Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 2 by Camila Xu and Version 1 by Vasile Valeriu Lupu.
Childhood obesity, a real pandemic, represents, in fact, a major public health problem worldwide, with an increasing trend in prevalence during the last decades, especially in developed countries. Due to its associated short- and long-term complications, it carries an important burden not only for health services, but also for society, since it is associated with both psychosocial problems like bullying, resulting in school absences and consequent poor school results, and organic conditions such as metabolic syndrome with insulin resistance, cardiovascular disorders, hypertension, dyslipidemia, non-alcoholic fatty liver disease, obstructive sleep apnea, and even neoplasia.
- prebiotics
- probiotics
- synbiotics
- child
- obesity
- gut microbiota
1. Introduction
Childhood obesity, a real pandemic, represents, in fact, a major public health problem worldwide, with an increasing trend in prevalence during the last decades, especially in developed countries [1,2,3,4,5][1][2][3][4][5]. Due to its associated short- and long-term complications, it carries an important burden not only for health services, but also for society, since it is associated with both psychosocial problems like bullying, resulting in school absences and consequent poor school results, and organic conditions such as metabolic syndrome with insulin resistance, cardiovascular disorders, hypertension, dyslipidemia, non-alcoholic fatty liver disease, obstructive sleep apnea, and even neoplasia [6,7,8,9,10,11,12,13,14][6][7][8][9][10][11][12][13][14]. According to the most recent reports of the World Health Organization (2020), 39 million children under the age of 5 years suffer from either overweight or obesity, with an impressive increase between 5 and 19 years, reaching up to 340 million cases [15]. Over the years, several studies focused on assessing the factors involved in the determinism of obesity, underlying a complex interplay between genetic background, and maternal and obesogenic factors such as maternal diet, eating habits, urbanization, or a sedentary lifestyle, but were not able to precisely state the role or the exact contribution of each of them in the complicated puzzle of obesity [16,17,18,19][16][17][18][19].
It is well-documented that, aside from bacteria, which are the main community harbored by the human intestine, the human intestine is also colonized by viruses, protozoa, and archae [20,21][20][21]. Although several studies proved that microbial DNA is present in samples of amniotic fluid, placenta, and meconium, suggesting that the colonization process might begin in utero, no unanimous consensus was reached yet in the global scientific community [22,23][22][23]. Thus, studies that assessed the role of the placental microbial composition, as well as those that compared meconium samples from different newborns, revealed important differences in terms of microbial composition, depending on maternal well-being [24,25,26,27][24][25][26][27]. Nevertheless, it is clearly established that the colonization process ends between 3–5 years of age, when the child’s microbiota resembles to that of an adult [28]. Several external factors were proven to be essential for the maturation of the microbiota composition, such as maternal diet, type of delivery and feeding, and the early use of antibiotics [6,29][6][29]. Thus, in vaginally born infants, studies revealed that facultative anaerobic bacteria, including Escherichia spp. and the Enterobacteriaceae family, along with Lactobacillus, are the main compounders of intestinal microbiota [30,31][30][31]. Other studies pointed out Sneathia, Lactobacillus, and Prevotella as the main genera colonizing the gut of term infants born vaginally [32]. Contrariwise, when born by cesarean section, the infant’s gut microbiome composition resembles that of the mother’s skin and nosocomial surroundings, consisting of lower levels of Escherichia-Shigella, Bifidobacteria, and Bacteroidetes when compared to vaginally born newborns, associated with an increase in Staphylococcus, Corynebacterium, and Propionibacterium [32,33,34,35,36,37][32][33][34][35][36][37]. The findings were correlated with an increased risk of obesity during childhood [38,39][38][39].
In addition, several differences were noticed depending on the type of feeding, consisting in a predominance of Bifidobacteria in breastfed infants and a more diversified microbiota in formula-fed infant with a predominance of Bacteroides, Staphylococci, Clostridia, Enterobacteria, Enterococci, and the genus Atopobium [6]. Human milk oligosaccharides were proven to favor the growth of Bifidobacteria and Bacteroides, acting also as modulators of innate immune responses, anti-inflammatory molecules, and prebiotics [40]. Moreover, maternal health is a crucial modulator of human milk composition with potential negative consequences on the infant’s gut microbiota [33]. Thus, maternal obesity was associated with a decrease in bacterial community diversity, resulting in higher levels of Staphylococcus and decreased levels of Bifidobacterium [41,42][41][42].
Important changes in microbiota composition occur following the introduction of complementary feeding, such as a considerable increase in alpha diversity due to the replacement of Actinobacteria and Proteobacteria with the phyla Bacteroidetes and Firmicutes, along with a significant increase in the production of short-chain fatty acids [43]. Thus, major changes in the gut microbial composition were noticed between 9 and 18 months of age, consisting in an increase of Bacteroides and Clostridium species, along with a decrease of Bifidobacteria, Enterobacteriaceae, and Lactobacili [44]. Gut microbial diversity and composition were proved to differ depending on the type of diet, with the suggestion that a high-fat Westernized diet might be related to an increased risk of developing obesity during childhood due to a decrease in Prevotellaceae [45]. The same study pointed out that, if consuming a carbohydrate-based diet, children experience a decrease in Bacteroideaceae and an increase in Prevotellaceae [45].
2. Obesity, Microbiota, and Prebiotics, Probiotics, and Synbiotics
It is well-known that both childhood obesity and its related metabolic complications are increasing in incidence during the last decades, but, unfortunately, the molecular basis of the relationship between body weight and its metabolic implications is not completely understood. Still, the most commonly reported obesity-associated metabolic disorder is insulin resistance, which, in fact, is the trigger of subsequent type 2 diabetes mellitus and cardiovascular diseases [4,7,8][4][7][8]. Therefore, the implications of pediatric obesity go beyond childhood into adulthood, increasing considerably the health-related costs. Other important long-term complications of obesity consist of chronic inflammation, which was proven to be present also in children [46], dyslipidemia, hypertension, hyperuricemia, obstructive sleep apnea, nonalcoholic fatty liver disease, and even cancer [6]. Although genetic background plays a major role in predisposing individuals to obesity, obesogenic factors such as eating habits, sedentary lifestyle, and urbanizations seem to be the ones that decide, finally, whether an individual will become obese or not [16]. Not all individuals with obesity display the same degree of metabolic-related complications, although these were proven to occur early during childhood in others. Thus, recently, several authors felt the need to distinguish between these two categories of obese individuals by defining two terms: metabolically ‘healthy’ obese (MHO) and metabolically ‘unhealthy’ obese (MUO) [47]. Moreover, it was proven that MHO might migrate into the MUO phenotype during puberty [48] (Table 1). Aside from the external obesogenic factors, studies suggested that the microbiota itself might be obesogenic, proving that fecal transplantation from an obese donor increases the risk of weight gain in the case of germ-free recipients following the same diet [49]. The colonization of the human intestine begins during birth and continues until the age of 3–5 years when it resembles that of an adult reaching maturity [50]. Thus, in the setting of vaginal delivery, the infant’s microbiome is similar to that of the maternal vaginal microbiota consisting of Escherichia spp. and Lactobacillus, but also other members of the Enterobacteriaceae family [30,31][30][31]. Within the first day after birth, the infant’s gut begins to be colonized with Bifidobacterium and Clostridium [31], which, in fact, is a consequence of breastfeeding, as it is well-documented that breastfed infants have higher levels of Bifidobacteria compared to those fed with a formula, whose microbiota is more diversified, including Staphylococci, Clostridia, Bacteroides, Enterobacteria, Enterococci, and the genus Atopobium [6]. Although important changes in the microbiome composition occur after the introduction of solid feeding, it seems that the microbiota assessed at the age of three months might be considered a more reliable predictor of the future risk of increased weight gain than the profile assessed at the age of 12 months [43]. In fact, the infant’s microbiota at the age of three months is mainly composed of Bifidobacteriaceae, Bacteroidaceae, Enterobacteriaceae, Lachnospiraceae, Veillonellaceae, and Ruminococcaceae [43]. Nevertheless, once complementary feeding is introduced, an important maturation stage occurs within the infant’s microbiota, resulting in an important increase in alpha diversity, consisting of Proteobacteria and Actinobacteria replacement by the phyla Bacteroidetes and Firmicutes, as well as a considerably increased production of short-chain fatty acids (SCFAs) [43] (Table 1). Several mechanisms were hypothesized regarding the role of intestinal microbiota in the development of obesity, such as the ability of intestinal microbes to extract energy from nondigestible polysaccharides resulting in an overproduction of SCFAs and subsequent impaired food absorption, a decrease in the AMP kinase with a decreased oxidization of fatty acids in the muscle, and a higher hepatic lipogenesis mediated via ChREBP/SREBP-1, along with an altered metabolism of bile acids affecting the effective transport of cholesterol [6]. In addition, it was also noticed that hormonal-mediated mechanisms participated in the pathogenesis of obesity via glucagon-like peptide 1 (GLP-1) and GLP-1 receptors G protein-coupled 43 and 41 (GPR41 and GPR43). In fact, GPR41 is involved in the synthesis of the intestinal anorexigenic hormone called peptide YY, responsible for regulating satiety, decreasing gastric emptying, and reducing intestinal transit time, as well as for increasing energy harvest, along with hepatic lipogenesis, while GPR43 has the ability to sense postprandial energy excess and to regulate energy expenditure [51,52,53][51][52][53]. A less studied relationship between the host energy metabolism is mediated by fasting-induced adipocyte factor (FIAF), which, if inhibited by gut microbes, leads to no inhibitory action on lipoprotein lipase (LPL) and the subsequent accumulation of adipocyte within peripheral tissues [54]. In terms of the bacterial composition of the gut microbiota, several studies indicated that, depending on the predominant species, it might have either a positive or negative impact on the development of obesity. Therefore, among the bacteria that were associated with a lower body mass index, studies pointed out Fecalibacterium prausnitzii and Clostridium difficile, Staphylococcus genus, or Bacteroidetes phylum [55]. Nevertheless, in term of Bacteroidetes, the findings reported in the current studies are rather controversial. Thus, a systematic review indicated a possible positive association between obesity and