Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Mati Ur Rahman and Version 2 by Wendy Huang.
Plant-parasitic nematodes significantly threaten agriculture and forestry, causing various diseases. They cause annual losses of up to 178 billion dollars worldwide due to their parasitism. Nematophagous fungi (NF) are valuable in controlling or reducing parasitic nematode diseases by killing nematodes through predatory behavior.
- nematophagous fungi
- nematodes
- predatory fungi
- egg- and female-parasitic fungi
- endoparasitic fungi
- toxins
1. Introduction
Nematodes are highly ubiquitous animal groups found throughout the planet. Despite being over 28,000 known species, they are challenging to differentiate. Out of all these species, 16,000 are recognized as parasitic towards animals, insects, or plants [1]. Over 4000 species of plant-parasitic nematodes (PPNs) have been identified [2] which severely threaten agricultural and horticultural crops [3]. These harmful pathogens are hideous crop parasites that cause global annual losses between USD 78 and 125 billion [4][5][4,5]. PPNs can actively move in the soil region adjacent to the plant roots, known as the rhizosphere, as well as on the aerial parts of the plant, or within the plant itself. They feed and breed on living organisms, with some nematodes feeding on plant parts such as flowers, leaves, stems, and seeds [6]. However, most of these worms feed on underground plant parts like roots, bulbs, and tubers, causing substantial plant damage ranging from mild harm to destruction [5].NF are capable of capturing, killing, and digesting nematodes [7][35]. They reside externally and internally within the host organism, exploiting it for sustenance. These fungi use specific traps to ensnare prey, hyphae tips to parasitize females and eggs, and conidia to adhere while generating toxins to attack nematodes. Based on these strategies, NF are traditionally classified into four groups (Figure 1): (1) The group of fungi that prey on nematodes using specialized traps; (2) some fungi are egg-parasitic and invade nematode eggs or females through their hyphal tips, (3) while others are endoparasitic and use their spores. (4) Additionally, toxin-producing fungi immobilize nematodes before invading them [8][9][14,36].
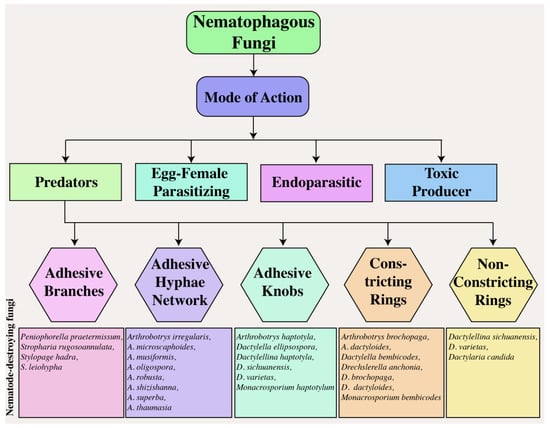
Figure 1.
Mode of action of nematophagous fungi.
2. Predatory Fungi
Predatory fungi employ hyphal structures in nematodes. Studies on nematode-trapping fungi have drawn much attention to their diverse and intricate catching structures [10][37]. The traps generated by the fungi’s mycelium adversely affect the nematode’s cuticle. The mycelium proliferates within the nematode’s body, resulting in the formation of a penetration peg, whose growth over time causes the hyphae to cover the outer surface of the colonized nematode [11][38]. The fungus A. oligospora has a unique mechanism for penetrating the cuticle of nematodes via penetrating tubes, and its impact on Meloidogyne javanica in tomato cultivars has been empirically demonstrated [12][39]. Predatory structures are vital for the life and activity of trapping fungi. Compared to regular hyphae, adhesive traps have a longer lifespan [13][14][40,41]. These specialized traps are utilized by over two hundred fungal species (found within the Zygomycota, Basidiomycota, and Ascomycota phyla) to capture free-living nematodes in soil [15][42]. NF play a vital role in maintaining the population of nematodes through natural methods, such as parasitism, trapping, and poisoning [16][17][43,44]. Basidiomycota trapping fungi use spores and adhesive knobs to capture nematodes [18][19][20][45,46,47]. The Orbiliaceae family comprises over 80% of nematode-trapping fungi within the Ascomycota Phylum, where constricting rings, adhesive networks, adhesive branches, adhesive knobs, and non-constricting rings are all used to ensnare nematodes (Figure 1) [21][22][48,49]. The study of nematode-trapping fungi in Zygomycota has faced obstacles due to inadequate isolation and culture techniques, despite the growing interest in it [23][50].
