1. Introduction
The respiratory system is also subject to circadian rhythm regulation by the suprachiasmatic nucleus (SCN), mainly mediated by the vagus nerve
[1][43]. In addition to this central regulation, a “pulmonary clock” was also described, by examining the expression of clock genes in multiple tissues in mice and humans
[1][2][43,44]. Disturbances in the circadian clock during chronic lung diseases could play a role in increased oxidative stress, the inflammatory response, metabolic imbalances, hypoxia/hyperoxia, mucus secretion, autophagy, and lung function disorders
[3][17]. Proper synchronization and maintenance of the circadian rhythm are essential for normal lung function. An alteration in the circadian clock, due to genetic or environmental factors, produces stress
[4][19].
2. Asthma
Asthma is a heterogeneous disease characterized by chronic inflammation and airway hyperresponsiveness and remodeling resulting in a wide variety of clinical presentations
[5][49]. Circadian rhythm disruption seems to influence asthma development and symptomology. The pulmonary function seems to be subject to day–night fluctuations, with an increase in airway resistance during sleep at night
[6][50]. Some patients present nocturnal worsening of symptoms
[7][51]. Multiple factors could contribute to this rhythmic worsening, including an increase in peak serum melatonin levels
[8][52], the supine position, which can induce airflow obstruction
[9][53], and air temperature cooling
[10][54], among others. Night workers also seem to have an increased risk of developing asthma
[11][55]. The inflammatory state also seems to be influenced by the circadian clock. Cortisol, an important immunosuppressor and anti-inflammatory hormone, shows daily fluctuations in its circulating levels, and its nadir (time point characterized by the lowest concentration) is reached at midnight. Moreover, children with asthma have lower circulating cortisol levels
[12][56], and disturbances in the fluctuations of cortisol have been associated with nocturnal asthma severity
[3][13][17,57].
Alterations in clock genes are associated with asthma severity. Asthmatic patients showed a significant downregulation of these genes
[14][15][58,59]; specifically, PER3 levels appeared diminished in nocturnal asthma
[15][59], and BMAL1 knock-out mice developed an asthmatic phenotype after viral exposure
[14][58]. Interestingly, inhibition of the BMAL1/FOXA2 signaling pathway produced increases in nocturnal IL-6 levels in respiratory tract epithelial cells
[16][60]. Furthermore, the induction of asthma was associated with a marked increase in lung inflammation in mice lacking BMAL1 expression in myeloid cells, increasing the number of eosinophils and IL-5 serum levels (
Figure 1)
[17][61].
3. Chronic Obstructive Pulmonary Disease
Chronic obstructive pulmonary disease (COPD), a progressive chronic respiratory disorder characterized by an obstructive ventilatory pattern
[18][62], which unlike asthma is rarely reversible, is very often related to smoking or exposure to pollutants as well as to accelerated lung aging, leading to chronic respiratory failure
[18][19][62,63]. Interestingly, COPD seems to follow a diurnal pattern, with patients reporting a substantial worsening of symptoms in the morning
[18][62]. This could be related to the changes in peak expiratory flow (PEF), which is subject to circadian variations, peaking in the afternoon
[20][21][64,65], while the maximum volume of air exhaled in the first second (FEV 1) is the lowest during the morning hours
[22][66]. Moreover, in murine models, cigarette smoke (CS) was shown to reduce the activity of sirtuin 1 deacetylase (SIRT1), a marker of aging and inflammation, which regulates BMAL and PER2 through acetylation (
Figure 1)
[19][23][24][63,67,68].
