Shiga-toxin-producing Escherichia coli (STEC) is typically detected on food products, mainly due to cross-contamination with faecal matter. The serotype O157:H7 has been of major public health concern due to the severity of illness caused, prevalence, and management. In the food chain, the main methods of controlling contamination by foodborne pathogens often involve the application of antimicrobial agents, which are now becoming less efficient. There is a growing need for the development of new approaches to combat these pathogens, especially those that harbour antimicrobial resistant and virulent determinants. Strategies to also limit their presence on food contact surfaces and food matrices are needed to prevent their transmission. SRecent studies have revealed that bacteriophages are useful non-antibiotic options for biocontrol of E. coli O157:H7 in both animals and humans. Phage biocontrol can significantly reduce E. coli O157:H7, thereby improving food safety. However, before being certified as potential biocontrol agents, the safety of the phage candidates must be resolved to satisfy regulatory standards, particularly regarding phage resistance, antigenic properties, and toxigenic properties.
- food borne infection
- antimicrobial resistance
- Escherichia coli O157:H7
- phage thera
1. Introduction
2. Evolution, Virulence and Pathogenicity of E. coli O157:H7
Non-toxigenic and less virulent E. coli O55:H7 strains are known to be ancestral cells of E. coli O157:H7 [24][25][36,37] that originated through a series of transitional phases [26][38]. According to the concept model, the locus of enterocyte effacement (LEE) was present in both the O55:H7 and O157:H7 strains, and it is capable of inducing diarrhoea through an attachment–effacement process [27][28][29][39,40,41]. Four sequential events that led to the emergence of E. coli O157:H7 include (i) acquisition of an stx operon, which is encoded in the genome of lambdoid prophages, (ii) acquisition of the rfb region encoding the O157 antigen, and (iii) loss of β-glucuronidase (GUD) activity [30][31][42,43]. Notably, the three main virulence factors of E. coli O157:H7 include (i) Shiga toxin operons, (ii) locus of enterocyte effacement, and (iii) byproducts of the F-like plasmid pO157 [32][33][44,45]. Based on the degree of its virulence, the Shiga toxin operon is divided into two major groups, namely, stx1 and stx2 genes [34][46]. Interesting, the Stx1 has three subtypes (Stx1a, Stx1c, and Stx1d), while Stx2 has seven subtypes (Stx2a, Stx2b, Stx2c, Stx2d, Stx2e, Stx2f, and Stx2g). It is widely acknowledged that diverse stx subtypes possess the ability to bind to multiple receptors, exhibiting varying affinities for each receptor. The primary target for Stx2a is the Gb3 receptor, whereas Stx1a can bind to both Gb3 and globotetraosylceramide (Gb4). The Stx2e subtype exhibits a broader spectrum of receptor binding capabilities, including interactions with globopentaosylceramide, pentahexosylceramides featuring Gb4-elongated core structures, and Gb4 with a preference for Gb5 [35][47]. Several other genes, apart from stx, have been associated with the pathogenicity and virulence of O157:H7 [36][48].2.1. Transmission of E. coli O157:H7 to Humans and Animals
E. coli serotype O157 can be transmitted through different routes, including direct contact with animal droppings, ingestion of contaminated food and water, and transmission from one person/animal to another [5][37][5,49] (Figure 1).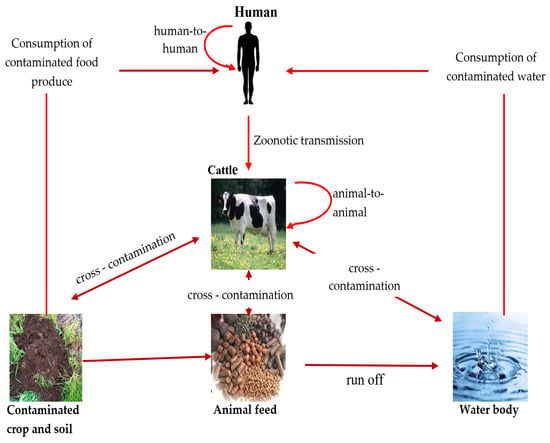
2.1.1. Contaminated Food as a Transmission Vector
Contaminated food plays a crucial role in the transmission of E. coli O157:H7 to humans [38][50]. Food products derived from animals (beef, lamb, chicken, pork, and ground beef) are considered to be the most common source of the pathogen and the transmission of the diseases amongst humans worldwide [39][51]. Studies have reported that E. coli O157:H7 illnesses have also been linked to the ingestion of other food products originating from cattle, such as unpasteurized milk and other dairy products [9][40][9,52]. In general, animal reservoirs are responsible for the greatest amount of E. coli O157:H7 transmission to humans, and food-related transmission is common, making it necessary to manage the pathogens in animal-derived foods, especially beef. As a result, E. coli O157:H7 poses a major risk to public health by endangering both human health and food safety.2.1.2. Contaminated Water as a Transmission Vector
