Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Emanuele Marzetti and Version 2 by Rita Xu.
Mitophagy is crucial for maintaining mitochondrial quality. The endosomal–lysosomal system is a more accessible pathway through which subtypes of extracellular vesicles, which also contain mitochondrial constituents, are released for disposal. The inclusion of mitochondrial components into extracellular vesicles occurs in the setting of mild mitochondrial damage and during impairment of lysosomal function. By releasing mitochondrial-derived vesicles, cells limit the unload of mitochondrial damage-associated molecular patterns with proinflammatory activity.
- damage-associated molecular patterns (DAMPs)
- endosomal–lysosomal system
- exosomes
- extracellular vesicles
1. Introduction
Mitophagy is a major mechanism of mitochondrial quality control. Mitophagy involves the degradation and recycling of damaged or inefficient mitochondria to maintain a healthy pool of organelles and ensure adequate energy provision to cells [1]. However, its assessment in vivo is challenging, which hampers its translational applications [2].
The endosomal–lysosomal system encompasses organelles and membranous components that form the endocytic pathway. Through the endosomal–lysosomal system, various cargo molecules are internalized and recycled. Furthermore, this pathway has emerged as a relevant and more accessible component of the mitochondrial quality control system [3][4][3,4]. Exosomes, a subtype of extracellular vesicles (EVs) with a diameter of 50–150 nm, are generated and released by the endosomal–lysosomal system. Exosomes are produced from intraluminal vesicles (ILVs), which originate from early endosomes undergoing inward budding of discrete membrane domains that subsequently evolve into multivesicular bodies (MVBs) [5][6][7][8][5,6,7,8]. MVBs are usually addressed to lysosomes for cargo degradation and recycling. However, under specific stimulation, MVBs can be shuttled toward the plasma membrane for exocytic fusion and subsequent extracellular release of ILVs [5][6][5,6]. Under these circumstances, ILVs are called exosomes.
Growing evidence has shown that, depending on the severity of mitochondrial damage, cells can selectively target mitochondrial components for lysosomal degradation and regulate the packaging of mitochondrial constituents into EVs [9]. Mitochondrial disposal via EVs seems to be an alternative strategy to canonical organelle degradation and recycling by mitophagy [9]. The EV pathway of mitochondrial quality control is triggered in the setting of mild mitochondrial damage to avoid wholesale organelle disposal while preventing the release of unpackaged damaged mitochondrial components. The latter would otherwise act as proinflammatory damage-associated molecular patterns (DAMPs) and install an inflammatory milieu [9]. Mitochondrial clearance has also been shown to be accomplished via the endosomal pathway and large EV generation when lysosomal degradation is inhibited [10].
The generation of mitochondrial-derived vesicles (MDVs), small vesicles that shuttle mitochondrial constituents to other organelles, is accomplished through the sorting of mitochondrial components via two different pathways. The first involves the delivery of mitochondrial material to EVs through sorting nexin 9 (SNX9)-dependent MDVs [11]. This vesicle class has been shown to participate in mitochondrial antigen presentation [11]. The second pathway targets MDVs containing damaged mitochondrial components to lysosomes for degradation in a process that is regulated by the mitophagy mediator Parkin [11]. Therefore, the characterization of MDV subtypes represents a unique tool for investigating the dynamics of mitochondrial quality control in health and disease [12]. MDVs also enable intercellular communication with either beneficial or harmful effects on recipient cells, depending on the cellular source as well as the nature of the cargo and the originating stimulus [13]. To further complicate the matter, a distinct population of EVs containing mitochondrial material—mitovesicles—has recently been described and attributed specific signaling functions [14]. All these factors contribute to a limited exploitation of MDVs in the clinical setting, for which a deeper characterization of MDV biology is needed [15].
2. Mitochondrial-Derived Vesicles
First identified in 2008 by Neuspiel et al. [16], MDVs are single- or double-membrane vesicles, generated, respectively, from the outer mitochondrial membrane (OMM) or inner mitochondrial membrane (IMM) and portions of the mitochondrial matrix. Three independent criteria are used for specifically defining MDVs: (i) generation independent of dynamin-related protein 1 (DRP1); (ii) small size (diameter of 70–150 nm); and (iii) selectivity of the cargo [16][17][16,17]. Mitochondria have inherited, from their bacterial ancestors, the ability to load their contents into vesicles for long-distance delivery as a constitutive and evolutionarily conserved process [18][19][18,19]. These similar prokaryotic vesicles are generated by local evaginations of mitochondrial membranes, vesicle budding, and release into the extracellular compartment. Bacteria release two main types of EVs: outer membrane vesicles (OMVs), generated from the outer plasma membrane layer, and outer–inner membrane vesicles (O–IMVs), containing both outer and inner membrane layer components [20]. OMVs mainly include bacterial lipopolysaccharides (LPSs) from the outer layer, peptidoglycans of the periplasmic space, and unstructured and organelle-free cytosol [21][22][21,22]. Conversely, ATP molecules, DNA, and proteins from