The molecular clock seems to have a mutual relation with the inflammatory response and is altered in pathological states such as COPD. In human airway epithelial cells and lung tissue of REV-ERBα KO mice, CS exposure produced a heightened inflammatory response, with an increase in neutrophils and the release of proinflammatory cytokines (IL-6, MCP-1, and KC), in addition to the expression of p16 (a pro-senescence marker) and of markers of chronic lung remodeling, as observed in emphysema/COPD
[25][26][69,70]. REV-ERBα is a nuclear heme receptor, a transcriptional repressor (binding and interacting with HDAC3 and NcoR), and a critical component of the molecular clock driving the daily rhythmicity of metabolism and inflammatory and immune responses
[25][69]. In contrast to the antagonistic regulator REV-ERB, the retinoic acid-like orphan receptor alpha/gamma (RORα/γ) acts as an activator, and these molecules together are considered a “stabilizing loop” of the molecular clock
[25][69]. Moreover, RORα levels are elevated in the lungs of COPD patients, but interestingly, RORα-deficient mice are protected against emphysema induced by elastase and CS exposure
[27][71]. Interestingly, RORα responds to DNA damage, and a decrease in this protein is associated with low levels of apoptosis
[23][67]. Interestingly, BMAL1 has also been shown to be underexpressed in human bronchial epithelial cells, lung tissue, peripheral blood mononuclear cells (PBMCs), sputum cell from COPD patients, and healthy cells exposed to CS
[24][28][29][68,72,73]. The BMAL1 and CLOCK proteins seem to play an integral role in regulating cellular senescence, since the decrease in their expression by CS exposure upregulates senescence mediated by the MAPK pathways, a process implicated in the development of COPD (
Figure 1)
[29][73].
4. Lung Cancer
Lung cancer is the most commonly diagnosed cancer type in both sexes combined (11.6%) and the leading cause of cancer death among men
[30][74]. Smoking has a causal link with the development of lung cancer, as shown by extensive epidemiologic research evidence
[31][32][33][75,76,77]. In recent years, the relationship between sleep (duration, quality, and timing) and cancer risk has been consolidated particularly in regard to the development of lung cancer
[34][35][36][37][78,79,80,81]. For instance, insomnia was causally associated with an increased risk of lung adenocarcinoma, while sleep with typical duration showed a protective effect on the risk of lung adenocarcinoma
[38][39][82,83]. Also, clock genes, primarily BMAL1, have been implicated in tumorigenesis
[40][41][84,85].
Clock genes’ behavior seems to depend on the type of lung cancer. In a recent study, the overexpression of CRY2, BMAL1, and RORα and the underexpression of Timeless and NPAS2 were associated with a favorable prognosis of lung adenocarcinoma. On the other hand, heightened expression of Sharp2 was observed in squamous cell carcinoma and was associated with poor survival. In the case of non-small cell lung cancer (NSCLC), clock genes establish a cancer circadian rhythm, asynchronic with respect to that of healthy tissues
[42][86]. Moreover, CS exposure decreased Nr1d1 expression in both tumor-resistant and tumor-susceptible mice, making them more prone to developing CS-related lung cancer (
Figure 1)
[43][87].
The circadian rhythm also seems to regulate the expression of tumor suppressor genes and proto-oncogenes, such as p53, a critical tumor suppressor protein involved in the DNA damage response and apoptosis, whose transcription, stability, and activity are modulated by BMAL1 and PER2
[44][45][46][88,89,90]. While BMAL1 binds to the promoter region of p53, PER2 binds to the p53 protein, providing stabilization and nuclear translocation for its activation, in the presence of DNA damage in cells
[44][45][46][47][48][49][88,89,90,91,92,93]. Furthermore, BMAL1 suppresses cell invasion by blocking the PI3K-AKT-MMP-2 pathway, as seen in BMAL1 KO human lung cancer and glioma cell lines
[50][94]. The PI3K-AKT-MMP-2 pathway is involved in a myriad of cellular processes that can be altered in cancerous environments, including cellular apoptosis, migration/invasion, cell growth, and angiogenesis, among others, through the regulation of FOXO, mTORC1/2, XIAP, Bcl2, and MDM2, the last of which inhibits p53 activity (
Figure 1)
[51][95].
