The underlying philosophy/hypothesis of organic farming—“healthy soils generate healthy crops which lead to healthy livestock and humans”—was first formulated by Sir Albert Howard and other organic farming pioneers. Organic crop production practices could increase the nutritional quality of food crops, the evidence now available suggests that there are nutritionally relevant composition differences between organic and conventional crops and that, overall, organic plant-foods contain higher concentrations of nutritionally desirable (phenolics, other antioxidants and/or mineral micronutrients) and lower concentrations of nutritionally undesirable chemicals (pesticide residues, cadmium, and/or Fusarium mycotoxins). However, agricultural intensification practices have reduced food quality and safety, the available evidence indicates that many “innovations” (monoculture/shorter rotations, mineral N and P fertilizers, pesticides, and short-straw cereal varieties) introduced as part of the agricultural intensification (or “green revolution”) have had negative effects on both crop health and the nutritional quality and/or safety of crops.
- organic farming
- conventional farming
- food composition
- human health
- food security
- phenolics
- antioxidants
- mineral micronutrients
- cadmium
- pesticides
1. Crop Rotation Design
2. Fertilization Regimes
2.1. Replacing Mineral N-Fertilizer with Organic Fertilizer Inputs
| ANOVA Results (p-Values) | ||||||||
|---|---|---|---|---|---|---|---|---|
| Crop Protection (CP) | Fertilization (F) | Man Effects | Inter-Action | |||||
| Crop | Parameter Assessed | CON | ORG | Mineral NPK | Cattle Manure | CP | F | CP×F |
| Potato | Marketable tuber yield (t/ha) | 9.2 | 7.7 | 9.3 | 7.6 | <0.001 | <0.001 | 0.003 |
| total phenolics (µg/g FW) | 285 | 293 | 277 | 301 | NS | 0.016 | NS | |
| vitamin C (µg/g FW) | 95 | 97 | 91 | 101 | NS | <0.001 | NS | |
| total carotenoids (µg/g FW) | 0.7 | 0.8 | 0.7 | 0.7 | 0.060 | NS | 0.018 | |
| folate (µg/g FW) | 0.17 | 0.18 | 0.17 | 0.18 | NS | <0.001 | 0.006 | |
| glycoalkaloids (mg/kg FW) | 42 | 40 | 45 | 37 | NS | <0.001 | NS | |
| Cabbage | Yield (t/ha FW) | 54 | 61 | 71 | 45 | <0.001 | <0.001 | 0.007 |
| total phenolics 1 (µg/g FW) | 11.0 | 9.4 | 8.8 | 11.6 | 0.093 | 0.001 | NS | |
| vitamin C (µg/g FW) | 224 | 223 | 211 | 236 | NS | 0.006 | NS | |
| total carotenoids (µg/g FW) | 3.5 | 3.2 | 3.0 | 3.7 | NS | 0.008 | 0.051 | |
| folate (µg/g FW) | 0.4 | 0.3 | 0.3 | 0.4 | <0.001 | 0.022 | NS | |
