Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Haibo Yuan and Version 2 by Rita Xu.
Vitamin C, a water-soluble vitamin with strong reducing power, cannot be synthesized by the human body and participates in a variety of important biochemical reactions. Vitamin C is widely used in the pharmaceutical, food, health care, beverage, cosmetics, and feed industries, with a huge market demand. The classical two-step fermentation method is the mainstream technology for vitamin C production. D-sorbitol is transformed into L-sorbose by Gluconobacter oxydans in the first step of fermentation; then, L-sorbose is transformed into 2-keto-L-gulonic acid (2-KGA) by a coculture system composed of Ketogulonicigenium vulgare and associated bacteria; and finally, 2-KGA is transformed into vitamin C through chemical transformation.
- Vitamin C
- 2-keto-L-gulonic acid
- mixed fermentation
1. Introduction
Vitamin C, a water-soluble vitamin of vital importance to the human body, participates in many important biochemical reactions in organisms and contains an unsaturated enediol structure, thus having a strong reducing ability. In addition to its antioxidant function, vitamin C is involved in collagen synthesis, hormone synthesis, carnitine synthesis, gene transcription, regulation of translation, L-tyrosine catabolism, and iron absorption in humans [1]. As primates, guinea pigs, and fish lack gulonolactone oxidase (GULO), the enzyme required for vitamin C synthesis, they rely on exogenous vitamin C intake [2]. Vitamin C is widely used in the pharmaceutical, food, health, beverage, cosmetic, and feed industries [3] and has a huge market demand.
At present, vitamin C is primarily produced through the classical two-step fermentation process (Figure 1). Specifically, in the first step of fermentation, Gluconobacter oxydans converts D-sorbitol into L-sorbose, which is a fast and efficient process. By adding 300 g/L D-sorbitol in batch feeding, the yield of L-sorbose reaches 279.7 g/L at 16 h [4]. Then, in the second fermentation step, L-sorbose is converted into 2-keto-L-gulonic acid (2-KGA) through a coculture system composed of Ketogulonicigenium vulgare and associated bacteria. Subsequently, 2-KGA is converted to vitamin C by esterification and lactonization via chemical catalysis. Among these reactions, the transformation of L-sorbose to 2-KGA takes a long time, with a low yield and conversion rate (Table 1), so it is the committed step that determines the yield and cost of vitamin C.
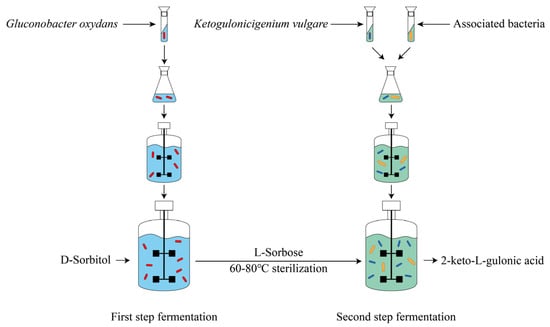
Figure 1. The process of producing the vitamin C precursor 2-keto-L-gulonic acid by two-step fermentation.
Table 1. Fermentation yield of the coculture of K. vulgare and different associated bacteria.