Melatonin is also an important regulator of oncogenic processes, especially those induced by hypoxia, and could serve as a potential therapeutic agent. Among the key anti-cancer mechanisms of melatonin are the inhibition of hypoxia-induced proliferation in cancer cells, the induction of apoptosis, anti-angiogenesis mechanisms, the inhibition of HIFs, the reduction in hypoxia-induced reactive oxygen species (ROS), the inhibition of cancer cell migration and invasion, the attenuation of epithelial–mesenchymal transition (EMT), the inhibition of endoplasmic reticulum stress, the reversion of altered states of metabolism (e.g., the Warburg effect), and the improvement of chemoradiotherapy sensitivity
[52][53][54][96,97,98].
NSCLC is associated with an overactivation of AKT
[55][99]. In a study with 1144 patients with such a disease, 3.7% of the patients were identified to have PIK3CA mutations in exons 9 and 20, and up to 57.1% of the patients showed additional aberrations in oncogenic drivers
[56][100]. Furthermore, the gene ARNTL (that encodes the BMAL protein) was found to protect against lung adenocarcinoma growth, and the ARNTL gene was shown to increase the expression of the circular RNA circGUCY1A2 and activate the miR-200c-3p/PTEN axis, initiating a tumor suppressive activity
[57][101].
The proteins differentiated embryonic chondrocyte expressed gene 1 (DEC1; BHLHE40/Stra13/Sharp2) and 2 (DEC2; BHLHE41/Sharp1), related to the circadian genes, can act as repressors and coactivators of BMAL1 and CLOCK or NPAS
[58][102]. These two proteins are implicated in regulating cell differentiation, circadian rhythms, apoptosis, the response to hypoxia, EMT, and carcinogenesis
[59][60][103,104]. As stated, HIFs are activated during hypoxia and are described to participate in different aspects of cancer pathophysiology. Interestingly, Sharp1 interacts with HIF-1α, driving it to proteosome degradation and preventing the formation of the HIF-1α/1β heterodimer that promotes metastatic gene expression
[61][105]. Sharp1 suppresses metastasis in various types of tumors including breast, endometrial, prostate, thyroid, and lung cancers
[62][63][64][65][66][106,107,108,109,110]. Paradoxically, in human breast cancer cells, HIF-2α was shown to increase Sharp1 expression through the activation of AKT in a hypoxic state, which in turn promoted c-Myc expression and favored cellular proliferation (
Figure 1)
[60][104].
Concerning the antitumor effects, the overexpression of Sharp1 inhibited the formation of colonies of lung cancer cells (A549, NCI-H520, and NCI-H596 cells) by promoting a decrease in cyclin D1 (CCND1) expression
[65][109]. In this sense, Sharp1 and Sharp2 seem to have opposing roles in cancer progression. While Sharp1 was shown to suppress EMT and metastasis by attenuating NOTCH1 signaling in endometrial cancer cells
[67][111] and TGF-β-associated EMT in prostate cancer PC-3 cells
[66][110], Sharp2 favored TGF-β-associated EMT
[59][103] and interplayed with NOTCH1 to favor cell growth and invasiveness
[66][110]. Furthermore, in fibroblasts, Sharp1 was activated during DNA damage and was involved in cell cycle arrest and apoptosis; an aberrant expression of Sharp1 could thus participate in tumorigenesis
[68][112]. Unexpectedly, in various cancer cell lines, the hypoxia-induced expression of both Sharp1 and Sharp2 downregulated the MLH1 protein, an essential component of the DNA mismatch repair (MMR) system
[69][113]. However, as stated, Sharp1/2 can have both repressor and oncogenic activity, and their function seems to vary by tissue type
[70][114].