2.1. Adhesive Branches
Adhesive branches, also known as adhesive columns, have a simpler morphological structure than other capture organs (Figure 1). These vertical branches consist of one to three cells that merge via anastomosis, forming adhesive hoops or networks with two-dimensional structures resembling crochet or lines. The nematode is easily captured upon contact with the branch due to its complete coverage by a delicate adhesive layer. Due to the proximity of adhesive branches, nematodes frequently become stuck to more sticky hyphae upon contact and struggle to detach themselves. The species commonly found in temperate soils with developed adhesive branches is Dactylella cionopaga [24][51]. These branches serve as typical trapping mechanisms for Monacrosporium cionopagum and M. gephyrophagum [25][52]. For example, M. cionopagum produces adhesive branches that trap and immobilize the sugar beet cyst nematode Heterodera schachtii [26][53]. Likewise, Gamsylella gephyropaga produces adhesive branches to trap nematodes [27][28][54,55].
2.2. Adhesive Hyphal Network
The adhesive network, widely distributed in fungi, is comprised of an upright lateral branch emerging from a vegetative hypha (Figure 1), extending approximately 20–25 μm from the parent hypha [29][56], and is characterized by a longer lifespan in comparison to typical hyphae [13][40]. These adhesive nets are constructed using intricate three-dimensional networks. A. oligospora, with a global distribution, is the most frequently observed species in this specific trapping structure [30][57]. Adhesive nets form from vegetative hyphae by curving a solitary lateral branch and can combine with parental hyphae. Adhesive nets are regarded as an evolutionary progression from adhesive branches. More lateral hyphae are generated from the parental hyphae, or a loop is formed to generate additional loops once a complex of interconnected loops that extend away from the potential hyphae in all logical directions is established. Nematodes are attracted to the network’s surface, which is coated with a thin layer of adhesives [31][58].
2.3. Adhesive Knobs
Adhesive knobs are specialized cells with a small layer of adhesive covering them (Figure 1). When a nematode becomes ensnared, the contact area between it and the spherical knob is limited, allowing it to resist and free itself. However, upon coming into contact with a flattened, sticky pad, the fungus takes control and traps the nematode. This significantly increases the adhering surface, resulting in a secure binding of the captured nematode, followed by fungal penetration that involves both enzymatic and physical mechanisms. For example, the fungus synthesis of collagenase aids in penetrating the nematode’s cuticle, while the dense sticky pad provides strength and rigidity, allowing the piercing hyphae to move toward the cuticle [24][51]. Once a spherical infection bulb has formed, assimilative hyphae emerge to consume the internal contents of the nematode [32][33][31,59]. Dactylellina arcuata, D. asthenopaga, D. leptosphora, D. copepodii, and D. ellipsospora use adhesive knobs to capture nematodes [34][35][36][60,61,62].