| total glucosinolates (g/g FW) | 1.37 | 1.23 | 1.13 | 1.48 | 0.037 | <0.001 | NS | |
| total volatiles 2 (mg/g FW) | 33 | 31 | 28 | 35 | NS | <0.001 | 0.032 | |
| Lettuce | Yield (t/ha FW) | 38 | 36 | 39 | 34 | NS | <0.001 | NS |
| total phenolics (µg/g FW) | 104 | 108 | 102 | 112 | NS | 0.036 | NS | |
| vitamin C (µg/g FW) | 6.8 | 6.7 | 6.8 | 6.7 | NS | NS | NS | |
| total carotenoids (µg/g FW) | 4.4 | 5.0 | 4.6 | 4.8 | 0.085 | NS | NS | |
| folate (µg/g FW) | 0.3 | 0.3 | 0.3 | 0.3 | 0.064 | 0.042 | NS | |
| Onion | Marketable yield (t/ha FW) | 22 | 20 | 21 | 21 | NS | NS | NS |
| total phenolics 3 (µg/g FW) | 725 | 721 | 740 | 706 | NS | NS | NS | |
| vitamin C (µg/g FW) | 100 | 93 | 86 | 107 | NS | NS | NS | |
| total carotenoids (µg/g FW) | 0.6 | 0.7 | 0.7 | 0.6 | NS | NS | 0.083 | |
| folate (µg/g FW) | 0.4 | 0.4 | 0.4 | 0.4 | NS | NS | NS | |
2.2. Replacing Superphosphate with Organic Fertilizer Inputs
2.3. Rhizobium and Mycorrhizal Fungal Inocula
3. Crop Protection
3.1. Soil/Substrate Suppressiveness against Pests and Diseases
3.2. Environmental Enrichment Practices
3.3. Elicitors of Crop Resistance
-
Chitin (a long-chain polymer of N-acetylglucosamine, which is a primary component of fungal cell walls and the exoskeleton of insects and crustaceans) is mainly produced from the shells of shrimps and other crustaceans;
-
Chitosan (a linear polysaccharide composed of randomly distributed β-(1→4)-linked D-glucosamine and N-acetyl-D-glucosamine) is produced by treating the chitin shells of shrimp and other crustaceans with an alkaline substance, such as sodium hydroxide;
-
Giant knotweed (Reynoutria sachalinensis Schmidt ex Maxim.) extracts (tradename: Milsana®).
3.4. Biological Control Products
3.5. Barrier, Mating Disruption, and Mass-Trapping Methods for Pest Control
3.6. Thermal Crop Protection Methods and Pesticide Products Used in Organic Farming
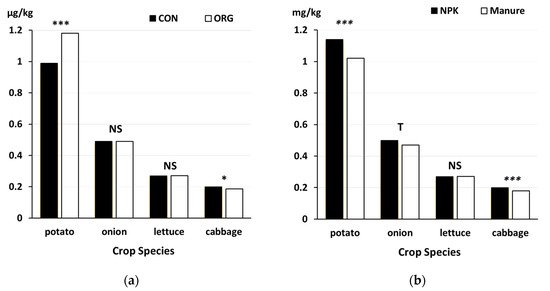
3.7. Synthetic Chemical Pesticides That Are Prohibited in Organic Farming
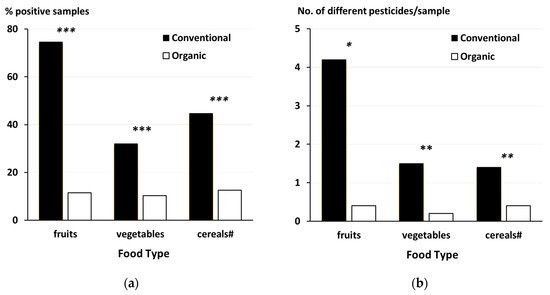
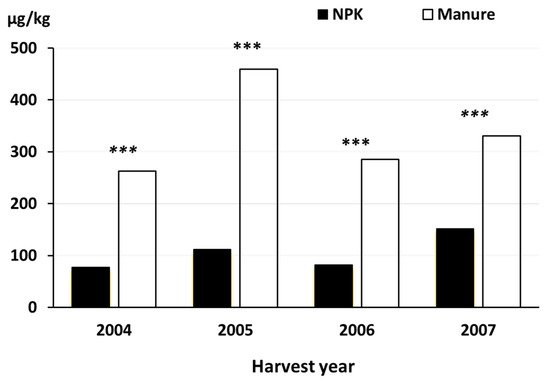
-
“Impairment of the innate physiological defenses by interruption of the shikimic acid pathway”;
-
“Interference with rhizosphere microbial ecology (in particular, GBHs have the potential to enhance the population and/or virulence of some phytopathogenic microbial species in the crop rhizosphere)”;
-
“As yet incompletely elucidated reduction in the uptake and utilization of nutrient metals by crops”.