5. Idiopathic Pulmonary Fibrosis
Idiopathic pulmonary fibrosis (IPF) is a chronic, progressive lung disease primarily affecting the elderly, characterized by irreversible damage and an unknown etiology, which offers limited treatment options. Typically, patients diagnosed with IPF face a grim prognosis, succumbing to respiratory failure within two to five years
[71][72][73][115,116,117]. The pathogenesis of IPF involves the activation of epithelial cells in response to various injuries, leading to the release of mediators that contribute to fibrosis. These mediators include cytokines and growth factors, notably, platelet-derived growth factor (PDGF), transforming growth factor β (TGF-β), tumor necrosis factor α (TNFα), endothelin-1 (ET-1), and connective tissue growth factor (CTGF)
[74][118]. Particularly, TGF-β1 plays a central role in fibrotic tissue formation within the lungs, which is a key driver of IPF development
[75][119]. TGF-β1 promotes fibroblast-to-myofibroblast differentiation, EMT, and the production of matrix metalloproteinases, all contributing to the formation of fibroblast foci
[76][120]. These foci generate excessive extracellular matrix deposition, predominantly, collagen deposition, leading to lung architectural scarring and destruction
[74][77][78][79][80][118,121,122,123,124]. Circadian oscillations within the lung epithelium were associated with inflammatory responses in IPF patients
[81][125]. These oscillations are implicated in the remodeling of the lung’s extracellular matrix, largely due to increased levels of the REV-ERBα protein, a negative regulator of BMAL1, which itself contributes to the regulation of inflammatory mediators
[82][126].
A reduced expression in the BHLHE41 gene
[83][134] was observed in hypoxic lung fibroblasts obtained from IPF individuals. As mentioned earlier, the protein encoded by this gene is recognized as a tumor suppressor. Consequently, the absence of its expression in the lung fibroblasts of IPF patients led us to hypothesize that BHLHE41 may play a significant role in fibrosis progression. Such a hypothesis gains credence considering the observation that TGF-β induces the upregulation of BHLHE40 and the downregulation of BHLHE41 in prostate cancer cells (PC-3)
[59][103]. It is worth noting that TGF-β triggers the phosphorylation of Smad2, subsequently leading to the activation of mesenchymal markers such as N-cadherin and vimentin, while concurrently suppressing epithelial markers like E-cadherin
[59][103]. This suggests that TGF-β may initiate EMT, a process known to be involved in developing pulmonary fibrosis. Furthermore, evidence indicates that the two genes BHLHE40 and BHLHE41 may have opposing effects in the context of cancer development.
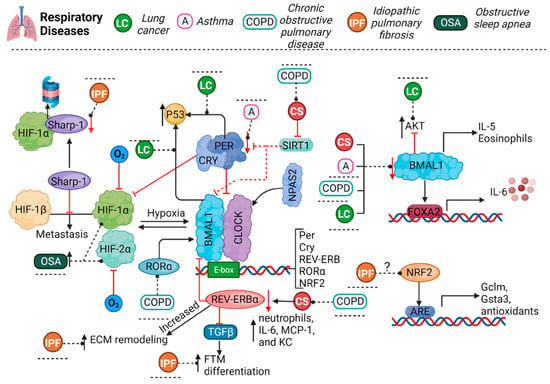
Figure 1. The molecular circadian clock involves an autoregulatory loop encompassing the transcription activators BMAL1, CLOCK and/or NPAS2, regulated by the repressors Per and CRY. During hypoxia, HIF-1α and HIF-2α activation increases. These molecules have a dual regulation: HIF-1α and BMAL1 bind together to clock gene promoters, while clock proteins bind to the promoter of HIF-1α. Per3 levels were found to be diminished in asthma. BMAL1 levels seem to be diminished in animal models or patients with asthma, COPD, or LC and are also diminished by CS exposure; inhibition of or decrease in BMAL1 can lead to an increase in IL-5 and IL-6 expression and in the number of eosinophils in the presence of asthma. COPD or CS exposure reduces SIRT1 expression, which regulates BMAL1 and PER2. Moreover, CS exposure in REV-ERBα KO mice increased the number of neutrophils and the expression of proinflammatory cytokines. In COPD, the levels of RORα, the counterpart to REV-ERBα and a regulator of BMAL1, are diminished. BMAL1 also regulates p53 expression, and PER binds to p53, regulating the stabilization and nuclear translocation of the protein. BMAL1 can also block the AKT signaling pathway, which is overactivated in lung cancer. Furthermore, the protein Sharp1 was shown to interact with HIF-1α to induce its proteasomal degradation and prevent its union with HIF-1β, which promotes the expression of metastatic genes. In IPF, the increment in REV-ERBα