Within the ruminant intestines, a diverse array of microbiota predominates, predominantly comprised of bacteria with a specialized role in breaking down intricate nutrients like cellulose and hemicellulose. This breakdown results in the conversion of these complex compounds into simpler constituents, such as glucose, which ultimately facilitates the assimilation of nutrients. The composition of these microbial communities within the rumen is contingent on a multitude of factors. These include breed, age, external environment, diet, and nutritional factors. Moreover, other variables, such as the process of weaning, energy requirements, and the presence of potentially toxic metabolic byproducts, can also exert an impact on the microbial composition and abundance within the rumen. External factors, including heat stress, psychological stress, environmental conditions, and dietary patterns, can likewise pose threats to the stability of rumen microflora.
- Microbiome
- Homeostasis
- Rumens
1. Introductions
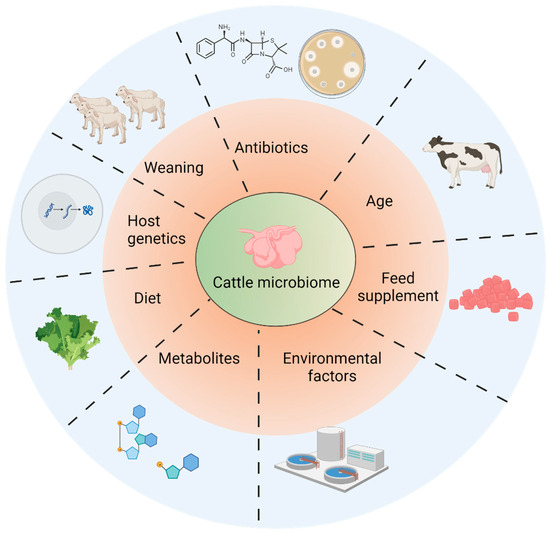
2. Factors Affecting Microbiome Establishment in Rumens
2.1. Age-Dependent Changes in Microbial Population
2.2. Stress-Related Changes in the Composition of the Microbiota
Heat stress is a multifaceted phenomenon influenced by a combination of factors, encompassing ambient temperature, relative humidity, solar radiation, and air movement. It gives rise to a range of noticeable symptoms, including elevated body temperature, accelerated breathing rate, decreased feed consumption, and heightened water intake. The ramifications of heat stress extend to reduced animal performance, and it also exerts an impact on the microflora present in their systems. When faced with heat stress, ruminants exhibit a response by curtailing their intake of dry matter. This serves the purpose of diminishing metabolic heat production, thereby facilitating the maintenance of a stable body temperature [78][21]. Among ruminants, dairy cattle stand out as particularly vulnerable to the effects of heat stress. This vulnerability is accentuated by the additional demands posed by milk production. As the specter of global warming looms larger, the prevalence of heat stress is increasingly becoming a pressing concern, not only for dairy cattle but for animals across the spectrum, including ruminants (as depicted in Figure 3). This underscores the far-reaching consequences of heat stress on animal well-being and productivity in the context of changing climatic conditions.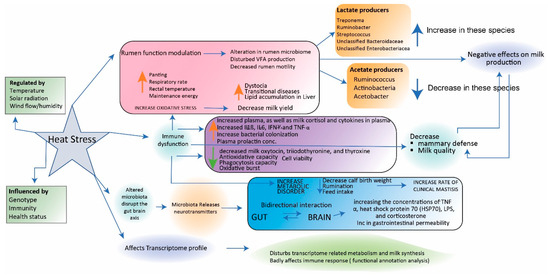
References
- Lo, B.C.; Chen, G.Y.; Núñez, G.; Caruso, R. Gut microbiota and systemic immunity in health and disease. Int. Immunol. 2021, 33, 197–209.
- Clemmons, B.A.; Voy, B.H.; Myer, P.R. Altering the gut microbiome of cattle: Considerations of host-microbiome interactions for persistent microbiome manipulation. Microb. Ecol. 2019, 77, 523–536.
- Sanjorjo, R.A.; Tseten, T.; Kang, M.-K.; Kwon, M.; Kim, S.-W. In Pursuit of Understanding the Rumen Microbiome. Fermentation 2023, 9, 114.
- Perez-Lopez, A.; Behnsen, J.; Nuccio, S.P.; Raffatellu, M. Mucosal immunity to pathogenic intestinal bacteria. Nat. Rev. Immunol. 2016, 16, 135–148.
- Li, F.; Li, C.; Chen, Y.; Liu, J.; Zhang, C.; Irving, B.; Guan, L.L. Host genetics influence the rumen microbiota and heritable rumen microbial features associate with feed efficiency in cattle. Microbiome 2019, 7, 92.
- Matthews, C.; Crispie, F.; Lewis, E.; Reid, M.; O’Toole, P.W.; Cotter, P.D. The rumen microbiome: A crucial consideration when optimising milk and meat production and nitrogen utilisation efficiency. Gut Microbes 2019, 10, 115–132.
- Rey, M.; Enjalbert, F.; Combes, S.; Cauquil, L.; Bouchez, O.; Monteils, V. Establishment of ruminal bacterial community in dairy calves from birth to weaning is sequential. J. Appl. Microbiol. 2014, 116, 245–257.