B. fragilis. Contrariwise, in the same review, the authors pointed out that other bacteria belonging to the Bacteoridetes phylum, i.e., Bacteroidetes and Prevotella, were less abundant in obese subjects in spite of their role as triggers of intestinal inflammation [55]. When assessing the entire Bacteroidetes phylum, the authors proved a decrease of their levels in children with obesity in comparison to the control group [55]. Similar to B. fragilis, C. leptum and E. hallii were also proven to be more abundant in obese/overweight infants and preschool/school-aged children [55]. All these findings reported in the previously mentioned studies were all related to the role of these bacteria as promoters or suppressors of inflammation [55] (Table 1). Early disruptions in microbial gut compositions, triggered especially by the administration of antibiotics, were proven to be associated with an increased predisposition to obesity [56]. An animal model study proved that the exposure of mice to low-dose penicillin induces important changes in the gut microbiota, which, if transplanted to germ-free recipients, trigger body fat deposition, weight gain, and the severe impairment of gut immune system functionality, consisting in diminishing the ileal expression of genes responsible for encoding antimicrobial peptides, as well as those which regulate T helper 17 cell populations [57]. Moreover, repeated administrations of antibiotics have the same effect in terms of weight gain, increasing the bone mineral content and growth as well due to microbiome perturbations, which, in fact, are directly related to the classes of antibiotics and the number of pulses [58]. Aside from antibiotics, a study performed on children from the United States also revealed the same effect on weight gain in the setting of proton pump or histamine type 2 receptor antagonists administered within the first two years of life, the association being stronger if multiple classes of antibiotics are prescribed, or if the course of therapy is prolonged [59]. Nutritional interventions including prebiotics, probiotics, and synbiotics were suggested to have a positive effect on improving the gut microbiota, reducing cardio-metabolic risk factors, such as obesity [60,61][60][61]. Thus, the modulation of intestinal microbiome composition using these compounds was recently suggested to have a major positive impact on obesity-associated gut dysbiosis [62]. Prebiotics represent non-digestible substrates meant to favor the development of beneficial bacterial species, which are selectively utilized by the host’s micro-organisms [63]. Probiotics contain live micro-organisms carefully selected in adequate amounts [64], while synbiotics represent a mixture of pre- and probiotics [65]. Thus, based on the aforementioned crosstalk between obesity and gut microbiota, studies tried to assess the impact of different bacteria supplementation on body weight. Akkermansia muciniphila is one of the bacteria that proved to be effective in both animal and human model studies. In a study performed on mice, Akkermansia muciniphila administration reversed insulin resistance and weight gain as a result of a high-fat diet [66], most likely due to its outer membrane protein which interacts with toll-like receptor 2 in order to improve the alterations of the gut barrier, resulting in the normalization of body weight and insulin resistance [67]. Moreover, a randomized, placebo-controlled, double-blind study involving subjects with overweight/obesity and insulin resistance pointed out that a supplementation of 1010 Akkermansia muciniphila per day for three months was associated with a decrease in plasma insulin and cholesterol levels, improving insulin sensitivity [68]. In spite of the lack of significance, the study also underlined that this bacterium might also have a positive effect on fat mass, hip circumference, and body weight. Similar results were also reported for Christensenella minuta in a study performed on mice [69]. The supplementation of Lactobacillus rhamnosus strain GG and Lactobacillus salivarius were also assessed in children or adolescents with overweight and obesity, but, although the first lead to important changes in gut microbial composition, none of them displayed an effect on the body mass index z score, or body fat mass [70,71,72][70][71][72]. Despite their lack of effectiveness when administered separately, a recent study also involving pediatric subjects pointed out that the supplementation of multi-strain probiotics consisting of Lactobacillus salivarius AP-32, Lactobacillus rhamnosus bv-77, and Bifidobacterium animalis CP-9 resulted in a lowered serum total cholesterol, low density lipoprotein, tumor necrosis factor-alpha, and leptin levels, and a reduced body mass index, but, at the same time, induced an elevation of adiponectin and high-density lipoprotein [73]. Moreover, the authors suggested that the positive effects on the lipid pathway might be related to Lactobacillus spp., while those on adiponectin might be related to Bifidobacterium animalis. Verma et al. also sustained that compounds that contain multiple species of probiotics (Lactobacillus plantarum DSM 24730, Lactobacillus plantarum DSM 24731, Lactobacillus plantarum DSM 24735, Lactobacillus plantarum DSM 24801, Lactobacillus paracasei DSM 24737, Lactobacillus salivarius DSM 24800, Lactobacillus delbrueckii DSM 25998, Bifidobacterium animalis DSM 24736, Bifidobacterium breve DSM 24732, and Pediococcus pentosaceus DSM 24734) might be effective in improving the gut microbial composition and fasting glucose levels in adolescents with severe obesity [74]. However, probiotics might not work as preventive therapies for obesity, according to a study in which the authors administered Lactobacillus paracasei F19 to 120 infants from the age of 4 to 13 months, during weaning, and after a close follow-up of this population, and noticed no modulatory effect of this probiotic on either growth or body composition at school age [75]. The findings reported in studies using Lactobacillus spp. as probiotics might differ even between pediatric and adult populations, according to a meta-analysis which observed that adults that were administered this probiotic experienced weight loss, while children presented minor weight gain [76]. Nevertheless, Luoto et al. defined two phases in the development of obesity—an initial one beginning in the perinatal phase to 48 months of age, and a second one after the age of 4 years—and stated that, if Lactobacillus rhamnosus GG is supplemented in future mothers 4 weeks before delivery is expected and in infants until the age of 6 months, the children will have a healthy growth pattern, but the effect lasts only during the first phase of obesity, with no effect at 10 years of age [77] (Table 1). It is well-documented that children born to mothers with overweight and obesity have a higher risk of becoming obese later in life [50,78,79,80][50][78][79][80]. Therefore, this risk might be lowered with proper nutritional intervention even during pregnancy. According to Saros et al., probiotics (Lactobacillus rhamnosus HN001 and Bifidobacterium animalis ssp. lactis 420) administration alone or combined with fish oil during pregnancy and in the first 6 months postpartum decreased the risk of obesity in their offspring at the age of 24 months [81]. Interestingly, recent evidence suggests that probiotics administered in pregnant women might act as a gene-targeted therapy since they might significantly decrease the DNA methylation in certain gene promoters of obesity in both mothers and their children, subsequently decreasing the risk for excessive weight gain [82]. Moreover, breast-milk-derived probiotics such as Lactobacillus plantarum 73a and Bifidobacterium animalis subsp. lactis INL1 were also suggested to act as potential candidates in the management of pediatric obesity according to the study of Oddi et al., who proved that the administration of these two probiotics isolated from breastmilk results in a decrease of phylum Proteobacteria and genera Shigella, Escherichia, and Clostridium sensu stricto 1, which are all known to trigger obesity and its associated inflammatory status or insulin resistance [83]. In terms of prebiotics, a clinical trial designed for improving the microbiota in obese individuals, including 48 healthy overweight and obese adults, assessed the administration of 21 g of oligofructose compared to a placebo every day for 12 weeks, and concluded that oligofructose favors weight loss, reduces calorie intake and the levels of ghrelin, also known as the hunger hormone, improves glucose regulation, and increases the concentrations of satiety hormone YY [51]. Another clinical trial including 42 children with overweight and obesity using oligofructose-enriched inulin administered daily for 16 weeks in a dose of 8 g/day indicated that it might be associated with a reduced weight gain, and improved truncal fat disposition and body weight z-scores, as well as lower levels of interleukin-6 and triglycerides, when compared to a group receiving an isocaloric placebo [52]. In addition, the authors also noticed an increase of fecal Bifidobacterium spp. in the overweight/obese group after the administration of oligofructose-enriched inulin. Other microbiota-targeting therapies for obesity involving prebiotics used fibers as supplements and noticed that individuals receiving high-dietary-fiber supplements for 12 weeks expressed an increased abundance of SCFA-producing bacteria within their gut microbial composition, associated also with improved fasting and postprandial blood glucose levels, as well as hemoglobin A1c [53] (Table 1). Synbiotics represent compounds that have the same properties of prebiotics and probiotics, together promoting probiotic survival in the gastrointestinal tract, and they might result in a better outcome regarding the host’s health when compared to prebiotics and probiotics separately [84,85][84][85]. A recent study performed on 60 children with overweight and obesity aged between 8 and 18 years assessed the combination between Lactobacillus indicus and Lactobacillus coagulans as probiotics, combined with short-chain fructo-oligosaccharide as a prebiotic, and noticed a significant reduction of waist–height ratio in children receiving this combination compared to those receiving the placebo [86]. Nevertheless, another study involving obese children and teenagers pointed out a decrease in weight and body mass index after one month of supplementation with synbiotics [87]. Similarly, a meta-analysis of nine randomized controlled trials proved that synbiotic supplementation in children and adolescents with obesity might be associated with a significant decrease in the body mass index z score [88]. Kilic Yildirim et al. conducted a randomized, double-blind, placebo-controlled trial on 61 children with exogenous obesity and pointed out that the group which received a synbiotic supplement once daily, consisting of a mixture of probiotics, including Lacticaseibacillus rhamnosus, Lactobacillus acidophilus, Bifidobacterium longum, Bifidobacterium bifidum, and Enterococcus faecium, combined with fructo-oligosaccharides, for 12 weeks presented significantly greater changes in weight, body mass index, and anthropometric parameters like waist circumference and waist-circumference-to-height ratio when compared to the placebo group [89]. Thus, the authors concluded that a multispecies synbiotic intake, along with exercise and diet, might represent an effective weight-loss strategy for pediatric obesity. Similar studies performed on adults revealed contradictory effects of synbiotics on both the body mass index, and body fat or anthropometric parameters in obese subjects [90,91][90][91] (Table 1). The impact and mechanism through which probiotics improve the wellbeing of children with obesity are related in Figure 1.Table 1.