4. Crop Breeding and Selection
4.1. Wheat
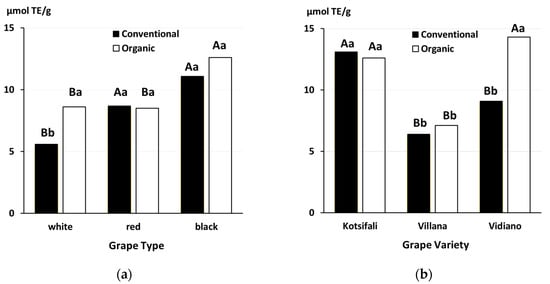
4.2. Grapes
References
- Hansen, A.L. The Organic Farming Manual: A Comprehensive Guide to Starting and Running a Certified Organic Farm; Storey Publishing: North Adams, MA, USA, 2010.
- Blake, F. Organic Farming and Growing; The Crowood Press: Marlborough, UK, 2005.
- Nandwani, D. Organic Farming for Sustainable Agriculture; Springer International Publishing: Basel, Switzerland, 2016.
- Haghighi, R.S.; Critchley, N.; Leifert, C.; Eyre, M.; Cooper, J. Individual and interactive effects of crop type and management on weed and seed bank composition in an organic rotation. Int. J. Plant Prod. 2013, 7, 243–268.
- Eyre, M.D.; Critchley, C.N.R.; Leifert, C.; Wilcockson, S.J. Crop sequence, crop protection and fertility management effects on weed cover in an organic/conventional farm management trial. Eur. J. Agron. 2011, 59, 4715–4724.
- Hill, O. How grazing livestock can beat blackgrass. Farmers Wkly. 2015. Available online: www.fwi.co.uk/arable/grazing-livestock-can-beat-blackgrass (accessed on 24 February 2021).
- Råberg, T.; Carlsson, G.; Jensen, E.S. Nitrogen balance in a stockless organic cropping system with different strategies for internal N cycling via residual biomass. Nutr. Cycl. 2018, 112, 165–178.
- Barbieri, P.; Pellerin, S.; Nesme, T. Comparing crop rotations between organic and conventional farming. Sci. Rep. 2017, 7, 13761.
- Sandra Wayman, S.; Kissing Kucek, L.; Mirsky, S.B.; Ackroyd, V.; Cordeau, S.; Ryan, M.R. Organic and conventional farmers differ in their perspectives on cover crop use and breeding. Renew. Agric. Food Syst. 2016, 32, 376–385.
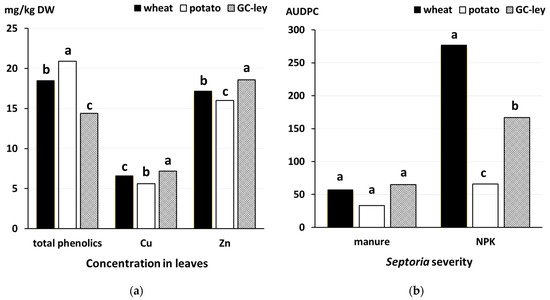
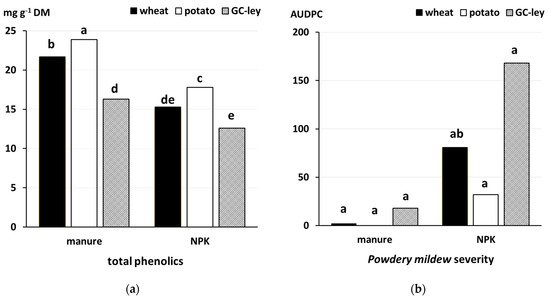
- Rempelos, L.; Almuayrifi, A.M.; Baranski, M.; Tetard-Jones, C.; Eyre, M.; Shotton, P.; Cakmak, I.; Ozturk, L.; Cooper, J.; Volakakis, N.; et al. Effects of agronomic management and climate on leaf phenolic profiles, disease severity and grain yield in organic and conventional wheat production systems. J. Agric. Food Chem. 2018, 66, 10369–10379.