The transmission of E. coli O157:H7 through water has been documented in various settings, including both drinking water that has been contaminated [41][56] and recreational water bodies [42][43][57,58]. Furthermore, studies conducted in Nigeria and South Africa demonstrated that water used for irrigation had notable influence on the dissemination of E. coli O157:H7 through the contamination of food products [44][45][59,60]. Untreated sewage released from hospitals, farms, and residential areas containing E. coli O157:H7 into nearby water bodies also increases the risk of human infections significantly [46][61]. Based on the evidence, E. coli O157:H7 easily survives in water and may persist for several weeks and even longer [43][58]. Strong rainstorms may cause sediments to be re-suspended, which could lead to an abrupt rise in E. coli O157:H7 concentrations in the water [47][62].2.1.3. Person-To-Person Transmission
The transmission of E. coli O157:H7 infection most often occurs through the faecal–oral route, primarily in settings where infected individuals come in close contact with others, such as in households, daycare centres, and healthcare facilities [48][49][64,65]. Children are more vulnerable than adults due to their immunological immaturity and limited understanding of appropriate hygiene practices [50][66].2.2. Epidemiology of E. coli O157:H7
According to Mesele and Abunna [51][67], E. coli O157:H7 is responsible for approximately 73,000 illnesses, 2000 hospitalizations, and 50–60 deaths annually in the United States. Reports that evaluated E. coli O157 outbreaks that occurred in the USA between 1982 and 2002 and were reported to the Centers for Disease Control and Prevention (CDC) showed 350 outbreaks, totalling 8598 cases and 1493 (17%) hospitalizations. A total of 354 (4%) presented with HUS and 40 (0.5%) died [52][68]. This highlights the significant public health potential of E. coli and also provides an overview of its epidemiology in the region. The varying aetiology of E. coli O157:H7 infections explains why the pathogen has also been isolated in humans, animals, food products, and the environment, especially in the African region [53][69], where public health policies are not adhered to strictly. Examples of documented cases in Africa are as follows: human infection that was reported in Johannesburg, South Africa in 1990 [54][70]; the detection of the pathogen in individuals with haemorrhagic colitis that resulted to fatalities in Bangui, Central African Republic [55][71]; and the isolation of the pathogen in East African countries (Tanzania, Kenya, and Ethiopia) [56][72].2.3. Treatment for E. coli O157:H7 Infections and Antimicrobial Resistance
Antibiotics are frequently used in animal production to prevent infection and as a growth promoter. This practice has made it difficult to treat infections caused by E. coli O157:H7, as the pathogens have become resistant to antibiotics. Given the virulence potential and pathogenicity of E. coli O157:H7, it is unfortunate that there is no specific treatment for E. coli O157:H7 infection [51][67]. In humans, antibiotics are typically not recommended because they can increase the risk of complications, such as HUS, a serious kidney disease. Although the spread of antimicrobial resistance in E. coli O157:H7 and other pathogens is being monitored and mitigated by surveillance [57][81], an alternative treatment method is necessary as a strategy to combat E. coli O157:H7 and infections caused by it, and also to serve as an additive to feed stock for the prevention of sicknesses. Bacteriophage therapy may be an effective alternative to antibiotic therapy for antibiotic-resistant E. coli O157:H7, both in humans and in animal production.3. Bacteriophages