The effects of prebiotics, probiotics, and symbiotic on children’s obesity.
| Reference (Author, Year) | Type of Study | Study Population | Objectives/Outcomes Measured | Prebiotics, Probiotics, and Synbiotic | Conclusions/Outcome |
|---|---|---|---|---|---|
| Vajro et al., 2011 [70] | Double-blind, placebo-controlled pilot study | n = 20 (10 probiotic/10 placebo) Age: 9–13 years BMI > 95th percentile |
Effects of short-term probiotic on children with obesity-related liver disease | Lactobacillus rhamnosus GG (12 billion CFU/day) for 8 weeks | Decrease in alanine aminotransferase and in antipeptidoglycan polysaccharide antibodies, irrespective of changes in BMI z score and visceral fat. |
| Gobel et al., 2012 [71] | Double-blind placebo-controlled trial | n = 50 (27 LS-33/23 placebo) Age: 12–15 years BMI > 95th percentile |
Effect of Lactobacillus salivarius Ls-33 on biomarkers related to inflammation and metabolic syndrome (MS) | Lactobacillus salivarius Ls-33 (1010 CFU/day) for 12 weeks | No effects of probiotic strain Ls-33 on either the inflammatory markers or the markers of MS. |
| Marcelo et al., 2022 [72] | Non-randomized controlled, prospective, double-blind interventional clinical trial | n = 44 (22 probiotic/22 placebo) Age: 8–17 years BMI > 95th percentile |
Impact of probiotic supplementation therapy on anthropometric values and body composition | Lactobacillus rhamnosus IAL 1883 for 6 months | Supplementation with Lactobacillus rhamnosus IAL 1883 was not effective for weight loss or improving the body composition. |
| Chen et al., 2022 [73] | Double-blind, randomized, placebo-controlled trial | n = 53 (27 probiotic/26 placebo) Age: 6–18 years BMI ≥ 85th percentile |
Effects of multi-strain probiotics on the gut microbiota and weight control/BMI, LDLC, HDLC, adiponectin, leptin | Multi-strain probiotics consisting of Lactobacillus salivarius AP-32, Lactobacillus rhamnosus bv-77, and Bifidobacterium animalis CP-9 for 12 weeks | Lowered serum TC, LDLC, TNF-α, and leptin, and reduced BMI. Elevation of adiponectin and HDLC. |
| Verma et al., 2021 [74] | Randomized, double-blind, placebo-controlled pilot study |
3.2. Metabolic Syndrome and Insulin Resistance
Recently, NAFLD was described as a manifestation of metabolic syndrome [101,102][101][102]. The prevalence of metabolic syndrome in the pediatric population varies between 1.2% and 22.6%, increasing to up to 60% in children with overweight and obesity [103]. Moreover, the prevalence was reported to be higher in boys and increases with age [104]. Recent evidence from the review of Fiore G. et al. sustains the beneficial role of pre-, pro-, and synbiotics in children with obesity and metabolic disorders [105]. The role of probiotics in the management of childhood metabolic syndrome was assessed in studies performed on both mothers and their infants, suggesting that the benefits of these supplements consist of regulating maternal obesity and increased blood glucose levels, as well as modulating the infant’s gut microbial composition through the placenta or breastmilk [106,107,108,109][106][107][108][109]. Unfortunately, two randomized clinical trials that assessed the impact of Lactobacillus rhamnosus GG and Lactobacillus paracasei ssp. F19 on at least one of the components of metabolic syndrome in children aged between 2 and 19 years failed to identify significant differences between the probiotic and the placebo group [75,77][75][77]. Recent evidence highlighted the role of Blautia species in modulating glucose metabolism in children [110]. Although certain species were associated with a negative effect due to their proinflammatory responses such as Blautia cocoides, others were, on the contrary, proven to exert anti-inflammatory effects including Blautia wexlerae and Blautia luti [110,111][110][111]. Taking into account the reduction of the latter two species in obese children, studies proved that dietary interventions including arabinoxylans added to wheat bran extract resulted in an increase of these bacterial species in the bacterial community of these pediatric patients [112,113][112][113]. Another larger interventional study included 101 obese participants with insulin resistance aged between 6–18 years and concluded that Bifidobacterium breve B632 and Bifidobacterium breve BR03 administered for 8 weeks leads to a significant improvement in insulin sensitivity, being also associated with lower levels of fasting insulin and alanine aminotransferase, but also decreased body mass index and waist circumference [114]. Lactobacillus rhamnosus bv-77, Lactobacillus salivarius, and Bifidobacterium animalis were assessed on a pediatric population from China and they were proven not only to significantly reduce body mass index in obese children, but also to cause a significant improvement in blood lipid content [73]. Bifidobacterium pseudocatenulatum CECT 7765 was recently assessed in a pediatric population with obesity and insulin resistance, and the authors concluded that this probiotic was associated with not only lower levels of C-reactive protein and monocyte chemoattractant protein-1, but also elevated levels of high-density lipoprotein cholesterol and omentin-1, highlighting the major anti-inflammatory effect of this bacteria in the studied population [115]. Contrariwise, a study which assessed the role of synbiotics (Bifidobacterium bifidum, Bifidobacterium longum, Lactobacillus acidophilus, Lacticaseibacillus rhamnosus, Enterococcus faecium, and fructo-oligosaccharides) failed to prove any effect on lipid parameters and glucose metabolism [89] (Table 2).Table 2.
Implications of prebiotics, probiotics, and symbiotic in obesity-related complications.
| Reference (Author, Year) | Type of Study | Study Population | Objectives | Prebiotics, Probiotics and Synbiotic | Obesity | Conclusions | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NAFLD/ NASH |
Metabolic Syndrome | Insulin Resistance | ||||||||||
| Loomba et al., 2017 [97] | Prospective study | n = 86 NAFLD Age: >18 years |
Identification of fecal-microbiome-derived metagenomic signature to detect advanced fibrosis in NAFLD | − | + | − | − | Escherichia, Enterobacteria, Proteobacteria, and Bacteroides are more commonly found in patients with NASH. | ||||
| Vajro et al., 2011 [70] | Double-blind, placebo-controlled pilot study | n = 20 (10 probiotic/10 placebo) Age: 9–13 years BMI > 95th percentile |
Effects of short-term probiotic on children with obesity-related liver disease | Lactobacillus rhamnosus GG (12 billion CFU/day) for 8 weeks | + | − | − | Lactobacillus rhamnosus decrease of ALT in children with hepatic steatosis. | ||||
| Alisi et al., 2014 [98] | Parallel-arm double-blind trial | n = 44 (22 VSL#3/22 placebo) Age: 9–13 years BMI > 85th percentile Obese + NAFLD |
Effect of VSL#3 on structural improvement of FL in obese children with biopsy-proven NAFLD | VSL#3 is a mixture of Streptococcus thermophilus, Bifidobacteria breve, Bifidobacteria infantis, Bifidobacteria longum, Lactobacillus acidophilus, L. plantarum, L. paracasei, and L. delbrueckii subsp. bulgaricus 1 sachet/day of VSL#3 < 10 years, 2 sachets/day of VSL#3 > 10 years vs. placebo for 4 months | + | − | − | Streptococcus thermophilus, lactobacilli, and bifidobacteria might improve the ultrasound of the liver after 4 months of administration. | ||||
| Famouri et al., 2017 [99] | Randomized, triple-blind, placebo-controlled trial | n = 64 (32 probiotic/32 placebo) Age: 10–18 years BMI > 85th percentile Obese + NAFLD |
Effects of probiotics on NAFLD in obese children and adolescents | Lactobacillus acidophilus ATCC B3208, 3 × 109 CFU; Bifidobacterium lactis DSMZ 32269, 6 × 109 CFU; Bifidobacterium bifidum ATCC SD6576, 2 × 109 CFU; Lactobacillus rhamnosus DSMZ 21690, 2 × 109 CFU 1 caps/day vs. placebo for 12 weeks | + | − | − | Lactobacillus rhamnosus, Lactobacillus acidophilus, Bifidobacterium lactis, and Bifidobacterium bifidum for 12 weeks in obese children → improving AST, ALT level, liver steatosis, and lipid profile. | ||||
| Rodrigo et al., 2022 [100] | n | = 15 (8 probiotic/7 placebo) Age: ≥13 years BMI ≥ 99th percentile |
Effect of probiotics on gut microbiota and insulin resistance | Double-blind, randomized, placebo-controlled trial | n = 84 (43 probiotic/41 placebo) Age: 5–15 years BMI > +2 standard deviation for age AST/ALT < 1 Hepatic steatosis, grade I to IIIVisbiome® containing probiotics (Lactobacillus plantarum DSM 24730, Lactobacillus plantarum DSM 24731, Lactobacillus plantarum DSM 24735, Lactobacillus plantarum DSM 24801, Lactobacillus paracasei DSM 24737, Lactobacillus salivarius DSM 24800, Lactobacillus delbrueckii DSM 25998, Bifidobacterium animalis DSM 24736, Bifidobacterium breve DSM 24732, and Pediococcus pentosaceus DSM 24734) two sachets/day 12 weeks |