- Leifert, C.; Rembiałkowska, E.; Nielson, J.H.; Cooper, J.M.; Butler, G.; Lueck, L. Effects of organic and ‘low input’ production methods on food quality and safety. In Improving Sustainability in Organic and Low Input Food Production Systems; Niggli, U., Leifert, C., Alföldi, T., Lueck, L., Willer, H., Eds.; FiBL: Frick, Switzerland, 2007; pp. 15–96.
- Palmer, M.W.; Cooper, J.; Tétard-Jones, C.; Średnicka-Tober, D.; Baranski, M.; Eyre, M.; Shotton, P.; Volakakis, N.; Cakmak, I.; Ozturk, L.; et al. The influence of organic and conventional fertilisation and crop protection practices, preceding crop, harvest year and weather conditions on yield and quality of potato (Solanum tuberosum) in a long-term management trial. Eur. J. Agron. 2013, 49, 83–92.
- Keiser, A.; Haeberli, M.; Stamp, P. Quality deficiencies on potato (Solanum tuberosum L.) tubers caused by Rhizoctonia solani, wireworms (Agriotes ssp.) and slugs (Deroceras reticulatum, Arion hortensis) in different farming systems. Field Crop. Res. 2012, 128, 147–155.
- Benaragama, D.; Leeson, J.L.; Shirtliffe, S.J. Understanding the long term weed community dynamics in organic and conventional crop rotations using the principal response curve method. Weed Sci. 2018, 67, 195–204.
- Robson, M.C.; Fowler, S.M.; Lampkin, N.H.; Leifert, C.; Leitch, M.; Robinson, D.; Watson, C.A.; Litterick, A.M. The agronomic and economic potential of break crops for ley/arable rotations in temperate organic agriculture. Adv. Agron. 2002, 77, 370–427.
- Paulsen, H.M.; Weißmann, F. Relevance of mycotoxins to product quality and animal health in organic farming. In Proceedings of the 14th IFOAM Organic World Congress, Victoria, Canada, 21–24 August 2002; Available online: https://orgprints.org/2023/1/2023-paulsen-weissmann-2002-14th-ifoam-ca-p212.pdf (accessed on 24 February 2021).
- Bernhoft, A.; Clasen, P.-E.; Kristoffersen, A.B.; Torp, M. Less Fusarium infestation and mycotoxin contamination in organic than in conventional cereals. Food Addit. Contam. 2010, 27, 842–852.
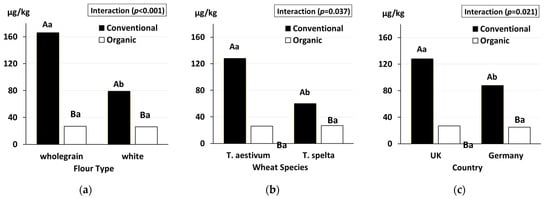
- Wang, J.; Hasanalieva, G.; Wood, L.; Markellou, E.; Iversen, P.O.; Bernhoft, A.; Seal, C.; Baranski, M.; Vigar, V.; Ernst, L.; et al. Effect of wheat species (Triticum aestivum vs. T. spelta), farming system (organic vs. conventional) and flour type (wholegrain vs. white) on composition of wheat flour; results of a retail survey in the UK and Germany—1. mycotoxin content. Food Chem. 2020, 327, 127011.
- Wang, J. Effect of Organic and Conventional Agronomic Practices and Variety Choice on Nutritional Quality, the Contents of Undesirable Compounds and Yield of Cereals. Ph.D. Thesis, Newcastle University, Newcastle Upon Tyne, UK, 2019.
- Thavarajah, D.; Siva, N.; Johnson, N.; McGee, R.; Thavarajah, P. Effect of cover crops on the yield and nutrient concentration of organic kale (Brassica oleracea L. var. acephala). Sci. Rep. 2019, 9, 10374.