2.4. Constricting Rings
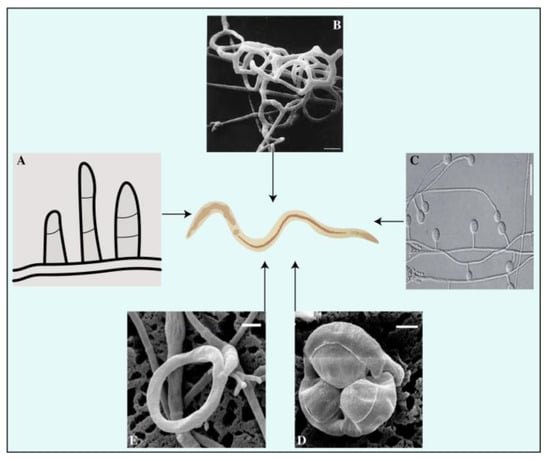
Constricting rings are hyphal branches with a circular arrangement typically composed of three cells (Figure 2). These structures are highly sophisticated and actively capture prey. A nematode entering the cavity triggers the rapid expansion of the three surrounding cells, resulting in a threefold increase in their volume. This process effectively seals the orifice and confines the nematode inside the cavity. Subsequently, the hyphae penetrate and assimilate the nematode [37][63]. Twelve species of hyphomycetes have been discovered to form constricting rings of varying internal diameters ranging from 20 to 40 μm [24][51]. Constricting rings are distinct from other mechanisms because they encircle nematodes upon contact with the inner edge of the trap, resulting in closure. The nematode is strangled by the expansion of an internally located cell wall of a constricting ring, causing inward swelling through the outer cell wall of these rings. Rapid water intake causes an increase in the volume of the cells forming the ring [38][64]. The traps of D. brochopaga mutants are considerably larger, almost eight times more than conventional traps. It has been observed that the cells in these traps release fluid droplets, reducing their volume. The humidity of the surrounding environment can be adjusted to facilitate the entry or exit of atmospheric water into or out of these cells. Additionally, a correlation has been established between the ambient humidity level and the frequency of ring closures [39][65]. The source of the water supply has been investigated, and evidence suggests that it mainly originates primarily from stalk cells or mycelium [40][66]. Furthermore, once the stalk cell closed, there was no noticeable movement of internal components, suggesting that water from the surrounding environment may have been retained [41][67]. This is supported by the observation that rings can continue to spread even after being detached from the original stalk on which they first appeared [39][42][65,68]. This idea is plausible as live nematodes are typically surrounded by a thin layer of water, which could serve as a sufficient source of moisture for the process of ring closure. Additionally, the ring closure process in D. brochopaga can be chemically induced, in addition to physical methods like touch, elevated temperature, or electrical stimulation [43][69]. When exposed to solutions containing methanol, ethanol, propanol, or butanol, or to chlorobutanol vapor, this fungus’s traps expanded in 10 to 15 s. In contrast, it is important to note that benzene, ether, and chloroform did not have any detectable effect, suggesting that unidentified variables drive this significant phenomenon [44][70].
2.5. Non-Constricting Rings
Non-constricting rings are a type of three-celled rings that grow on a short supportive stalk originating from prostrate septate hyphae (Figure 2D). The nematodes display passive behavioral responses during the predation process. As noted, the attachment point between the supporting stalk and the ring was weakened. As the nematode attempted to free itself, the ring often detached, indicating that the fungus may have facilitated its escape by allowing it to carry the non-constricting ring tightly wrapped around its body. This seems to be a favored method for achieving widespread dissemination in the soil [45][71]. For example, fungi, such as Dactylaria candida and D. lysipaga, which produce non-constricting rings, often create adhesive knobs [34][46][60,72] and capture nematodes using non-constricting rings [35][61]. A similar pattern was observed in Dactylellina daliensis via non-constricting rings [47][73].
3. Egg- and Female-Parasitic Fungi
Research on egg- and female-parasitic fungi has been in progress since the 1990s. These fungi employ appressoria (Purpureocillium spp. and Pochonia spp.), zoospores (Nematophthora gynophila), lateral mycelial branches, and penetration pegs to parasitize eggs, females, and other growth stages of the PPNs [52][78]. An in vitro assessment was conducted to evaluate the parasitism of 10 isolates of P. chlamydosporia on Globodera pallida eggs; the levels of observed pathogenicity ranged from 34% to 49%. The event of impulsive hatching occurs when P. chlamydosporia isolates aggressively parasitize immature eggs as opposed to those containing second-stage juveniles [53][79]. Additionally, the use of wild-type Beauveria bassiana 08F04 and transformant G10 resulted in a substantial decrease in the cereal cyst nematode (female) population in the roots [54][80]. In a greenhouse study, it was found that the presence of the arbuscular mycorrhizal fungus (AMF) Glomus etunicatum reduced the population of H. glycines female nematodes by 28.21% in root systems compared to untreated roots. This finding suggests that G. etunicatum may play a role in promoting the ability of host plants to tolerate the presence of the soybean cyst nematode (SCN) [55][81]. Figure 3 displays fungal species that parasitize the egg and female.