Bacteriophages are viruses that exclusively infect bacteria. Although there were arguments regarding the discovery of phages, it has been acknowledged that the first scientists to discover the phages were William Twort and Felix d’Herelle [58][59][82,83]. Twort, an English medical bacteriologist, postulated that viruses were the causative agents of this phenomenon after observing a similar process in Micrococcus [60][61][62][84,85,86]. According to d’Herelle’s findings, the presence of an imperceptible microorganism was detected in stool samples obtained from dysentery patients, which were devoid of bacteria due to filtration [63][87]. The occurrence of clear zones observed in bacterial lawns can be attributed to the presence of “invisible germs”. D’Herelle provided the initial elucidation regarding the mechanism by which these viruses proliferate by exploiting bacteria, resulting in the death and subsequent disintegration of the bacteria. After their discovery, d’Herelle used phage preparations to treat bacterial dysentery [64][88]. Owing to its success in treating bacterial infection, the use phage therapy escalated in countries such as Tbilisi, Georgia, and Poland to treat conditions such as typhoid, fever, dysentery, surgical wound infections, peritonitis urinary tract infection, and septicaemia [65][89]. However, the use of phage therapy was halted by the discovery of the first antibiotic, penicillin, in 1940. In addition, lack of knowledge of phage therapy decreased the use of phage therapy. Despite this, Soviet countries continued to use phages to treat bacterial infections in humans. Bacteriophages are ubiquitously found in the vicinity of their host bacteria. The phage replication cycle encompasses the processes of adsorption, penetration, nucleic acid replication, and virion assembly. Consequently, the cellular membrane undergoes lysis, leading to the release of mature virions. While it is true that certain phages can infect multiple bacterial species or strains, most phages exhibit specificity towards a particular bacterial species or strain. The determination of specificity is regulated by the existence of receptors located on the outer surface of bacterial cells, such as lipopolysaccharide (LPS), flagella, and/or other surface proteins [66][90]. Bacteriophages can exhibit either a lytic or lysogenic life cycle, depending on the specific life cycle they adopt after infecting a host cell. Lytic phages undergo a lytic cycle, which starts when the bacteriophage binds to specific receptors on the surface of the bacterial cell. The phage then injects its DNA into the host cell and assumes control of the cellular machinery. While using the host cell’s transcription and translation mechanisms, phage-specific proteins are synthesized, and the phage’s DNA is replicated. The components then assemble into new phage particles, and they accumulate and exert pressure on the cell’s internal environment, causing the bacterial cell to lyse or rupture. In the lytic lifecycle of a bacteriophage, the lysis process is a crucial phase. It enables the newly produced phage particles to disseminate and infect other susceptible bacterial cells, ultimately resulting in the phage population increasing. Temperate phages, on the other hand, undergo the lysogenic cycle, wherein their genetic material integrates and coexists with the host genome in a stable manner, forming a prophage. This prophage replicates along with the host cell during replication. According to Pinto et al. [67][80], temperate phages have the ability to transition into the lytic cycle when they encounter cellular stress.4. Challenges Faced in the Application of E. coli O157:H7 Phages
There are a number of difficulties and factors to take into account when using phages as a biocontrol for E. coli O157:H7 either on food products or as a medication in humans and animals. Phages are very specific to individual bacterial strains [68][144]. Since E. coli O157:H7 strains can vary genetically and develop resistance to phages over time, it can be difficult to find phages that effectively target these strains. As a result, a combination of different phages in the form of a cocktail may be necessary for effective biocontrol of this pathogen. E. coli O157:H7 has a low infectious dosage (10 to 100 CFU/mL) and can survive in low-pH conditions, such as the acidic environment of the stomach and acidic food, which is one of its stress resistance mechanisms [69][145]. It can be therefore difficult to deliver phages to the site of infection because they may need to endure stomach acid and other digestive processes in order to reach the intestinal E. coli O157:H7 in the gut. For phages to be considered effective at inhibiting E. coli O157:H7, they must possess acid resistance in order to survive in the acidic environment of the stomach. Furthermore, phage products can be expensive to create, produce, and distribute due to their sensitivity to environmental factors like pH and temperature [70][146]; their