- Tilman, D.; Cassman, K.G.; Matson, P.A.; Naylor, R.; Polasky, S. Agricultural sustainability and intensive production practices. Nature 2002, 418, 671–677.
- Cordell, D.; Drangert, J.-O.; White, S. The story of phosphorus: Global food security and food for thought. Glob. Environ. Chang. 2009, 19, 292–305.
- Chowdhury, R.B.; Moore, G.A.; Weatherley, A.J.; Arora, M. Key sustainability challenges for the global phosphorus resource, their implications for global food security, and options for mitigation. J. Clean. Prod. 2017, 140, 945–963.
- Cordell, D.; Jackson, M.; White, S. Phosphorus flows through the Australian food system: Identifying intervention points as a roadmap to phosphorus security. Environ. Sci. Policy 2013, 29, 87–102.
- Farm Carbon Toolkit. Fertiliser Production. 2021. Available online: www.farmcarbontoolkit.org.uk/toolkit/fertiliser-production (accessed on 25 February 2021).
- European Union. Commission Implementation Regulation (EU) 2016/673 of 29 April 2016 Amending Regulation (EC) No 889/2008 Laying Down Detailed Rules for the Implementation of Council Regulation (EC) No 834/2007 on Organic Production and Labelling of Organic Products with Regard to Organic Production, Labelling and Control. 2016. Available online: https://eur-lex.europa.eu/eli/reg_impl/2016/673/oj (accessed on 23 February 2021).
- Niggli, U. History and concepts of food quality and safety in organic food production and processing. In Handbook of Organic Food Safety and Quality; Cooper, J., Niggli, U., Leifert, C., Eds.; Woodhead Publishing Ltd.: Cambridge, UK, 2007; pp. 9–24.
- Alburquerque, J.A.; de la Fuente, C.; Campoy, M.; Carrasco, L.; Nájera, I.; Baixauli, C.; Caravaca, F.; Roldán, A.; Cegarra, J.; Bernal, M.P. Agricultural use of digestate for horticultural crop production and improvement of soil properties. Eur. J. Agron. 2012, 119–128.
- Gerlach, F.; Grieb, B.; Zerger, U. Sustainable Biogas Production; A Handbook for Organic Farmers; FiBL Projecte GmbH: Frankfurt, Germany, 2013; Available online: https://aa.ecn.cz/img_upload/8d8825f1d3b154e160e6e5c97cf9b8b3/sustaingas_handbook_en.pdf (accessed on 4 December 2021).
- Magistrali, A.; Vavera, R.; Janovska, D.; Rempelos, L.; Cakmak, I.; Leifert, C.; Grausgruber, H.; Butler, G.; Wilkinson, A.; Bilsborrow, P. Evaluating the effect of agronomic management practices on the performance of differing spelt (Triticum spelta) cultivars in contrasting environments. Field Crop. Res. 2020, 255, 107869.
- Rempelos, L.; Almuayrifi, M.S.B.; Barański, M.; Tetard-Jones, C.; Barkla, B.; Cakmak, I.; Ozturk, L.; Cooper, J.; Volakakis, N.; Hall, G.; et al. The effect of agronomic factors on crop health and performance of winter wheat varieties bred for the conventional and the low input farming sector. Field Crop. Res. 2020, 254, 107822. Available online: https://authors.elsevier.com/sd/article/S0378429019315990 (accessed on 4 November 2021).
- Margaritopoulou, T.; Toufexi, E.; Kizis, D.; Balayiannis, G.; Anagnostopoulos, C.; Theocharis, A.; Rempelos, L.; Troyanos, Y.; Leifert, C.; Markellou, E. Reynoutria sachalinensis extract elicits SA-dependent defence responses in courgette genotypes against powdery mildew caused by Podosphaera xanthii. Sci. Rep. 2020, 10, 3354.
- Sander, J.F.; Heitefuss, R. Susceptibility to Erysiphe graminis f. sp. tritici and phenolic acid content of wheat as influenced by different levels of nitrogen fertilization. J. Phytopathol. 1988, 146, 495–507.