- Li, R.W.; Connor, E.E.; Li, C.; Baldwin, V.I.R.L.; Sparks, M.E. Characterization of the rumen microbiota of pre-ruminant calves using metagenomic tools. Environ. Microbiol. 2012, 14, 129–139.
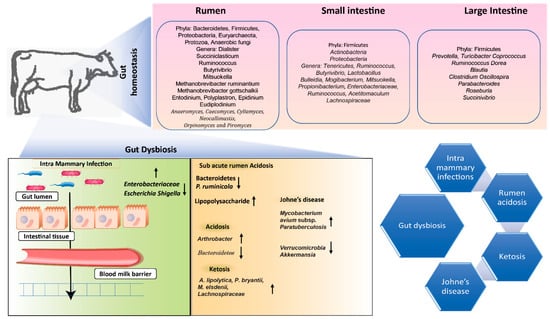
- Khalil, A.; Batool, A.; Arif, S. Healthy cattle microbiome and dysbiosis in diseased phenotypes. Ruminants 2022, 2, 134–156.
- De Barbieri, I.; Hegarty, R.S.; Silveira, C.; Gulino, L.M.; Oddy, V.H.; Gilbert, R.A.; Klieve, A.V.; Ouwerkerk, D. Programming rumen bacterial communities in newborn Merino lambs. Small Rumin Res. 2015, 129, 48–59.
- Meale, S.J.; Popova, M.; Saro, C.; Martin, C.; Bernard, A.; Lagree, M.; Yáñez-Ruiz, D.R.; Boudra, H.; Duval, S.; Morgavi, D.P. Early life dietary intervention in dairy calves results in a long-term reduction in methane emissions. Sci. Rep. 2021, 11, 3003.
- Abecia, L.; Waddams, K.E.; Martínez-Fernandez, G.; Martín-García, A.I.; Ramos-Morales, E.; Newbold, C.J.; Yáñez-Ruiz, D.R. An antimethanogenic nutritional intervention in early life of ruminants modifies ruminal colonization by Archaea. Archaea 2014, 2014, 841463.
- O’Callaghan, T.F.; Ross, R.P.; Stanton, C.; Clarke, G. The gut microbiome as a virtual endocrine organ with implications for farm and domestic animal endocrinology. Domest. Anim. Endocrinol. 2016, 56, S44–S55.
- Liu, S.; Yu, Z.; Zhong, H.; Zheng, N.; Huws, S.; Wang, J.; Zhao, S. Functional gene-guided enrichment plus in situ microsphere cultivation enables isolation of new crucial ureolytic bacteria from the rumen of cattle. Microbiome 2023, 11, 76.
- McAllister, T.A.; Cheng, K.J.; Okine, E.K.; Mathison, G.W. Dietary, environmental and microbiological aspects of methane production in ruminants. Can. J. Anim. Sci. 1996, 76, 231–243.
- Mekuriaw, S.; Tsunekawa, A.; Ichinohe, T.; Tegegne, F.; Haregeweyn, N.; Kobayashi, N.; Tassew, A.; Mekuriaw, Y.; Walie, M.; Tsubo, M.; et al. Effect of Feeding Improved Grass Hays and Eragrostis tef Straw Silage on Milk Yield, Nitrogen Utilization, and Methane Emission of Lactating Fogera Dairy Cows in Ethiopia. Animals 2020, 10, 1021.
- Hempel, S.; Menz, C.; Pinto, S.; Galán, E.; Janke, D.; Estellés, F.; Müschner-Siemens, T.; Wang, X.; Heinicke, J.; Zhang, G.; et al. Heat stress risk in European dairy cattle husbandry under different climate change scenarios–uncertainties and potential impacts. Earth Syst. Dyn. 2019, 10, 859–884.
- Khafipour, E.; Li, S.; Tun, H.M.; Derakhshani, H.; Moossavi, S.; Plaizier, J.C. Effects of grain feeding on microbiota in the digestive tract of cattle. Anim. Front. 2016, 6, 13–19.
- Deusch, S.; Camarinha-Silva, A.; Conrad, J.; Beifuss, U.; Rodehutscord, M.; Seifert, J. A structural and functional elucidation of the rumen microbiome influenced by various diets and microenvironments. Front. Microbiol. 2017, 8, 1605.
- Zhou, M.; Ghoshal, B.; Stothard, P. Distinctive roles between rumen epimural and content bacterial communities on beef cattle feed efficiency: A combined analysis. Curr. Res. Microb. Sci. 2021, 2, 100085.
- Arshad, M.A.; Hassan, F.U.; Rehman, M.S.; Huws, S.A.; Cheng, Y.; Din, A.U. Gut microbiome colonization and development in neonatal ruminants: Strategies, prospects, and opportunities. Anim. Nutr. 2021, 7, 883–895.
- Plaizier, J.C.; Li, S.; Danscher, A.M.; Derakshani, H.; Andersen, P.H.; Khafipour, E. Changes in microbiota in rumen digesta and feces due to a grain-based subacute ruminal acidosis (SARA) challenge. Microb. Ecol. 2017, 74, 485–495.
- Thornton, P.; Nelson, G.; Mayberry, D.; Herrero, M. Impacts of heat stress on global cattle production during the 21st century: A modelling study. Lancet Planet. Health 2022, 6, e192–e201.