quality and effectiveness can be compromised if they are transported and stored in unsuitable environmental conditions. The issue of storage and transportation has made it difficult for patients situated in low-resource environments to have access to them. Additionally, research has shown that phage therapy may be more effective when combined with other treatments, such as antibiotics [71][72][147,148]. Since its usage in humans could raise ethical concerns, it is crucial to carefully consider the best sequence and combination of therapies that will not cause further complications in infected patients. Inadequate clinical data also pose a challenge in the application of E. coli O157:H7 phages; while there have been some case reports and small clinical studies on phage therapy [73][149], more large-scale, randomised controlled trials are required to establish the safety and efficacy of phages as a biocontrol agent in treating E. coli O157:H7 infections.Phage Resistance by E. coli O157:H7
The evolution of phage resistance in E. coli O157:H7 is a continuous and natural process. As a result of repeated use of phages against E. coli O157:H7, in vitro experiments have shown that E. coli O157:H7 can develop phage resistance [74][150]. To avoid phage infection, they employ a variety of processes that involve the following techniques. Phage inability to attach: E. coli O157:H7 may alter the architecture of its cell surfaces to restrict phages from attaching to its surface [75][151]. To attach to particular receptors on the bacterial cell wall, phages often use receptor-binding proteins [76][152]. Phage attachment may be prevented by changes to these receptors. E. coli O157:H7 can change its surface characteristics, such as the lipopolysaccharide (LPS) layer, thereby making it challenging for phages to adhere to the bacterial cell [77][153]. CRISPR-Cas Systems: Some E. coli O157:H7 strains have Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR) and CRISPR-associated (CRISPR-Cas) systems, which are a form of adaptive immunity against phages. CRISPR-Cas systems play a significant role in limiting phage infection and proliferation as an important bacteriophage resistance mechanism [78][154]. By using these systems, bacteria can collect and preserve genetic material from earlier phage contacts and use this information to locate and eliminate the phage DNA when there is a contact with the same phage. Restriction–Modification (R-M) Systems: E. coli O157:H7 and other organisms may have R-M systems. These systems are made up of enzymes that protect bacterial cells by breaking down foreign DNA (including phage DNA) that enters the cell. They recognize and cleave phage DNA sequences on the recognition site [79][155], thereby inhibiting phage genetic material from multiplying. Abortive Infection: The infected E. coli strain employs abortive infection mechanisms, resulting in self-destruction prior to the completion of the phage’s replication cycle [80][156]. This behaviour effectively inhibits the phage epidemic from spreading to neighbouring cells, thus ensuring the safety of the bacterial colony. Understanding the mechanisms of phage resistance is critical in phage therapy. The mechanisms described above are strategies adopted by numerous bacterial populations to protect themselves from phage predation, and they can differ amongst strains of E. coli O157:H7. Due to the pressure exerted by phages, the pathogen can develop resistance to phages over time. As a result, phage cocktails can be used as a method to target multiple mechanisms or in combination with other treatment options to overcome bacterial resistance.5. Determining the Safety of E. coli O157:H7 Specific Bacteriophages as a Biocontrol Agent
To maximise the potential of phage treatment of E. coli O157:H7 infections, phage candidates must be safe and capable of serving as biocontrol agents without having negative effects on people or the environment. Firstly, phage’s specificity for E. coli O157:H7 must be ensured by isolating and properly characterising candidate phage; afterwards, phages should be screened for the existence of genes linked to virulence and antibiotic resistance within the phage’s genome simply because genes that encode toxins and allergens must not be present in phages intended for biocontrol as they can be detrimental to humans and animals [81][108]. New generation sequencing techniques