| Associated Bacteria | Fermentation Container | Time (h) |
L-Sorbose Concentration (g/L) | 2-KGA Concentration (g/L) |
Conversion Rate (%) |
References |
|---|---|---|---|---|---|---|
| Xanthomonas maltophilia IFO12692 | 3 L fermentor | 60 | 126 | 124.0 | - | [5] |
| Bacillus cereus 112 | Flask | 45 | 85 | 63.4 | - | [6] |
| Bacillus megaterium 116 | Flask | 45 | 85 | 64.5 | - | [6] |
| Bacillus megaterium 116 and Bacillus cereus 112 (1:3, v/v) | Flask | 45 | 85 | 69.0 | - | [6] |
| Bacillus cereus HB601 | Flask | 96 | 80 | - | 93.0 | [7] |
| Bacillus thuringiensis 320 | 260 m3 fermentor | 48 | 88–92 | 90.2 | 94.5 | [8] |
| Bacillus endophyticus Hbe603 | Flask | 72 | - | 70.0 | 93.0 | [9] |
| Bacillus subtilis A9 | Flask | 48 | 92.5 | 71.2 | - | [10] |
| Bacillus cereus 112 | Flask | 55 | 110 | 98.5 | 89.5 | [11] |
| Bacillus pumilus SH-B9 | Flask | 72 | 80 | 63.1 | - | [12] |
| Saccharomyces cerevisiae VTC2 | Flask | - | 20 | 13.2 | - | [13] |
Although the two-step fermentation method has advantages such as mature technology and high yield, it suffers from shortcomings such as high energy consumption, large equipment investment, and complex operation brought about by the two sterilization and fermentation steps. Therefore, extensive research has been conducted on one-step fermentation, and some researchers have achieved high 2-KGA yields with such an approach. For example, sorbose dehydrogenase (SDH) and sorbosone dehydrogenase (SNDH) of G. oxydans T100 were coexpressed in G. oxydans G624, which was a chemical mutation to inhibit the L-idonate pathway and the replacement of the original promoter with that of Escherichia coli tufB, resulting in a 2-KGA yield of 130 g/L from 150 g/L of D-sorbitol [14]. Wang et al. [15] deleted the gene involved in the L-sorbose metabolism of G. oxydans and cocultured G. oxydans with K. vulgare, obtaining a 2-KGA yield of 76.6 g/L within 36 h. Zhou et al. used the Gluconobacter oxydans ATCC9937 to direct the production of 2-KGA from D-glucose by balancing intracellular and extracellular D-glucose metabolic flux. The 2-KGA titer reached 30.5 g/L [16]. Despite some progress, the developed one-step fermentation method is not yet suitable for industrial production because of the low yield and low conversion rate.
The coculture process has been widely applied in wastewater treatment, biodegradation of textile azo dyes, treatment of contaminated soil, production of biofuels, and various bulk chemicals and natural products, with high application value [17]. Compared to monoculture, coculture systems can perform more complex tasks and have higher stability and robustness to environmental disturbances [17][18][17,18]. In addition, the coculture system can alleviate the growth damage or poor biosynthetic behavior of a single strain caused by excessive cell resource consumption and heavy metabolic burden through division of labor strategies [19]. The coculture system not only makes tasks that cannot be completed by a single strain possible, such as the coculture of Clostridium thermocellum to efficiently hydrolyze cellulose and Thermoanaerobacterium saccharolyticum to produce ethanol from sugars, which produced 38 g/L of ethanol from 92 g/L of avicel [20], but also improves yield and fermentation intensity, such as the coculture of associated bacteria with K. vulgare for 2-KGA production.
In the second step of the two-step fermentation method, K. vulgare, which is responsible for converting L-sorbose into 2-KGA, grows slowly and hardly produces 2-KGA when cultured alone [21][22][21,22]. However, when cocultured with associated bacteria that neither metabolize L-sorbose nor produce 2-KGA, K. vulgare can grow smoothly and efficiently to produce 2-KGA. A variety of associated bacteria, such as Xanthomonas maltophilia [5], Bacillus cereus, Bacillus megaterium [6], Bacillus thuringiensis [8], Bacillus endophyticus [9], Bacillus subtilis [10], Bacillus pumilus [12], and Saccharomyces cerevisiae [13], have previously been studied and exploited. Among them, B. megaterium is the most popular one for industrial production.
Stable interactions in microbial consortia generally rely on communication between cells through the coutilization of different substrates in the environment, the sequential conversion and reutilization of substrates, the complement of metabolites, and other ways to meet the normal growth of a single cell in a multicellular system [23]. However, in the second step of the two-step fermentation process for vitamin C production, what mechanism does the associated bacteria use to promote the growth of K. vulgare and the production of 2-KGA? This has always been a research hotspot, and researchers have conducted extensive research and in-depth exploration.