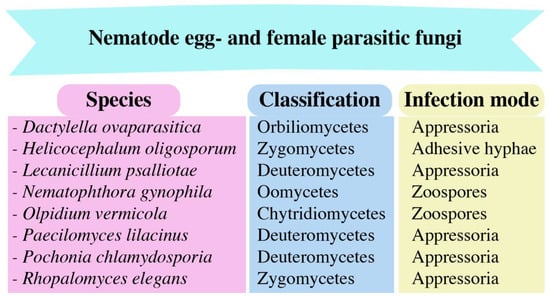
Figure 3.
The nematode egg- and female-parasitic fungi and their infection modes.
4. Endoparasitic Fungi
Endoparasitic fungi are a category of nematophagous fungi which infect nematodes by producing spores. These spores can either be internalized by the nematodes through ingestion, leading to infection, or attach to the nematode epidermis, initiating the infection [8][56][57][14,82,83]. Endoparasites utilize spores, such as conidia and zoospores, for infection, which may attach to the nematode cuticle or be ingested [58][84]. It has been found that endoparasitic fungi can reduce the number of root-knot nematodes that create galls on tomatoes and alfalfa in greenhouse experiments [8][14]. These fungi exhibit varying degrees of diversity, with studies indicating differences in their production of conidia per infected nematode. D. coniospora fungi produce a significant amount of conidia, with up to 10,000 per hyphal material, while H. rhossoliensis yields 100–1000 conidia per infected nematode. Conidia germinate immediately, and assimilative hyphae infiltrate and absorb the entire contents of the nematode body, enabling the fungus to penetrate the host’s outer layer [59][85]. D. coniospora is an aggressive endoparasitic fungus that targets nematodes. The endoparasitic fungus Drechmeria coniospora YMF1.01759 strain exhibited excellent nematode-infecting ability. The study revealed that it hindered nematodes from hatching their eggs, infected them with spores, and produced active metabolites that killed them [60][86].
5. Toxin Production
Some nematophagous fungi produce toxins that kill nematodes and impact plant defense and resistance mechanisms against parasitic nematodes [61][62][63][87,88,89]. Toxin-producing fungi originate from various orders and families. The fungus assaults nematodes via the secretion of inhibitory metabolites without physically interacting and immobilized them [52][64][78,90]. After immobilization, the hyphae penetrate the nematode cuticle. Culture filtrates of these fungi contain strong enzymatic (proteolytic and chitinolytic) activities, low-molecular-weight metabolites, and specific non-volatile oil components that cause larval death or inhibit egg hatching [65][91]. The metabolites secreted by the fungi alter the composition of nematode eggs and prevent embryonic development, rendering them unable to hatch due to their varying shapes and sizes. Similarly, fungi produce toxic chemicals, other than enzymes, that immobilize nematodes and later consume them [66][92]. Basidiomycetes are the predominant fungi that produce toxins. Recent research on Basidiomycetous fungi (Coprinus comatus and Stropharia rugosoannulata) has revealed that the action mechanisms of these toxins against nematodes are varied and multifaceted [67][93]. Among Basidiomycetes, numerous Pleurotus species produce toxins with nematotoxic activity [66][68][92,94]. For example, P. ostreatus produces trans-2-decenoic acid, a compound obtained from linoleic acid that is detrimental to nematodes, insects, and other fungi [69][95]. Basidiomycetes are not the only fungi that generate these kinds of toxins; some fungi also produce toxins that are harmful to nematodes, but these are not nematophagous [11][38]. These compounds exhibit diverse chemical properties, including simple fatty acids or other organic acids such as lactones, pyrones, anthraquinones, benzoquinones, alkaloids, furans, peptaibiotics, and cyclodepsipeptides.