like the Whole Genome Sequencing (WGS) can be used to analyse the phage genome and determine the presence of these genes. WGS is a method of typing that relies on sequencing of the entire genome of an isolate, allowing for the identification of variations at the level of individual nucleotides [82][157]. Based on the findings of the study conducted by Lee et al. [83][158], WGS was used to determine the absence of virulence factors, toxins, antibiotic resistance, and potential allergen-coding genes in the genome of the isolated phage KFS-EC. The phage which was obtained from wastewater samples from a slaughterhouse in Korea exhibits specificity exclusively towards E. coli O157:H7 and was considered to be suitable as a biocontrol agent. Having considered the genomic screening of phages, conducting animal studies to assess the safety of the phages in the target host organisms (e.g., livestock) is essential before a large scale production. Studies on the effectiveness of phages in treating experimentally infected animals [84][85][161,162] have demonstrated that using mice, chickens, or sheep in biocontrol trials will allow for the monitoring of any negative effects on animal health [86][163]. Clinical trial studies can evaluate the safety of phage therapy in people, including possible allergic reactions or other adverse effects. They can also determine the right concentration and dosage of phages required to control E. coli O157:H7 in people. Due to the sensitive nature and ethical issues associated with phage application, stakeholders like farmers, food processors, and healthcare professionals should obtain training on how to handle phages safely and responsibly in the real-world settings. Furthermore, while ensuring phage purity and quality during preparation, there is a possibility of generated residues and contaminants causing environmental hazards, which could pose a health risk. It is crucial to monitor the potential environmental impact of using phages, such as phage persistence in the environment and potential ecological disruptions.6. Conclusions
In the current era of antimicrobial resistance and the pursuit of alternative methods to combat E. coli O157:H7 infections, there has been a renewed interest in phage research. This is evident in the accumulated evidence suggesting that phage application is an effective method of biocontrol of foodborne bacterial pathogens on fresh produce and other foods. Phages has been used as a natural antimicrobial method to reduce E. coli O157:H7 from the food supply. Phage-based medications have been approved by the Food and Drug Administration (FDA) in certain countries, such as the United States, Canada, and Israel for the treatment of E. coli strains, including the use of various phage cocktails. Numerous studies have also produced novel phages and showed their effectiveness in inhibiting the growth of E. coli O157:H7 in food products without changing the organoleptic properties. They have also been proven to reduce E. coli O157:H7 in vivo through the use of mice. Ongoing research involves the construction of phage libraries, thereby enabling the use of phage cocktails to inhibit phage mutant strains, increase host range, and increase infectivity. Nevertheless, the European Union remains concerned about the utilization of these products due to the limited availability of safety data and comprehensive research on the potential consequences of phage discharge into the environment. One of the most important aspects of creating One Health approaches to lowering health risks for people, agricultural systems, and natural ecosystems is the effective use of “safe” phages as a biocontrol agent in combating pathogens like E. coli O157:H7 and resolving the issue of antibiotic resistance in preharvest livestock environments. In order to advance the development of novel antibacterial strategies, there is a need to conduct further investigation into the phage properties that make them safe and suitable as a biocontrol and the underlying mechanism through which phages can effectively inhibit the expression of host genes. The utilization of advanced technologies like WGS to carefully scrutinize and characterize phage genomes presents a solution to this concern. This, in turn, will improve both patient care and infection control strategies, ultimately leading to a decrease in the incidence of severe E. coli O157:H7 infections.