Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Prakash Pandey and Version 3 by Rita Xu.
Living organisms rely on pH levels for a multitude of crucial biological processes, such as the digestion of food and the facilitation of enzymatic reactions. Among these organisms, animals, including insects, possess specialized taste organs that enable them to discern between acidic and alkaline substances present in their food sources.
- taste
- acid
- alkali
- OTOP1
- Alkaliphile
1. Introduction
Sensory perception plays a vital role in the survival and well-being of animals, enabling them to navigate their environment and meet their fundamental requirements, such as securing nourishment, finding shelter, ensuring reproductive success, ensuring safety, and engaging in meaningful interactions with other members of their ecosystem. This complex procedure requires the synchronization of multiple separate sensory systems, each skilled in transmitting vital data to the brain for prompt analysis and reaction. Among these, mammals have evolved five primary sensory organs, namely the eye for vision, the ear for hearing, the tongue for taste, the nose for smell, and the skin for tactile perception, collectively facilitating the discernment of visual cues, auditory signals, flavors, scents, and tactile sensations. Much like other mammals, Drosophila melanogaster possesses distinct sensory organs that empower it to distinguish favorable from unfavorable environmental stimuli, enabling its survival. Drosophila exhibits a sophisticated sensory apparatus that aids in the recognition of external stimuli. Their remarkable compound eyes are particularly adept at facilitating various aspects of visual behavior, serving as a pivotal tool for environmental navigation and identification [1]. Furthermore, the species relies on the Johnston’s organ, situated within the antenna, which serves the dual purpose of detecting sounds and facilitating mechanosensation, thereby allowing fruit flies to respond to auditory and mechanical stimuli with precision [2][3][4][5][2,3,4,5]. In addition to these, Drosophila possesses a diverse array of olfactory organs, each specialized in the detection and processing of a wide range of odors [6]. These olfactory receptors play a critical role in the fly’s ability to identify and respond to specific volatile chemical cues within their surroundings. Moreover, the gustatory organs of the fruit fly enable the perception of taste, allowing them to differentiate between various food sources and potentially harmful substances [7][8][9][10][11][12][7,8,9,10,11,12]. This intricate sensory system collectively equips Drosophila with the tools necessary for efficient perception and response to the external environment. Within the adult body of Drosophila, hair-like projections develop into taste organs in various locations, including the proboscis, legs, wing margins, and ovipositor [13][14][15][16][17][18][19][13,14,15,16,17,18,19]. Of these, the tip of the proboscis comprises the bifurcate labellum, which plays a particularly crucial role in detecting taste by coming into contact with food or chemical compounds. The labellum of Drosophila contains 31 taste sensilla on each side, arranged symmetrically. These sensilla are instrumental in chemosensations, especially concerning non-volatile compounds. There are three distinct types of sensilla in the labellum, short (S-type), intermediate (I-type), and long (L-type), differentiated by their size [16][20][21][16,20,21]. Each sensillum is innervated by two to four gustatory receptor neurons (GRNs), one mechanosensory neuron, and three supporting cells, with their signals projected to the subesophageal zone (SEZ) of the brain, the region responsible for taste sensation [13][22][23][24][13,22,23,24]. Additionally, taste sensation is also mediated by the hairless labellar taste peg situated between pseudotracheal rows, which is innervated by one chemosensory neuron and one mechanosensory neuron [25]. It is believed that taste pegs can only detect food when the flies open their labial palps. Furthermore, the adult fly pharynx, which acts as an internal molecular sensor, contains three different hairless internal taste organs: the dorsal cibarial sense organ (DCSO), the ventral cibarial sense organ (VCSO), and the labral sense organ (LSO) [26].
More precisely, the presence of specific molecular components like gustatory receptors (GRs) [7][27][7,27], ionotropic receptors (IRs) [28][29][28,29], pickpocket (PPK) ion channels [30][31][32][30,31,32], and transient receptor potential (TRP) ion channels [33][34][33,34] acts as mediators between external chemical cues and the fruit fly’s brain. These components play a crucial role in transducing chemical information into neural signals. The GRNs present in sensilla are sensitive to different types of chemical stimuli entering via pores at the tip of the sensilla. For example, the S-type sensilla contains four different GRNs, each responding to bitter or aversive compounds, sweet tastes, water, or specific minerals such as Na+ and Ca2+ [13][18][19][32][13,18,19,32]. In contrast, the L-type sensilla possesses four sets of GRNs that are sensitive to sweet tastes, water, low concentrations of salt, and others which have not yet been identified [19][35][36][37][38][19,35,36,37,38]. Similarly, the I-type sensilla are equipped with two sets of GRNs responding to sweet and bitter compounds, including both low and high concentrations of salt [39][40][41][39,40,41]. In this intricate web of molecular and sensory interactions, Drosophila’s taste organs play a pivotal role in helping the fly navigate its environment, ensuring its sustenance and survival. These sensory mechanisms provide a fascinating window into the broader world of biological systems and their adaptation to complex ecological challenges.