Improve fasting glucose levels and gut microbial composition (decrease F/B ratio). | ||||||
| Effects of probiotics on metabolic derangement in obese children with nonalcoholic fatty liver disease/nonalcoholic steatohepatitis (NAFLD/NASH). | BioKult 14 strain probiotic capsule: 1 caps/day < 12 years, 2 caps/day ≥ 12 years 6 months | + | − | − | Significant decrease of body mass index irrespective of the group which received probiotic with 14 bacterial strains or the placebo group. | Karlsson et al., 2015 [75] | Double-blind, randomized, placebo-controlled intervention trial | n = 120 (58 probiotic/62 placebo) Age: 8–9 years |
Long-term effect of feeding with Lactobacillus paracasei F19 (LF19) on body composition, growth, and metabolic markers (TC, HDLC, apo A-1, apo B, TG, glucose, AST, ALT, S-insuline) | Administration of | ||
| Kilic Yildirim et al., 2022 [89] | Lactobacillus paracasei F19 | from 4 to 13 months of age, 1 × 10 | 8 | CFU/at least once daily | LF19 had no modulatory effect on growth (BMI z-score, sagittal abdominal diameter, fat-free mass, fat mass index, truncal fat %, android or gynoid fat %) or body composition at school age and no long-term impact on metabolic markers. | |||||||
| Randomized, double-blind, placebo-controlled trial | n | = 61 (30 synbiotic/31 placebo) Age: 8–17 years BMI > 95th percentile |
Effects of synbiotic on anthropometric measurements, glucose metabolism, and lipid parameters | Synbiotic including Lacticaseibacillus rhamnosus, Lactobacillus acidophilus, Bifidobacterium longum, Bifidobacterium bifidum, and Enterococcus faecium, combined with fructo-oligosaccharides for 12 weeks vs. placebo. | No impact on NAFLD | − | − | Synbiotics (Bifidobacterium longum, Bifidobacterium bifidum, Enterococcus faecium, Lactobacillus acidophilus, Lacticaseibacillus rhamnosus, and fructo-oligosaccharides) → no impact on NAFLD and no effect on lipid parameters and glucose metabolism. | Luoto et al., 2010 [77] | |||
| Karlsson et al., 2015 [75, | Randomized, double-blind, prospective follow-up study | 77][75][77]n = 113 (77 probiotics/82 placebo) |
Double-blind, randomized, placebo-controlled intervention trialAge: 2 weeks–10 years | nImpact of perinatal probiotic intervention on childhood growth patterns and the development of overweight during a 10-year follow-up | = 120 (58 probiotic/62 placebo) Age: 8–9 yearsAdministration of Lactobacillus rhamnosus |
Long-term effect of feeding with Lactobacillus paracasei F19 (LF19) on body composition, growth, and metabolic markers (TC, HDLC, apo A-1, apo B, TG, glucose, AST, ALT, S-insuline) (1 × 1010 CFU) in mothers 4 weeks before delivery and in infants until the age of 6 months | Early probiotics may restrain excessive weight gain during the first years of life. | |||||
| Administration of | Lactobacillus paracasei F19 | from 4 to 13 months of age, 1 × 10 | 8 | CFU/at least once daily | − | No effect | − | Lactobacillus rhamnosus GG and Lactobacillus paracasei ssp. F19 → no effect on metabolic syndrome. | Saros et al., 2023 [81] | Double-blind, placebo-controlled randomized trial | n = 439 pregnant women BMI ≥ 25 kg/m2 pre-pregnancy n = 330 children Age ≤ 2 years 4 parallel groups: fish oil + placebo, probiotics + placebo, probiotics + fish oil, placebo + placebo |
Fish oil and/or probiotic effect in pregnant women with overweight or obesity on the tendency of their 24-month-old children to become overweight |
| Benítez-Páez et al., 2020 [110] | Cross-sectional study | n= 51 (16 lean/20 obese/15 obese + IR Age: 5–17 years BMI z-score of ≥2 SD |
Study of gut microbiota profile of lean and obese children with/without insulin resistance and associations with specific obesity-related complications and metabolic inflammation | Administration of Lactobacillus rhamnosus HN001 and Bifidobacterium animalis ssp. lactis 420 (1010 CFU/day) alone or combined with fish oil during pregnancy and in the first 6 months postpartum | Decreased the risk of obesity in their children at the age of 24 months. | |||||||
| − | − | + | − | Blautia species | → modulating glucose metabolism in children. | Depletion of | B. luti and B. wexlerae species occur in obesity and lead to metabolic inflammation and insulin resistance. | Nicolucci et al., 2017 [52] | Randomized, double-blind, placebo-controlled trial | n | ||
| Solito et al., 2023 [114] | Cross-over, double-blind, randomized control trial | = 42 (22 OI/20 placebo) Age: 7–12 years BMI ≥ 85th percentile |
Effects of prebiotics on body composition, markers of inflammation, bile acids in fecal samples, and composition of the intestinal microbiota | n= 101 (51 probiotic/50 placebo) obese+ IRAdministration of oligofructose-enriched inulin (OI) 8 g/day for 16 weeks | Age: 6–18 years |
Impact of a probiotic supplementation in pediatric obesity on weight, metabolic alterations, and selected gut microbial groups | 2 × 109 CFU/AFU/day of Bifidobacterium breve BR03 (DSM 16604) and B. breve B632 (DSM 24706) or placebo for 8 weeksOI administration reduces body weight z-score, percent body fat, percent trunk fat, serum interleukin 6, and triglycerides; increase fecal Bifidobacterium spp. | |||||
| − | Positive effect | Improve insulin sensitivity | Bifidobacterium breve B632 | and | Bifidobacterium breve BR03 | administered for 8 weeks → improvement in insulin sensitivity and decreased body mass index and waist circumference. | Atazadegan et al., 2023 [86] | Randomized double-blind, placebo-controlled trial | n = 60 (30 synbiotic/30 control) | |||
| Chen et al., 2022 [73] | Double-blind, randomized, placebo-controlled trial | Age: 8–18 years BMI ≥ 85th percentile |
Effect of synbiotic on anthropometric indices and body composition | Combination of | n = 53 (27 probiotic/26 placebo)Lactobacillus indicus 6 × 109 colony forming units (CFU) and Lactobacillus coagulans 6 × 109 CFU twice a day as probiotics combined with short-chain fructo-oligosaccharide for 8 weeks | A significant reduction of waist–height ratio. | ||||||
| Effects of multi-strain probiotics on the gut microbiota and weight control/BMI, LDLC, HDLC, adiponectin, and leptin | Multi-strain probiotics consisting of | Lactobacillus salivarius AP-32 | , | Lactobacillus rhamnosus bv-77, and Bifidobacterium animalis CP-9 for 12 weeks | − | Positive effect | − | Lactobacillus salivarius and Bifidobacterium animalis on a pediatric population from China → significantly reduce body mass index in obese children and a significant improvement of blood lipid content | Ipar et al., 2015 [87] | Open-label, randomised, controlled study | n = 86 (43 synbiotic/43 control) Age: 5–17 years BMI > 95th percentile |
Effects of a synbiotic on anthropometric measurements, lipid profile, and oxidative stress parameters |
| Sanchis-Chordà et al., 2019 [115] | A probiotic mixture including | Lactobacillus acidophilus | , | Lactobacillus rhamnosus, Bifidobacterium bifidum, Bifidobacterium longum, and | Prospective analytical intervention study | n = 56 (28 probiotic/28 control Age: 7–16 years obese + IR BMI z-score of ≥2 SDEnterococcus faecium, combined with fructo-oligosaccharydes for one month |
Effect of Decrease of weight and BMI. Decrease of serum total cholesterol, low density lipoprotein cholesterol and total oxidative stress levels. |
|||||
| Kilic Yildirim et al., 2022 [89] | Randomized, double-blind, placebo-controlled trial | n = 61 (30 synbiotic/31 placebo) Age: 8–17 years BMI > 95th percentile |
Effects of synbiotic on anthropometric measurements, glucose metabolism, and lipid parameters | Synbiotic including Lacticaseibacillus rhamnosus, Lactobacillus acidophilus, Bifidobacterium longum, Bifidobacterium bifidum, and Enterococcus faecium, combined with fructo-oligosaccharides for 12 weeks vs. placebo. | Decreased weight, BMI, and anthropometric parameters. | |||||||
Legend: ALT: alanine transaminase; apo A-1: apolipoprotein A-1; apo B: apolipoprotein B; AST: aspartate transaminase; BMI: body mass index; CFU: colony forming units; F/B ratio: firmicutes-to-bacteroidetes; HDLC: high-density lipoprotein cholesterol; LDLC: low-density lipoprotein cholesterol; SHIME®: Simulator of the Human Intestinal Microbial Ecosystem; TC: total cholesterol; TG: triglycerides; TNF-α: tumor necrosis factor-alpha.