leads to ECM remodeling, and REV-ERBα suppression promotes fibroblast-to-myofibroblast differentiation. NRF2 is positively regulated by BMAL1-CLOCK, and NRF2 increases the expression of Gclm and Gsta3, proteins involved in glutathione metabolism. Additionally, Sharp1 expression is reduced in IPF. In OSA, HIFs are overactivated and increase the expression of Clock, BMAL1, and CRY2. A, asthma; COPD, chronic obstructive pulmonary disease; LC, lung cancer; IPF, idiopathic pulmonary disease; OSA, obstructive sleep apnea; O2, oxygen; BMAL1, brain and muscle ARNT-like 1; NPAS2, neuronal PAS domain protein 2; CLOCK, circadian locomotor output cycles kaput; PER, period; CRY, cryptochrome; SIRT1, sirtuin1; HIF, hypoxia-inducible factor; IL-5, interleukin 5; IL-6, interleukin 6; MCP-1, monocyte chemoattractant protein 1; KC, keratinocyte chemoattractant; AKT, protein kinase B; ECM, extracellular matrix; NRF2, nuclear factor erythroid 2-related factor 2; ARE, antioxidant response element.
6. Obstructive Sleep Apnea
Obstructive sleep apnea (OSA) is the most common type of sleep-related breathing disorder, characterized by repeated episodes of complete or partial airway collapse in association with a decrease in oxygen saturation or with arousal from sleep. These episodes lead to fragmented and nonrestorative sleep that, over time, impact negatively on cardiovascular health, cognitive performance, and quality of life
[84][85][135,136]. Apnea is defined as an interruption of airflow for at least ten seconds, resulting in a complete lack of breathing; these events are obstructive, central, or mixed. Obstructive apneas occur when the airway is blocked, but respiratory effort is present in the thorax and abdomen. In central apneas, there is no respiratory effort or airflow, indicating that the brain is not sending signals to stimulate breathing. Mixed apneas are a combination of these two dysfunctions
[85][86][87][88][136,137,138,139]. The standard diagnosis for OSA involves nocturnal polysomnography, which records the number of apneas and hypopneas per hour of effective sleep. The Apnea–Hypopnea Index (AHI) is used to assess the severity of the condition
[89][90][91][92][140,141,142,143]. The main symptoms of OSA include loud snoring, nighttime awakenings, daytime sleepiness, and fatigue
[84][92][135,143]. Possible additional signs include morning headaches, memory and concentration problems, night sweats, and wheezing
[85][136]. OSA patients often experience cycles of oxygen desaturation followed by reoxygenation during sleep
[93][94][144,145]. States of hypoxia, such as the prolonged period of hypobaric hypoxia present during long-duration flights, are shown to disrupt multiple processes regulated by the circadian rhythm, including the maintenance of core body temperature
[95][146], melatonin secretion
[96][147], and the cortisol secretion cycle
[97][148]. These changes significantly modify sleep (latency, time, and quality of sleep) and can be influenced by individual characteristics such as age, physical fitness, and the sympathetic reaction to hypoxia.
To provide further support for this association, bidirectional interactions between HIF-1α and the circadian clock have been demonstrated. For instance, BMAL1 and CLOCK form heterodimers and regulate the rhythmic expression of HIF-1α
[98][41]. Moreover, in a hypoxic environment, alterations in the expression of circadian genes were observed. Overexpression of HIF-1α and HIF-2α was shown to increase the gene expression of CLOCK, BMAL1, and CRY2, while decreasing the expression of PER1, PER2, PER3, CRY1, and CKIε
[99][149]. In a separate study involving individuals with obstructive sleep apnea (OSA), it was noted that the transcription of BMAL1, CLOCK, and CRY2 was irregular, and the levels of CRY1 and PER3 significantly decreased at midnight. This suggests that the latter two genes could potentially serve as prognostic markers for OSA severity (
Figure 1)
[100][150]. Although the role of circadian gene polymorphisms in respiratory pathologies is relatively unexplored, recent findings suggest their potential significance. For instance, a study revealed that mutations in the BHLHE41 gene were associated with reduced total sleep
[101][151].