Taste preferences in Drosophila also have profound ecological implications. They guide foraging behavior, influence food source selection, impact breeding site choices, and even contribute to competition and niche partitioning among different Drosophila populations. Drosophila’s taste neurons and receptors assess food sources, shaping their preferences and guiding foraging. Favorable food sources lead to concentrated fly populations, impacting resource distribution. Drosophila can also act as pollinators, spreading pollen from one plant to another as they feed on nectar, affecting plant ecology. Taste cues guide females to identify and select appropriate substrates for oviposition. Consequently, taste preferences influence the distribution of Drosophila larvae within their environment, impacting their development and survival. Diverse taste perception facilitates niche partitioning, enabling the coexistence of multiple Drosophila species by exploiting different food sources and reducing competition. Understanding these taste preferences is essential for unraveling the intricate ecological dynamics and interactions between these fruit flies and their environment.
Drosophila’s food preferences go beyond taste alone, influenced by nutritional needs, genetic variation, environmental conditions, and learned behavior. While taste guides their initial preferences, nutritional requirements can override taste, and genetic diversity affects their perception of specific compounds [42][43][51,52]. Environmental factors, like food availability and competition, also play a significant role. Drosophila can learn from experience, allowing them to adapt and make optimal food choices. These multifaceted factors ensure their adaptability and reproductive success in diverse environments.
Sourness, characterized by low pH or acidity, represents a fundamental taste sensation [44][53]. It is universally appealing in moderate concentrations, yet becomes unappealing at higher levels, across both vertebrate and invertebrate species [45][46][47][48][54,55,56,57]. Conversely, foods with high pH or alkaline properties generally lack appeal [49][50][51][58,59,60]. Nevertheless, in mildly alkaline conditions, food with compounds like ammonia and certain amines is preferred, influenced by the intricacies of the sensory system [52][53][61,62] (Figure 1A). Notably, the attraction to mildly basic food is believed to be linked to the presence of low concentrations of salt [49][58]. Commonly, acidic components are found in raw fruits and spoiled food items, adding to the significance of detecting sourness as a warning sign. Conversely, basic or alkaline taste is triggered in certain vegetables, legumes, nuts, and other items due to their elevated pH levels. Current research across various species, both vertebrate and invertebrate, has substantiated that basic taste also qualifies as one of the fundamental taste qualities [49][51][54][55][56][58,60,63,64,65].
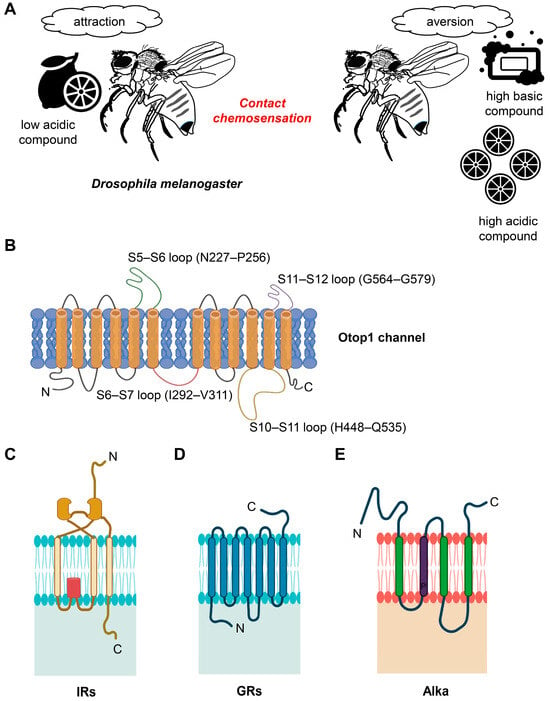
Figure 1. Behavior of acid and basic compounds and structure of OTOP1, GRs, IRs, and Alka. (A) Representation of D. melanogaster’s sensitivity to pH Levels. D. melanogaster is attracted to compounds with low acidity (low pH) while displaying aversion to highly acidic and highly basic compounds (high pH). (B) Human Otop1 channel membrane configuration. The human Otop1 channel is thought to have a membrane-spanning structure, with the N domain encompassing transmembrane segments S1 through S6, the C domain including S7 to S12, interconnected loops between these segments, and intracellular termini. (C) Architectural features of GRs. GRs possess a unique architectural composition characterized by seven transmembrane domains (TMDs), with their C-terminal regions located externally, distinguishing them from typical G-protein coupled receptors. (D) Topology of IRs. IRs exhibit a three-TMD structure, including a pore region, which shares structural similarities with mammalian glutamate receptors. (E) Proposed structural arrangement of alka. Alka is anticipated to have a structural configuration that includes a persistent proline residue (P) within the TM2 segment.