- Leser, C.; Treutter, D. Effects of nitrogen supply on growth, contents of phenolic compounds and pathogen (scab) resistance of apple trees. Physiol. Plant. 2004, 123, 49–56.
- Sun, Y.; Guo, J.; Li, Y.; Luo, G.; Li, L.; Yuan, H.; Mur, L.A.J.; Guo, S. Negative effects of the simulated nitrogen deposition plant phenolic metabolism: A meta-analysis. Sci. Total Environ. 2020, 719, 137442.
- Lin, D.; Xiao, M.; Zhao, J.; Li, Z.; Xing, B.; Li, X.; Kong, M.; Li, L.; Zhang, Q.; Liu, Y.; et al. An Overview of Plant Phenolic Compounds and Their Importance in Human Nutrition and Management of Type 2 Diabetes. Molecules 2016, 21, 1374.
- Brandt, K.; Leifert, C.; Sanderson, R.; Seal, C.J. Agroecosystem management and nutritional quality of plant foods: The case of organic fruits and vegetables. CRC Crit. Rev. Plant Sci. 2011, 30, 177–197.
- Baranski, M.; Średnicka-Tober, D.; Volakakis, N.; Seal, C.; Sanderson, R.; Stewart, G.B.; Benbrook, C.; Biavati, B.; Markellou, E.; Giotis, H.; et al. Higher antioxidant and lower cadmium concentrations and lower incidence of pesticide residues in organically grown crops: A systematic literature review and meta-analysis. Br. J. Nutr. 2014, 112, 794–811.
- European Commission. Final Report Summary—QUALITYLOWINPUTFOOD (Improving Quality and Safety and Reduction of Cost in the European Organic and ‘Low Input’ Supply Chains. 2009. Available online: https://cordis.europa.eu/project/id/506358/reporting (accessed on 24 February 2021).
- Wang, J.; Chatzidimitriou, E.; Wood, L.; Hasanalieva, G.; Markellou, E.; Iversen, P.O.; Seal, C.; Baranski, M.; Vigar, V.; Ernst, L.; et al. Effect of wheat species (Triticum aestivum vs T. spelta), farming system (organic vs conventional) and flour type (wholegrain vs white) on composition of wheat flour—Results of a retail survey in the UK and Germany—2. Antioxidant activity, and phenolic and mineral content. Food Chem. X 2020, 6, 100091.
- Hasanaliyeva, G. Effect of Organic Production Methods on Antioxidant Activity and Concentrations in Grapes, Grape Juice and Wine; Results from Meta-Analyses, and Farm and Retail Surveys. Ph.D. Thesis, Newcastle University, Newcastle Upon Tyne, UK, 2017.
- Murphy, K.M.; Campbell, K.G.; Lyon, S.R.; Jones, S.S. Evidence of varietal adaptation to organic farming systems. Field Crop. Res. 2007, 102, 172–177.
- Hasanaliyeva, G.; Chatzidimitrou, E.; Wang, J.; Baranski, M.; Volakakis, N.; Seal, C.; Rosa, E.A.S.; Iversen, P.O.; Vigar, V.; Barkla, B.; et al. Effects of Production Region, Production Systems and Grape Type/Variety on Nutritional Quality Parameters of Table Grapes; Results from a UK Retail Survey. Foods 2020, 16, 1874.
- Hasanaliyeva, G.; Chatzidimitrou, E.; Wang, J.; Baranski, M.; Volakakis, N.; Pakos, P.; Seal, C.; Rosa, E.A.S.; Markellou, E.; Iversen, P.O.; et al. Effect of organic and conventional Production Methods on Fruit Yield and Nutritional Quality Parameters in Table Grapes and Wine made from three traditional Cretan Grape Varieties; Results from a Farm Survey. Foods 2021, 10, 476.
- Gosling, P.; Hodge, A.; Goodlass, G.; Bending, G.D. Arbuscular mycorrhizal fungi and organic farming. Agric. Ecosyst. Environ. 2006, 113, 17–35.