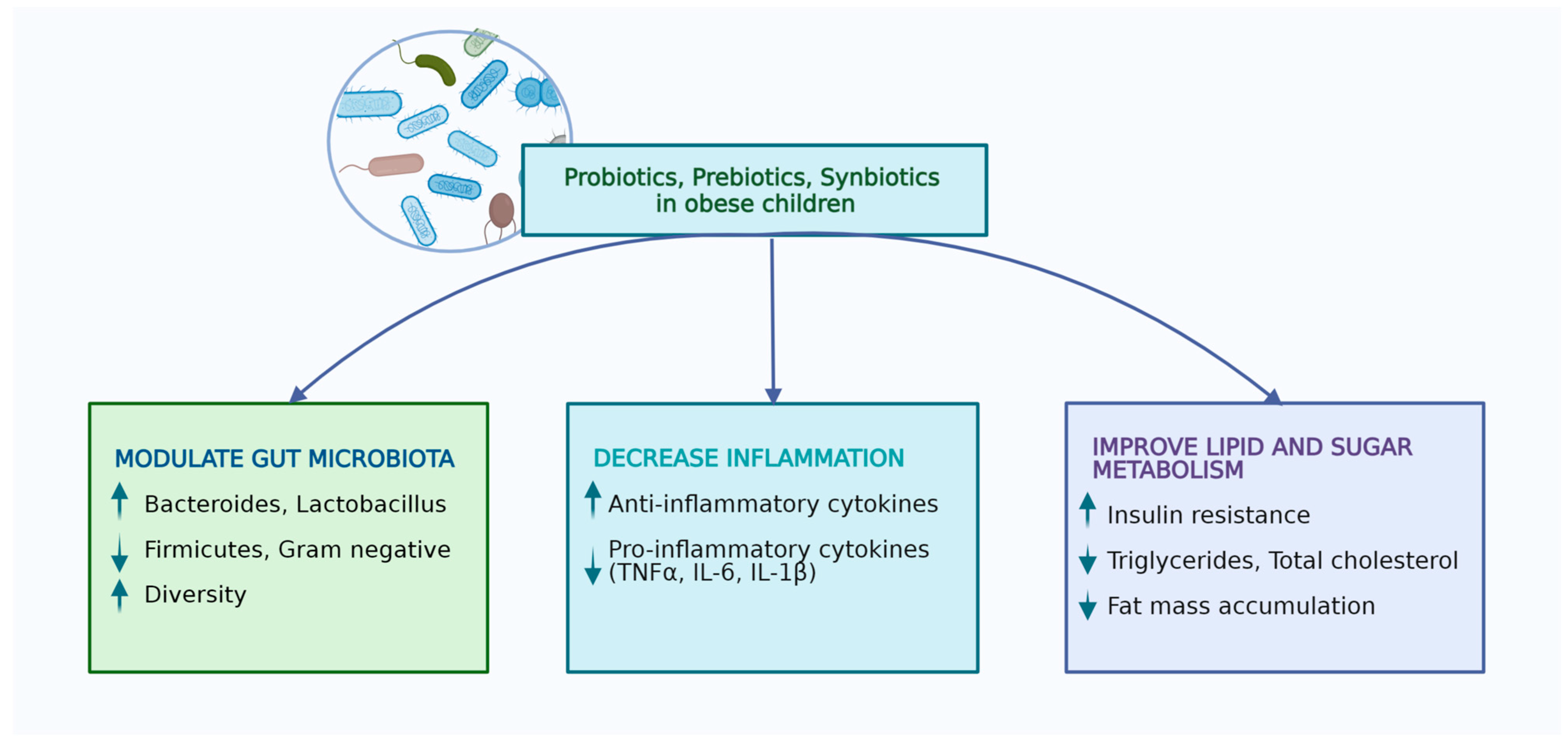
Figure 1.
Effects of probiotics, prebiotics, and synbiotics in obese children (performed with Biorender licence).
3. Implications in Obesity-Related Complications
3.1. Non-Alcoholic Fatty Liver Disease (NAFLD) and Non-Alcoholic Steatohepatitis (NASH)
The average prevalence of NAFLD in the general pediatric population is 7.6%, but, when assessing only the subgroup of obese children, it reaches up to 34.2%, affecting more commonly the male gender [92]. NAFLD comprises a wide spectrum of hepatic disorders, varying from isolated steatosis and liver fibrosis to cirrhosis, resulting eventually in the development of hepatocellular carcinoma. This condition is recognized when more than 5% of hepatocytes develop steatosis, noticed either radiologically or histologically without the presence of other causes that might trigger it, such as hereditary liver disorders, viral hepatitis, or increased alcohol consumption [93]. NASH is a more severe form of liver impairment consisting of hepatocyte degeneration, presenting a balloon shape associated with diffuse inflammatory infiltrate within the hepatic lobules plus/minus fibrosis [94]. Taking into account the fact that the liver is primarily exposed to the toxic products of the gut such as damaged metabolites, bacteria, and their products [95], it is not surprising that several studies tried to find the relationship between the gut microbial composition and the occurrence of NAFLD. Thus, certain authors noticed that the gut microbiota signature is different for each stage of NAFLD [96], as proven by the fact that species like Escherichia, Enterobacteria, Proteobacteria, and Bacteroides are more commonly found in patients with NASH when compared to matched controls [97]| Bifidobacterium pseudocatenulatum CECT 7765 | |||
| on inflammatory cytokines, cardiometabolic risk factors, and gut microbiota composition in obese children with IR | |||
| B. pseudocatenulatum (CECT 7765) | |||
| 1 × 10 | |||
| 9 | |||
| –1 × 10 | |||
| 10 | |||
| CFU per day for 91 days vs. control | |||
| − | |||
| + | + | Bifidobacterium pseudocatenulatum CECT 7765 | → determine lower levels of C-reactive protein and monocyte chemoattractant protein-1 and elevated levels of high-density lipoprotein cholesterol and omentin-1 → anti-inflammatory effect |
Legend: BMI: body mass index; CFU: colony forming units; FL: fatty liver; HDLC: high-density lipoprotein cholesterol; IR: insulin resistance; LDLC: low-density lipoprotein cholesterol.
References
- Strauss, R.S.; Pollack, H.A. Epidemic Increase in Childhood Overweight, 1986–1998. JAMA 2001, 286, 2845–2848.
- GBD 2015 Obesity Collaborators; Afshin, A.; Forouzanfar, M.H.; Reitsma, M.B.; Sur, P.; Estep, K.; Lee, A.; Marczak, L.; Mokdad, A.H.; Moradi-Lakeh, M.; et al. Health Effects of Overweight and Obesity in 195 Countries over 25 Years. N. Engl. J. Med. 2017, 377, 13–27.
- Han, J.C.; Lawlor, D.A.; Kimm, S.Y.S. Childhood Obesity. Lancet 2010, 375, 1737–1748.
- Zhao, X.; Gang, X.; Liu, Y.; Sun, C.; Han, Q.; Wang, G. Using Metabolomic Profiles as Biomarkers for Insulin Resistance in Childhood Obesity: A Systematic Review. J. Diabetes Res. 2016, 2016, 8160545.
- Concepcion, J.; Chen, K.; Saito, R.; Gangoiti, J.; Mendez, E.; Nikita, M.E.; Barshop, B.A.; Natarajan, L.; Sharma, K.; Kim, J.J. Identification of Pathognomonic Purine Synthesis Biomarkers by Metabolomic Profiling of Adolescents with Obesity and Type 2 Diabetes. PLoS ONE 2020, 15, e0234970.
- Bosco, A.; Loi, M.; Pinna, G.; Pintus, R.; Fanos, V.; Dessì, A. Childhood Obesity and the Cryptic Language of the Microbiota: Metabolomics’ Upgrading. Metabolites 2023, 13, 414.
- Mastrangelo, A.; Martos-Moreno, G.Á.; García, A.; Barrios, V.; Rupérez, F.J.; Chowen, J.A.; Barbas, C.; Argente, J. Insulin Resistance in Prepubertal Obese Children Correlates with Sex-Dependent Early Onset Metabolomic Alterations. Int. J. Obes. 2016, 40, 1494–1502.
- Ten, S.; Maclaren, N. Insulin Resistance Syndrome in Children. J. Clin. Endocrinol. Metab. 2004, 89, 2526–2539.
- Butte, N.F.; Liu, Y.; Zakeri, I.F.; Mohney, R.P.; Mehta, N.; Voruganti, V.S.; Göring, H.; Cole, S.A.; Comuzzie, A.G. Global Metabolomic Profiling Targeting Childhood Obesity in the Hispanic Population. Am. J. Clin. Nutr. 2015, 102, 256–267.
- Barlow, S.E.; Dietz, W.H. Obesity Evaluation and Treatment: Expert Committee Recommendations. The Maternal and Child Health Bureau, Health Resources and Services Administration and the Department of Health and Human Services. Pediatrics 1998, 102, E29.
- Cardel, M.I.; Atkinson, M.A.; Taveras, E.M.; Holm, J.-C.; Kelly, A.S. Obesity Treatment Among Adolescents: A Review of Current Evidence and Future Directions. JAMA Pediatr. 2020, 174, 609–617.
- Berger, N.A. Young Adult Cancer: Influence of the Obesity Pandemic. Obesity 2018, 26, 641–650.
- Wahl, S.; Yu, Z.; Kleber, M.; Singmann, P.; Holzapfel, C.; He, Y.; Mittelstrass, K.; Polonikov, A.; Prehn, C.; Römisch-Margl, W.; et al. Childhood Obesity Is Associated with Changes in the Serum Metabolite Profile. Obes. Facts 2012, 5, 660–670.
- Borka Balas, R.; Meliț, L.E.; Sarkozi, D.; Ghiga, D.V.; Mărginean, C.O. Cyberbullying in Teenagers—A True Burden in the Era of Online Socialization. Medicine 2023, 102, e34051.
- WHO|Overweight and Obesity. Available online: http://www.who.int/gho/ncd/risk_factors/overweight_adolescents_text/en/ (accessed on 27 March 2021).
- Mǎrginean, C.O.; Mǎrginean, C.; Meliţ, L.E. New Insights Regarding Genetic Aspects of Childhood Obesity: A Minireview. Front. Pediatr. 2018, 6, 271.
- Rugină, C.; Mărginean, C.O.; Meliţ, L.E.; Giga, D.V.; Modi, V.; Mărginean, C. Relationships between Excessive Gestational Weight Gain and Energy and Macronutrient Intake in Pregnant Women. J. Int. Med. Res. 2020, 48, 300060520933808.
- Mărginean, C.; Mărginean, C.O.; Bănescu, C.; Meliţ, L.; Tripon, F.; Iancu, M. Impact of Demographic, Genetic, and Bioimpedance Factors on Gestational Weight Gain and Birth Weight in a Romanian Population: A Cross-Sectional Study in Mothers and Their Newborns: The Monebo Study (STROBE-Compliant Article). Medicine 2016, 95, e4098.
- Aggarwal, B.; Jain, V. Obesity in Children: Definition, Etiology and Approach. Indian J. Pediatr. 2018, 85, 463–471.
- Sender, R.; Fuchs, S.; Milo, R. Revised Estimates for the Number of Human and Bacteria Cells in the Body. PLoS Biol. 2016, 14, e1002533.