7. Influenza and COVID-19
Influenza, a highly contagious and often fatal disease, claims nearly half a million lives annually worldwide. Until now, four distinct types of influenza viruses have been recognized: A, B, C, and D. Only types A and C viruses are recognized to infect humans. Types A and B viruses co-circulate as the primary seasonal strains, giving rise to a spectrum of respiratory infections and related complications, ranging from mild to severe, in human populations. The typical influenza symptoms encompass fever, fatigue, cough, and body aches
[102][103][104][105][152,153,154,155]. Almost 10% of the global population falls prey to influenza each year
[102][104][152,154], since this virus incubates for a period ranging from 1 to 4 days
[106][156].
Lung damage inflicted by influenza is associated with disruptions in the circadian clock, compromised lung function, and decreased survival rates
[107][157]. The protein BMAL1 plays a pivotal role in alveolar epithelial cells (AECs), with specific BMAL1 deletion disrupting lung neutrophil infiltration, biomechanical functions, and the response to influenza infection
[108][158]. Conversely, the genetic deletion of BMAL1 and the overexpression or activation of REV-ERB using synthetic agonists inhibit the replication of hepatitis C virus (HCV) and related flaviviruses like dengue and Zika. The overexpression of REV-ERB denies HCV cell entry, viral RNA replication, and the release of infectious particles through lipid signaling pathway disruption
[109][159]. REV-ERB agonists inhibit HIV transcription and replication in vivo in primary cell lines
[110][160]. Together, these findings indicate the important role of circadian genes in viral infections and their influence on immune responses.
In recent times, the world endured one of its most devastating pandemics, resulting in the loss of millions of lives due to a Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) outbreak, responsible for Coronavirus Disease 2019 (COVID-19)
[111][112][161,162]. During the COVID-19 pandemic, two factors significantly impacted sleep quality: irregular schedules and confinement
[113][114][163,164]. Such disruptions altered the daily routines, including office and work schedules, social engagements, and recreational activities
[113][163]. It is well documented that even a single night of sleep deprivation can lead to mood swings and a weakened immune system
[113][115][163,165]. This connection could result in disturbances in mitochondrial metabolism and influence the normal functions of the immune response
[116][166]. Interestingly, several viruses inhibit mitochondrial melatonin production
[116][166]; notably, the loss of circadian activation coincides with suppressed REV-ERBα expression, indicating a possible link between clock genes and inflammatory pathways
[117][167].
The circadian clock influences both innate and adaptive immune systems, impacting processes from leukocyte mobilization and chemotaxis to cytokine release and T-cell differentiation
[118][168]. The pathogenicity of viral infections can be influenced by the host’s circadian clock through two primary mechanisms: (1) a direct regulation of viral replication within the target cells; (2) indirect effects on innate and adaptive immune responses
[119][120][121][169,170,171]. Intriguingly, recent studies concluded that the circadian clock, via REV-ERBα and BMAL1, significantly influences the airway inflammatory responses
[81][122][125,172]. Circadian cycle genes regulate cytokines, including IL-6, a function that exhibits diurnal variation and has implications in inflammatory processes, as seen in patients with rheumatoid arthritis
[123][173]. Such a relationship could also shed light on pharmacological treatments variability; for instance, the effectiveness of glucocorticoids depends on the intact functioning of the circadian clock in the airways
[81][125]. Therefore, viral infections, inflammatory responses, and circadian clock genes appear to share intricate pathways, and comprehending these connections may enhance
theour understanding of the pathogenesis of such infectious diseases.