the cytoplasm and the inner membrane are mostly included in O–IMVs [21][22][21,22]. EVs are implicated in multiple bacterial activities, from intracolonial signaling via quorum sensing [23] to intercellular communication via proteins shuttling, as well as in the modulation of immunogenic host invasion, elimination of bacterial competitors, and formation of biofilms [24]. MDVs in eukaryotic cells have more specialized roles compared with their prokaryotic counterparts and have evolved for different purposes [18]. In the setting of early mitochondrial stress, MDVs can shuttle oxidized cargo to lysosomes for degradation, or unload it in the extracellular compartment via EVs, thereby regulating mitochondrial mass more rapidly than mitophagy [3][4][9][3,4,9]. MDVs are highly heterogeneous, and several subtypes have been identified, which also depend on the cell type [25][26][27][25,26,27]. Their heterogeneity is reflected by the multiple processes in which they are involved, including mitochondrial fission, biogenesis of peroxisomes, resistance to oxidative stress and infections, and innate immunity signaling [11][28][29][30][31][11,28,29,30,31]. Alterations in the mechanisms generating MDVs have been associated with several pathological conditions, such as neurodegeneration and cardiomyocyte damage under ischemia/hypoxia [4][32][33][4,32,33]. Moreover, altered MDV generation and release have been associated with aging, autoimmune diseases, cancer, and infections [11][34][35][11,34,35]. MDV generation is a housekeeping process that occurs at a basal level under physiological conditions [25][36][25,36] and is enhanced during pathological stress. An increase in the production of reactive oxygen species (ROS) inflicts damage on proteins, lipids, and nucleic acids, and cells can trigger MDV formation to allow clearance of abnormal mitochondrial particles [28][36][37][28,36,37]. In the setting of mild oxidative stress, the oxidation of mitochondrial membrane proteins initiates the local activation of phosphatase and tensin homologue (PTEN)-induced putative kinase 1 (PINK1)/Parkin, guiding the budding of oxidized membrane proteins and their inclusion into vesicles [4][11][4,11]. Under physiological conditions, PINK1 is constitutively imported through the mitochondrial import channel of the outer membrane and, when inside the organelle, is cleaved by the protease of the inner membrane (PARL) [38]. Upon cleavage, PINK1 is retro-translocated to the cytoplasm for rapid proteasomal degradation [39], while Parkin resides in the cytosol in the form of an autoinhibited E3 ubiquitin-ligase [33]. Under mild stress, mitochondrial depolarization disrupts the activity of mitochondrial import channels and blocks the internalization of PINK1 that becomes stalled at the import channel or at the outer mitochondrial membrane [40][41][40,41]. Here, PINK1 undergoes autophosphorylation and, in turn, phosphorylates the ubiquitin protein and the ubiquitin-like domains of Parkin. These events stabilize Parkin in an active state and facilitate MDV generation and release [40][41][40,41]. Matheoud et al. [11] have shown that the biogenesis of MDVs also requires recruitment of the Ras-related protein (Rab9) and SNX9, although the regulation of this process remains unclear. Rab9 is a small GTPase located at the trans-Golgi network and late endosomes, and is implicated in pathways regulating the endo−lysosomal trafficking [42]. Conversely, SNX9 binds directly to the regulator of endocytosis, mediated by the protein clathrin [43]. This pathway generates MDVs for mediating mitochondrial antigen presentation after proteasomal breaking of mitochondrial components into lysosomes and loading of mitochondrial antigens onto major histocompatibility complex class I (MHC I) molecules at the endoplasmic reticulum for their subsequent transfer to cell surface [11][44][45][11,44,45] (Figure 1). Through this pathway, MDVs can regulate survival, development, activation, and differentiation of immune cells (e.g., T-lymphocytes and macrophages) [44][46][47][48][44,46,47,48]. The destination of MDVs to MVBs for final lysosomal fusion and degradation has also been described as a mechanism to selectively package and dispose mitochondrial constituents via EVs [28][49][28,49]. This process prevents the release of oxidized mitochondrial DAMPs with proinflammatory properties [28][49][28,49].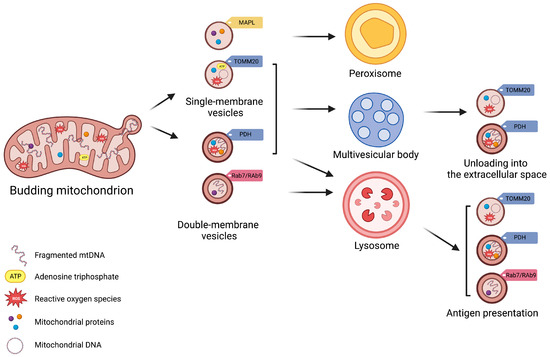
Figure 1. Schematic Representation of Subtypes of Mitochondrial-Derived Vesicles. Mitochondrial-derived vesicles (MDVs) can be classified according to their membrane composition and cargo selection. Based on membrane structure and composition, MDVs can be distinguished as single- or double-membrane vesicles. Single-membrane MDVs incorporate outer mitochondrial membrane proteins, while double-membrane MDVs include outer and inner mitochondrial membrane proteins and constituents of the mitochondrial matrix. According to cargo and membrane protein markers, MDV subtypes include single-membrane MDVs that bear mitochondrial-anchored protein ligase (MAPL) and the import channel translocase of the outer mitochondrial membrane complex subunit 20 (TOMM20), and double-membrane MDVs with pyruvate dehydrogenase (PDH). Vesicle subtypes follow different degradative pathways. Peroxisome is the final destination of single-membrane MDVs with MAPL, while those with TOMM are excreted via multivesicular bodies (MVBs) as exosomes. Double-membrane MDVs with PDH are also released via MVBs. Finally, Ras-related protein (Rab) 7-/9-guided MDV generation mediate mitochondrial antigen presentation via major histocompatibility complex (MHC) class I.