- Harris, K.; Kassis, A.; Major, G.; Chou, C.J. Is the Gut Microbiota a New Factor Contributing to Obesity and Its Metabolic Disorders? J. Obes. 2012, 2012, 879151.
- Blaser, M.J.; Devkota, S.; McCoy, K.D.; Relman, D.A.; Yassour, M.; Young, V.B. Lessons Learned from the Prenatal Microbiome Controversy. Microbiome 2021, 9, 8.
- Walter, J.; Hornef, M.W. A Philosophical Perspective on the Prenatal in Utero Microbiome Debate. Microbiome 2021, 9, 5.
- Aagaard, K.; Ma, J.; Antony, K.M.; Ganu, R.; Petrosino, J.; Versalovic, J. The Placenta Harbors a Unique Microbiome. Sci. Transl. Med. 2014, 6, 237ra65.
- Bassols, J.; Serino, M.; Carreras-Badosa, G.; Burcelin, R.; Blasco-Baque, V.; Lopez-Bermejo, A.; Fernandez-Real, J.-M. Gestational Diabetes Is Associated with Changes in Placental Microbiota and Microbiome. Pediatr. Res. 2016, 80, 777–784.
- Zheng, J.; Xiao, X.; Zhang, Q.; Mao, L.; Yu, M.; Xu, J.; Wang, T. The Placental Microbiota Is Altered among Subjects with Gestational Diabetes Mellitus: A Pilot Study. Front. Physiol. 2017, 8, 675.
- Hu, J.; Nomura, Y.; Bashir, A.; Fernandez-Hernandez, H.; Itzkowitz, S.; Pei, Z.; Stone, J.; Loudon, H.; Peter, I. Diversified Microbiota of Meconium Is Affected by Maternal Diabetes Status. PLoS ONE 2013, 8, e78257.
- Rodríguez, J.M.; Murphy, K.; Stanton, C.; Ross, R.P.; Kober, O.I.; Juge, N.; Avershina, E.; Rudi, K.; Narbad, A.; Jenmalm, M.C.; et al. The Composition of the Gut Microbiota throughout Life, with an Emphasis on Early Life. Microb. Ecol. Health Dis. 2015, 26, 26050.
- Meliț, L.E.; Mărginean, C.O.; Săsăran, M.O. The Yin-Yang Concept of Pediatric Obesity and Gut Microbiota. Biomedicines 2022, 10, 645.
- Vangay, P.; Ward, T.; Gerber, J.S.; Knights, D. Antibiotics, Pediatric Dysbiosis, and Disease. Cell Host Microbe 2015, 17, 553–564.
- Beller, L.; Deboutte, W.; Falony, G.; Vieira-Silva, S.; Tito, R.Y.; Valles-Colomer, M.; Rymenans, L.; Jansen, D.; Van Espen, L.; Papadaki, M.I.; et al. Successional Stages in Infant Gut Microbiota Maturation. mBio 2021, 12, e0185721.
- Dominguez-Bello, M.G.; Blaser, M.J.; Ley, R.E.; Knight, R. Development of the Human Gastrointestinal Microbiota and Insights from High-Throughput Sequencing. Gastroenterology 2011, 140, 1713–1719.
- Kumbhare, S.V.; Patangia, D.V.V.; Patil, R.H.; Shouche, Y.S.; Patil, N.P. Factors Influencing the Gut Microbiome in Children: From Infancy to Childhood. J. Biosci. 2019, 44, 49.
- Azad, M.B.; Konya, T.; Maughan, H.; Guttman, D.S.; Field, C.J.; Chari, R.S.; Sears, M.R.; Becker, A.B.; Scott, J.A.; Kozyrskyj, A.L.; et al. Gut Microbiota of Healthy Canadian Infants: Profiles by Mode of Delivery and Infant Diet at 4 Months. CMAJ 2013, 185, 385–394.
- Rutayisire, E.; Huang, K.; Liu, Y.; Tao, F. The Mode of Delivery Affects the Diversity and Colonization Pattern of the Gut Microbiota during the First Year of Infants’ Life: A Systematic Review. BMC Gastroenterol. 2016, 16, 86.
- Song, S.J.; Dominguez-Bello, M.G.; Knight, R. How Delivery Mode and Feeding Can Shape the Bacterial Community in the Infant Gut. CMAJ 2013, 185, 373–374.
- Jakobsson, H.E.; Abrahamsson, T.R.; Jenmalm, M.C.; Harris, K.; Quince, C.; Jernberg, C.; Björkstén, B.; Engstrand, L.; Andersson, A.F. Decreased Gut Microbiota Diversity, Delayed Bacteroidetes Colonisation and Reduced Th1 Responses in Infants Delivered by Caesarean Section. Gut 2014, 63, 559–566.
- Kuhle, S.; Woolcott, C.G. Caesarean Section Is Associated with Offspring Obesity in Childhood and Young Adulthood. BMJ Evid.-Based Med. 2017, 22, 111.
- Rutayisire, E.; Wu, X.; Huang, K.; Tao, S.; Chen, Y.; Tao, F. Cesarean Section May Increase the Risk of Both Overweight and Obesity in Preschool Children. BMC Pregnancy Childbirth 2016, 16, 338.
- Thurl, S.; Munzert, M.; Henker, J.; Boehm, G.; Müller-Werner, B.; Jelinek, J.; Stahl, B. Variation of Human Milk Oligosaccharides in Relation to Milk Groups and Lactational Periods. Br. J. Nutr. 2010, 104, 1261–1271.
- Cabrera-Rubio, R.; Collado, M.C.; Laitinen, K.; Salminen, S.; Isolauri, E.; Mira, A. The Human Milk Microbiome Changes over Lactation and Is Shaped by Maternal Weight and Mode of Delivery. Am. J. Clin. Nutr. 2012, 96, 544–551.
- Collado, M.C.; Laitinen, K.; Salminen, S.; Isolauri, E. Maternal Weight and Excessive Weight Gain during Pregnancy Modify the Immunomodulatory Potential of Breast Milk. Pediatr. Res. 2012, 72, 77–85.
- Gupta, A.; Osadchiy, V.; Mayer, E.A. Brain-Gut-Microbiome Interactions in Obesity and Food Addiction. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 655–672.
- Bergström, A.; Skov, T.H.; Bahl, M.I.; Roager, H.M.; Christensen, L.B.; Ejlerskov, K.T.; Mølgaard, C.; Michaelsen, K.F.; Licht, T.R. Establishment of Intestinal Microbiota during Early Life: A Longitudinal, Explorative Study of a Large Cohort of Danish Infants. Appl. Environ. Microbiol. 2014, 80, 2889–2900.
- Nakayama, J.; Yamamoto, A.; Palermo-Conde, L.A.; Higashi, K.; Sonomoto, K.; Tan, J.; Lee, Y.-K. Impact of Westernized Diet on Gut Microbiota in Children on Leyte Island. Front. Microbiol. 2017, 8, 197.
- Mărginean, C.; Meliț, L.; Ghiga, D.; Mărginean, M. Early Inflammatory Status Related to Pediatric Obesity (STROBE Compliant Article). Front. Pediatr. 2019, 7, 241.
- Blüher, S.; Schwarz, P. Metabolically Healthy Obesity from Childhood to Adulthood—Does Weight Status Alone Matter? Metabolism 2014, 63, 1084–1092.
- Wan Mohd Zin, R.M.; Jalaludin, M.Y.; Yahya, A.; Nur Zati Iwani, A.K.; Md Zain, F.; Hong, J.Y.H.; Mokhtar, A.H.; Wan Mohamud, W.N. Prevalence and Clinical Characteristics of Metabolically Healthy Obese versus Metabolically Unhealthy Obese School Children. Front. Endocrinol. 2022, 13, 971202.
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An Obesity-Associated Gut Microbiome with Increased Capacity for Energy Harvest. Nature 2006, 444, 1027–1031.
- Meliț, L.E.; Mărginean, C.O.; Mărginean, C.D.; Săsăran, M.O. The Peculiar Trialogue between Pediatric Obesity, Systemic Inflammatory Status, and Immunity. Biology 2021, 10, 512.
- Parnell, J.A.; Reimer, R.A. Weight Loss during Oligofructose Supplementation Is Associated with Decreased Ghrelin and Increased Peptide YY in Overweight and Obese Adults. Am. J. Clin. Nutr. 2009, 89, 1751–1759.
- Nicolucci, A.C.; Hume, M.P.; Martínez, I.; Mayengbam, S.; Walter, J.; Reimer, R.A. Prebiotics Reduce Body Fat and Alter Intestinal Microbiota in Children Who Are Overweight or With Obesity. Gastroenterology 2017, 153, 711–722.
- Zhao, L.; Zhang, F.; Ding, X.; Wu, G.; Lam, Y.Y.; Wang, X.; Fu, H.; Xue, X.; Lu, C.; Ma, J.; et al. Gut Bacteria Selectively Promoted by Dietary Fibers Alleviate Type 2 Diabetes. Science 2018, 359, 1151–1156.
- Pérez-Losada, M.; Castro-Nallar, E.; Bendall, M.L.; Freishtat, R.J.; Crandall, K.A. Dual Transcriptomic Profiling of Host and Microbiota during Health and Disease in Pediatric Asthma. PLoS ONE 2015, 10, e0131819.
- Indiani, C.M.D.S.P.; Rizzardi, K.F.; Castelo, P.M.; Ferraz, L.F.C.; Darrieux, M.; Parisotto, T.M. Childhood Obesity and Firmicutes/Bacteroidetes Ratio in the Gut Microbiota: A Systematic Review. Child. Obes. 2018, 14, 501–509.
- Edwards, P.T.; Kashyap, P.C.; Preidis, G.A. Microbiota on Biotics: Probiotics, Prebiotics, and Synbiotics to Optimize Growth and Metabolism. Am. J. Physiol.-Gastrointest. Liver Physiol. 2020, 319, G382–G390.
