytogenes | Cephalosporins | Penicillin-binding proteins, multidrug resistance transporters, cell envelope proteins, etc. [3]. |
| E. coli | Macrolides | Macrolides modifying genes such as mphA; efflux pump [7]. |
| K. pneumonia | Ampicillin | SHV beta-lactamase, the fosfomycin resistance gene fosA, and the nalidixic acid efflux pump OqxAB [8]. |
| A. baumanii | Cephalsporins, ampicillin, glycopeptides, and carbapenems | Class C (AmpC) and Class D beta-lactamases located in chromosome [9]. |
2.1.2. Acquired Resistance
Acquired resistance is related mainly to two mechanisms:- (a)
-
Mutations in target genes located at chromosomal or extrachromosomal elements which are vertically transmitted in the same bacteria species. The mutation rate increases when bacteria are actively multiplying as, for example, during the acute phase of host infection.
- (b)
-
Horizontal gene transfers occur through mobile elements that can be transmitted both intraspecies and among different bacteria genera, i.e., the vancomycin-resistant gene (vanA) from Enterococcus to S. aureus. Plasmids, prophages, pathogenicity islands, restriction and modification systems, transposons, and insertion sequences are able to move within the host genome as well as jump across genomes. Mobile elements can change their insertion location and copy number and produce frequent gene gain and loss, modifying and co-evolving with chromosomal genomes. The genetic modifications induced by mobile elements can deeply affect bacterial fitness, contributing to their adaptation to new environments and, ultimately, producing evolutionarily distinct species over time. Once the acquisition of resistance determinants is established in few strains, antibiotic misuse and pressure drive the positive selection of resistant over sensitive strains.
2.1.3. Acquired Resistance through Bacterial Cooperation
The study of biofilms as sources of persistent infection and contamination underpinned the importance of looking at microorganisms in natural conditions, as members of complex communities which modify their capacities. In contrast to laboratory conditions in which single colonies of the same strain are grown and analyzed in pure cultures, in natural external environments and internally in a host, bacterial cells are “compelled” to interact with many different components [11]. “Social structure” has been a fitness evolutionary advantage for “social animals” including humans passing from nomadism to settled communities. Individuals each gain a different “social role and activity”, and all work together for the wellness of the community. In bacteria, a cooperative resistance process has been observed when resistant bacteria producing beta-lactamases can help antibiotic-sensitive bacteria survive during antibiotic treatment [12]. Cooperative action has been observed in sensitive and resistant mixed cultures of Salmonella tiphymurium strains toward the antibiotic ceftriaxone with an increase in the MIC value upon an increase in cell density [13].
References
- Adriaenssens, N.; Coenen, S.; Versporten, A.; Muller, A.; Minalu, G.; Faes, C.; Vankerckhoven, V.; Aerts, M.; Hens, N.; Molenberghs, G.; et al. European Surveillance of Antimicrobial Consumption (ESAC): Outpatient antibiotic use in Europe (1997–2009). J. Antimicrob. Chemother. 2011, 66, vi3–vi12.
- Youle, M.; Rohwer, F.; Stacy, A.; Whiteley, M.; Steel, B.C.; Delalez, N.J.; Nord, A.L.; Berry, R.M.; Armitage, J.P.; Kamoun, S.; et al. The Microbial Olympics. Nat. Rev. Microbiol. 2012, 10, 583–588.
- World Health Organization. Global Priority List of Antibiotic-Resistant Bacteria to Guide Research, Discovery, and Development of New Antibiotics; World Health Organization: Geneva, Switzerland, 2017; pp. 1–7.
- Krawczyk-Balska, A.; Markiewicz, Z. The intrinsic cephalosporin resistome of Listeria monocytogenes in the context of stress response, gene regulation, pathogenesis and therapeutics. J. Appl. Microbiol. 2016, 120, 251–265.
- Hollenbeck, B.L.; Rice, L.B. Intrinsic and acquired resistance mechanisms in enterococcus. Virulence 2012, 3, 421–569.
- Strateva, T.; Yordanov, D. Pseudomonas aeruginosa—A phenomenon of bacterial resistance. J. Med. Microbiol. 2009, 58 Pt 9, 1133–1148.
- Nguyen, M.C.P.; Woerther, P.L.; Bouvet, M.; Andremont, A.; Leclercq, R.; Canu, A. Escherichia coli as reservoir for macrolide resistance genes. Emerg. Infect. Dis. 2009, 15, 1648–1650.
- Bernardini, A.; Cuesta, T.; Tomás, A.; Bengoechea, J.A.; Martínez, J.L.; Sánchez, M.B. The intrinsic resistome of Klebsiella pneumoniae. Int. J. Antimicrob. Agents 2019, 53, 29–33.
- Kyriakidis, I.; Vasileiou, E.; Pana, Z.D.; Tragiannidis, A. Acinetobacter baumannii Antibiotic Resistance Mechanisms. Pathogens 2021, 10, 373.
- Chambers, H.F.; DeLeo, F.R. Waves of resistance: Staphylococcus aureus in the antibiotic era. Nat. Rev. Microbiol. 2009, 7, 629–641.
- Donlan, R.M. Biofilms: Microbial life on surfaces. Emerg. Infect. Dis. 2002, 8, 881–890.
- Yurtsev, E.A.; Conwill, A.; Gore, J. Oscillatory dynamics in a bacterial cross-protection mutualism. Proc. Natl. Acad. Sci. USA 2016, 113, 6236–6241.
- Dawan, J.; Ahn, J. Assessment of cooperative antibiotic resistance of Salmonella Typhimurium within heterogeneous population. Microb. Pathog. 2021, 157, 104973.