- Isanaka, S.; Langendorf, C.; Berthé, F.; Gnegne, S.; Li, N.; Ousmane, N.; Harouna, S.; Hassane, H.; Schaefer, M.; Adehossi, E.; et al. Routine Amoxicillin for Uncomplicated Severe Acute Malnutrition in Children. N. Engl. J. Med. 2016, 374, 444–453.
- Nobel, Y.R.; Cox, L.M.; Kirigin, F.F.; Bokulich, N.A.; Yamanishi, S.; Teitler, I.; Chung, J.; Sohn, J.; Barber, C.M.; Goldfarb, D.S.; et al. Metabolic and Metagenomic Outcomes from Early-Life Pulsed Antibiotic Treatment. Nat. Commun. 2015, 6, 7486.
- Stark, C.M.; Susi, A.; Emerick, J.; Nylund, C.M. Antibiotic and Acid-Suppression Medications during Early Childhood Are Associated with Obesity. Gut 2019, 68, 62–69.
- Barengolts, E. Gut Microbiota, Prebiotics, Probiotics, and Synbiotics in Management of Obesity and Prediabets: Review of Randomized Controlled Trials. Endocr. Pract. 2016, 22, 1224–1234.
- Park, D.-Y.; Ahn, Y.-T.; Huh, C.-S.; McGregor, R.A.; Choi, M.-S. Dual Probiotic Strains Suppress High Fructose-Induced Metabolic Syndrome. World J. Gastroenterol. 2013, 19, 274–283.
- Sankararaman, S.; Noriega, K.; Velayuthan, S.; Sferra, T.; Martindale, R. Gut Microbiome and Its Impact on Obesity and Obesity-Related Disorders. Curr. Gastroenterol. Rep. 2023, 25, 31–44.
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert Consensus Document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) Consensus Statement on the Definition and Scope of Prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502.
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert Consensus Document. The International Scientific Association for Probiotics and Prebiotics Consensus Statement on the Scope and Appropriate Use of the Term Probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514.
- Swanson, K.S.; Gibson, G.R.; Hutkins, R.; Reimer, R.A.; Reid, G.; Verbeke, K.; Scott, K.P.; Holscher, H.D.; Azad, M.B.; Delzenne, N.M.; et al. The International Scientific Association for Probiotics and Prebiotics (ISAPP) Consensus Statement on the Definition and Scope of Synbiotics. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 687–701.
- Everard, A.; Belzer, C.; Geurts, L.; Ouwerkerk, J.P.; Druart, C.; Bindels, L.B.; Guiot, Y.; Derrien, M.; Muccioli, G.G.; Delzenne, N.M.; et al. Cross-Talk between Akkermansia Muciniphila and Intestinal Epithelium Controls Diet-Induced Obesity. Proc. Natl. Acad. Sci. USA 2013, 110, 9066–9071.
- Plovier, H.; Everard, A.; Druart, C.; Depommier, C.; Van Hul, M.; Geurts, L.; Chilloux, J.; Ottman, N.; Duparc, T.; Lichtenstein, L.; et al. A Purified Membrane Protein from Akkermansia Muciniphila or the Pasteurized Bacterium Improves Metabolism in Obese and Diabetic Mice. Nat. Med. 2017, 23, 107–113.
- Depommier, C.; Everard, A.; Druart, C.; Plovier, H.; Van Hul, M.; Vieira-Silva, S.; Falony, G.; Raes, J.; Maiter, D.; Delzenne, N.M.; et al. Supplementation with Akkermansia Muciniphila in Overweight and Obese Human Volunteers: A Proof-of-Concept Exploratory Study. Nat. Med. 2019, 25, 1096–1103.
- Goodrich, J.K.; Waters, J.L.; Poole, A.C.; Sutter, J.L.; Koren, O.; Blekhman, R.; Beaumont, M.; Van Treuren, W.; Knight, R.; Bell, J.T.; et al. Human Genetics Shape the Gut Microbiome. Cell 2014, 159, 789–799.
- Vajro, P.; Mandato, C.; Licenziati, M.R.; Franzese, A.; Vitale, D.F.; Lenta, S.; Caropreso, M.; Vallone, G.; Meli, R. Effects of Lactobacillus Rhamnosus Strain GG in Pediatric Obesity-Related Liver Disease. J. Pediatr. Gastroenterol. Nutr. 2011, 52, 740–743.
- Gøbel, R.J.; Larsen, N.; Jakobsen, M.; Mølgaard, C.; Michaelsen, K.F. Probiotics to Adolescents with Obesity: Effects on Inflammation and Metabolic Syndrome. J. Pediatr. Gastroenterol. Nutr. 2012, 55, 673–678.
- Marcelo, T.L.P.; Pellicciari, C.R.; Artioli, T.O.; Leiderman, D.B.D.; Gradinar, A.L.T.; Mimica, M.; Kochi, C. Probiotic Therapy Outcomes in Body Composition of Children and Adolescent with Obesity, a Nonrandomized Controlled Trial. Arch. Endocrinol. Metab. 2022, 66, 815–822.
- Chen, A.-C.; Fang, T.-J.; Ho, H.-H.; Chen, J.-F.; Kuo, Y.-W.; Huang, Y.-Y.; Tsai, S.-Y.; Wu, S.-F.; Lin, H.-C.; Yeh, Y.-T. A Multi-Strain Probiotic Blend Reshaped Obesity-Related Gut Dysbiosis and Improved Lipid Metabolism in Obese Children. Front. Nutr. 2022, 9, 922993.
- Verma, A.; Nelson, M.T.; DePaolo, W.R.; Hampe, C.; Roth, C.L. A Randomized Double-Blind Placebo Controlled Pilot Study of Probiotics in Adolescents with Severe Obesity. J. Diabetes Metab. Disord. 2021, 20, 1289–1300.
- Karlsson Videhult, F.; Öhlund, I.; Stenlund, H.; Hernell, O.; West, C.E. Probiotics during Weaning: A Follow-up Study on Effects on Body Composition and Metabolic Markers at School Age. Eur. J. Nutr. 2015, 54, 355–363.
- Dror, T.; Dickstein, Y.; Dubourg, G.; Paul, M. Microbiota Manipulation for Weight Change. Microb. Pathog. 2017, 106, 146–161.
- Luoto, R.; Kalliomäki, M.; Laitinen, K.; Isolauri, E. The Impact of Perinatal Probiotic Intervention on the Development of Overweight and Obesity: Follow-up Study from Birth to 10 Years. Int. J. Obes. 2010, 34, 1531–1537.
- Mărginean, C.; Mărginean, C.O.; Iancu, M.; Szabo, B.; Cucerea, M.; Melit, L.E.; Crauciuc, A.; Bănescu, C. The Role of TGF-B1 869 T>C and PPAR Γ2 34 C>G Polymorphisms, Fat Mass, and Anthropometric Characteristics in Predicting Childhood Obesity at Birth: A Cross-Sectional Study According the Parental Characteristics and Newborn’s Risk for Child. Obes.ity (the Newborns Obesity’s Risk) NOR Study. Medicine 2016, 95, e4265.
- Mărginean, C.; Mărginean, C.O.; Iancu, M.; Meliţ, L.E.; Tripon, F.; Bănescu, C. The FTO Rs9939609 and LEPR Rs1137101 Mothers-Newborns Gene Polymorphisms and Maternal Fat Mass Index Effects on Anthropometric Characteristics in Newborns: A Cross-Sectional Study on Mothers-Newborns Gene Polymorphisms-The FTO-LEPR Study (STROBE-Compliant Article). Medicine 2016, 95, e5551.
- Mărginean, C.O.; Mărginean, C.; Bănescu, C.; Meliţ, L.E.; Tripon, F.; Iancu, M. The Relationship between MMP9 and ADRA2A Gene Polymorphisms and Mothers-Newborns’ Nutritional Status: An Exploratory Path Model (STROBE Compliant Article). Pediatr. Res. 2019, 85, 822–829.
- Saros, L.; Vahlberg, T.; Koivuniemi, E.; Houttu, N.; Niinikoski, H.; Tertti, K.; Laitinen, K. Fish Oil And/Or Probiotics Intervention in Overweight/Obese Pregnant Women and Overweight Risk in 24-Month-Old Children. J. Pediatr. Gastroenterol. Nutr. 2023, 76, 218–226.
- Vähämiko, S.; Laiho, A.; Lund, R.; Isolauri, E.; Salminen, S.; Laitinen, K. The Impact of Probiotic Supplementation during Pregnancy on DNA Methylation of Obesity-Related Genes in Mothers and Their Children. Eur. J. Nutr. 2019, 58, 367–377.
- Oddi, S.; Huber, P.; Rocha Faria Duque, A.L.; Vinderola, G.; Sivieri, K. Breast-Milk Derived Potential Probiotics as Strategy for the Management of Childhood Obesity. Food Res. Int. 2020, 137, 109673.
- Markowiak, P.; Śliżewska, K. Effects of Probiotics, Prebiotics, and Synbiotics on Human Health. Nutrients 2017, 9, 1021.
- Gurry, T. Synbiotic Approaches to Human Health and Well-Being. Microb. Biotechnol. 2017, 10, 1070–1073.
- Atazadegan, M.A.; Heidari-Beni, M.; Entezari, M.H.; Sharifianjazi, F.; Kelishadi, R. Effects of Synbiotic Supplementation on Anthropometric Indices and Body Composition in Overweight or Obese Children and Adolescents: A Randomized, Double-Blind, Placebo-Controlled Clinical Trial. World J. Pediatr. 2023, 19, 356–365.
- Ipar, N.; Aydogdu, S.D.; Yildirim, G.K.; Inal, M.; Gies, I.; Vandenplas, Y.; Dinleyici, E.C. Effects of Synbiotic on Anthropometry, Lipid Profile and Oxidative Stress in Obese Children. Benef. Microbes 2015, 6, 775–782.
- Mohammadi, H.; Ghavami, A.; Hadi, A.; Askari, G.; Symonds, M.; Miraghajani, M. Effects of Pro-/Synbiotic Supplementation on Anthropometric and Metabolic Indices in Overweight or Obese Children and Adolescents: A Systematic Review and Meta-Analysis. Complement. Ther. Med. 2019, 44, 269–276.
- Kilic Yildirim, G.; Dinleyici, M.; Vandenplas, Y.; Dinleyici, E.C. Effects of Multispecies Synbiotic Supplementation on Anthropometric Measurements, Glucose and Lipid Parameters in Children With Exogenous Obesity: A Randomized, Double Blind, Placebo-Controlled Clinical Trial (Probesity-2 Trial). Front. Nutr. 2022, 9, 898037.
- Hadi, A.; Arab, A.; Khalesi, S.; Rafie, N.; Kafeshani, M.; Kazemi, M. Effects of Probiotic Supplementation on Anthropometric and Metabolic Characteristics in Adults with Metabolic Syndrome: A Systematic Review and Meta-Analysis of Randomized Clinical Trials. Clin. Nutr. 2021, 40, 4662–4673.
- Hadi, A.; Alizadeh, K.; Hajianfar, H.; Mohammadi, H.; Miraghajani, M. Efficacy of Synbiotic Supplementation in Obesity Treatment: A Systematic Review and Meta-Analysis of Clinical Trials. Crit. Rev. Food Sci. Nutr. 2020, 60, 584–596.
- Anderson, E.L.; Howe, L.D.; Jones, H.E.; Higgins, J.P.T.; Lawlor, D.A.; Fraser, A. The Prevalence of Non-Alcoholic Fatty Liver Disease in Children and Adolescents: A Systematic Review and Meta-Analysis. PLoS ONE 2015, 10, e0140908.
- Puri, P.; Sanyal, A.J. Nonalcoholic Fatty Liver Disease: Definitions, Risk Factors, and Workup. Clin. Liver Dis. 2012, 1, 99–103.
- Chalasani, N.; Younossi, Z.; Lavine, J.E.; Charlton, M.; Cusi, K.; Rinella, M.; Harrison, S.A.; Brunt, E.M.; Sanyal, A.J. The Diagnosis and Management of Nonalcoholic Fatty Liver Disease: Practice Guidance from the American Association for the Study of Liver Diseases. Hepatology 2018, 67, 328–357.
- Miele, L.; Marrone, G.; Lauritano, C.; Cefalo, C.; Gasbarrini, A.; Day, C.; Grieco, A. Gut-Liver Axis and Microbiota in NAFLD: Insight Pathophysiology for Novel Therapeutic Target. Curr. Pharm. Des. 2013, 19, 5314–5324.
- Chen, J.; Vitetta, L. Gut Microbiota Metabolites in NAFLD Pathogenesis and Therapeutic Implications. Int. J. Mol. Sci. 2020, 21, 5214.
- Loomba, R.; Seguritan, V.; Li, W.; Long, T.; Klitgord, N.; Bhatt, A.; Dulai, P.S.; Caussy, C.; Bettencourt, R.; Highlander, S.K.; et al. Gut Microbiome-Based Metagenomic Signature for Non-Invasive Detection of Advanced Fibrosis in Human Nonalcoholic Fatty Liver Disease. Cell Metab. 2017, 25, 1054–1062.e5.
- Alisi, A.; Bedogni, G.; Baviera, G.; Giorgio, V.; Porro, E.; Paris, C.; Giammaria, P.; Reali, L.; Anania, F.; Nobili, V. Randomised Clinical Trial: The Beneficial Effects of VSL#3 in Obese Children with Non-Alcoholic Steatohepatitis. Aliment. Pharmacol. Ther. 2014, 39, 1276–1285.
- Famouri, F.; Shariat, Z.; Hashemipour, M.; Keikha, M.; Kelishadi, R. Effects of Probiotics on Nonalcoholic Fatty Liver Disease in Obese Children and Adolescents. J. Pediatr. Gastroenterol. Nutr. 2017, 64, 413–417.
- Rodrigo, T.; Dulani, S.; Nimali Seneviratne, S.; De Silva, A.P.; Fernando, J.; De Silva, H.J.; Jayasekera; Wickramasinghe, V.P. Effects of Probiotics Combined with Dietary and Lifestyle Modification on Clinical, Biochemical, and Radiological Parameters in Obese Children with Nonalcoholic Fatty Liver Disease/Nonalcoholic Steatohepatitis: A Randomized Clinical Trial. Clin. Exp. Pediatr. 2022, 65, 304–311.
- Lin, H.; Yip, T.C.-F.; Zhang, X.; Li, G.; Tse, Y.-K.; Hui, V.W.-K.; Liang, L.Y.; Lai, J.C.-T.; Chan, S.L.; Chan, H.L.-Y.; et al. Age and the Relative Importance of Liver-Related Deaths in Nonalcoholic Fatty Liver Disease. Hepatology 2023, 77, 573–584.
- Loomba, R.; Sanyal, A.J. The Global NAFLD Epidemic. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 686–690.
- Tailor, A.M.; Peeters, P.H.M.; Norat, T.; Vineis, P.; Romaguera, D. An Update on the Prevalence of the Metabolic Syndrome in Children and Adolescents. Int. J. Pediatr. Obes. 2010, 5, 202–213.
- Friend, A.; Craig, L.; Turner, S. The Prevalence of Metabolic Syndrome in Children: A Systematic Review of the Literature. Metab. Syndr. Relat. Disord. 2013, 11, 71–80.
- Fiore, G.; Pascuzzi, M.C.; Di Profio, E.; Corsello, A.; Agostinelli, M.; La Mendola, A.; Milanta, C.; Campoy, C.; Calcaterra, V.; Zuccotti, G.; et al. Bioactive Compounds in Childhood Obesity and Associated Metabolic Complications: Current Evidence, Controversies and Perspectives. Pharmacol. Res. 2023, 187, 106599.
- Abrahamsson, T.R.; Sinkiewicz, G.; Jakobsson, T.; Fredrikson, M.; Björkstén, B. Probiotic Lactobacilli in Breast Milk and Infant Stool in Relation to Oral Intake during the First Year of Life. J. Pediatr. Gastroenterol. Nutr. 2009, 49, 349–354.
- Laitinen, K.; Poussa, T.; Isolauri, E. Nutrition, Allergy, Mucosal Immunology and Intestinal Microbiota Group Probiotics and Dietary Counselling Contribute to Glucose Regulation during and after Pregnancy: A Randomised Controlled Trial. Br. J. Nutr. 2009, 101, 1679–1687.
- Rautava, S.; Collado, M.C.; Salminen, S.; Isolauri, E. Probiotics Modulate Host-Microbe Interaction in the Placenta and Fetal Gut: A Randomized, Double-Blind, Placebo-Controlled Trial. Neonatology 2012, 102, 178–184.
- Ilmonen, J.; Isolauri, E.; Poussa, T.; Laitinen, K. Impact of Dietary Counselling and Probiotic Intervention on Maternal Anthropometric Measurements during and after Pregnancy: A Randomized Placebo-Controlled Trial. Clin. Nutr. 2011, 30, 156–164.
- Benítez-Páez, A.; Gómez Del Pugar, E.M.; López-Almela, I.; Moya-Pérez, Á.; Codoñer-Franch, P.; Sanz, Y. Depletion of Blautia Species in the Microbiota of Obese Children Relates to Intestinal Inflammation and Metabolic Phenotype Worsening. mSystems 2020, 5, e00857-19.
- Tuovinen, E.; Keto, J.; Nikkilä, J.; Mättö, J.; Lähteenmäki, K. Cytokine Response of Human Mononuclear Cells Induced by Intestinal Clostridium Species. Anaerobe 2013, 19, 70–76.
- Benítez-Páez, A.; Kjølbæk, L.; Gómez Del Pulgar, E.M.; Brahe, L.K.; Astrup, A.; Matysik, S.; Schött, H.-F.; Krautbauer, S.; Liebisch, G.; Boberska, J.; et al. A Multi-Omics Approach to Unraveling the Microbiome-Mediated Effects of Arabinoxylan Oligosaccharides in Overweight Humans. mSystems 2019, 4, e00209-19.
- Kjølbæk, L.; Benítez-Páez, A.; Gómez Del Pulgar, E.M.; Brahe, L.K.; Liebisch, G.; Matysik, S.; Rampelli, S.; Vermeiren, J.; Brigidi, P.; Larsen, L.H.; et al. Arabinoxylan Oligosaccharides and Polyunsaturated Fatty Acid Effects on Gut Microbiota and Metabolic Markers in Overweight Individuals with Signs of Metabolic Syndrome: A Randomized Cross-over Trial. Clin. Nutr. 2020, 39, 67–79.
- Solito, A.; Bozzi Cionci, N.; Calgaro, M.; Caputo, M.; Vannini, L.; Hasballa, I.; Archero, F.; Giglione, E.; Ricotti, R.; Walker, G.E.; et al. Supplementation with Bifidobacterium Breve BR03 and B632 Strains Improved Insulin Sensitivity in Children and Adolescents with Obesity in a Cross-over, Randomized Double-Blind Placebo-Controlled Trial. Clin. Nutr. 2021, 40, 4585–4594.
- Sanchis-Chordà, J.; Del Pulgar, E.M.G.; Carrasco-Luna, J.; Benítez-Páez, A.; Sanz, Y.; Codoñer-Franch, P. Bifidobacterium Pseudocatenulatum CECT 7765 Supplementation Improves Inflammatory Status in Insulin-Resistant Obese Children. Eur. J. Nutr. 2019, 58, 2789–2800.
More