- Lee, S.-W.; Lee, E.-H.; Eom, A.-H. Effects of Organic Farming on Communities of Arbuscular Mycorrhizal Fungi. Mycobiology 2008, 36, 19–23.
- Säle, V.; Aguilera, P.; Laczko, E.; Mäder, P.; Berner, A.; Zihlmann, U.; van der Heijden, M.G.A.; Oehl, F. Impact of conservation tillage and organic farming on the diversity of arbuscular mycorrhizal fungi. Soil Biol. Biochem. 2015, 84, 38–52.
- Grant, C.A.; Sheppard, S.C. Fertilizer impacts on cadmium availability in agricultural soils and Crops. Hum. Ecol. Risk Assess. 2008, 14, 210–228.
- Cooper, J.; Sanderson, R.; Cakmak, I.; Ozturk, L.; Shotton, P.; Carmichael, A.; Sadrabadi Haghighi, R.; Tetard-Jones, C.; Volakakis, N.; Eyre, M.; et al. Effect of organic and conventional crop rotation, fertilization and crop protection practices on metal contents in wheat (Triticum aestivum). J. Agric. Food Chem. 2011, 59, 4715–4724.
- Dharma-wardana, M.W.C. Fertilizer usage and cadmium in soils, crops and food. Environ. Geochem. Health 2018, 40, 2739–2759.
- Abou-Shanab, R.A.I.; Wongphatcharachai, M.; Sheaffer, C.C.; Sadowsky, M.J. Response of dry bean (Phaseolus vulgaris L.) to inoculation with indigenous and commercial Rhizobium strains under organic farming systems in Minnesota. Symbiosis 2019, 78, 125–134.
- Ardakani, M.R.; Pietsch, G.; Moghaddam, A.; Raza, A.; Friedel, J.K. Response of Root Properties to Tripartite Symbiosis between Lucerne (Medicago sativa L.), Rhizobia and Mycorrhiza. Am. J. Agric. Biol. Sci. 2009, 4, 266–277.
- Wilkinson, A. Improving the Agronomic Management and Utilization of Breadmaking Wheat. Ph.D. Thesis, Newcastle University, Newcastle upon Tyne, UK, 2020.
- Aryal, U.K.; Shah, S.K.; Xu, H.L.; Fujita, M. Growth, Nodulation and Mycorrhizal Colonization in Bean Plants Improved by Rhizobial Inoculation with Organic and Chemical Fertilization. J. Sustain. Agric. 2007, 29, 71–83.
- Faye, A.; Stewart, Z.P.; Ndung’u-Magiroi, K.; Diouf, M.; Ndoye, I.; Diop, T.; Dalpé, Y.; Prasad, P.V.V.; Lesueur, D. Testing of Commercial Inoculants to Enhance P Uptake and Grain Yield of Promiscuous Soybean in Kenya. Sustainability 2020, 12, 3803.
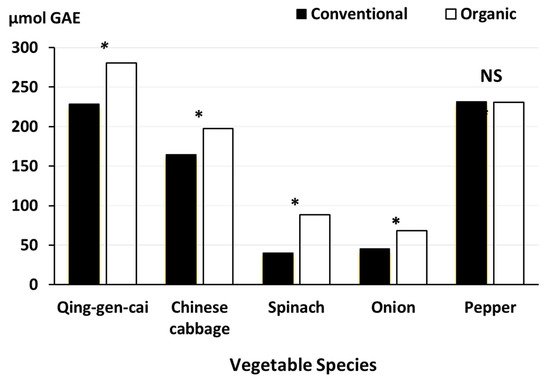
- Mukherjee, A.; Omondi, E.C.; Hepperly, P.R.; Seidel, R.; Heller, W.P. Impacts of Organic and Conventional Management on the Nutritional Level of Vegetables. Sustainability 2020, 12, 8965.
- Gianinazzi, S.; Schüepp, H.; Barea, J.M.; Haselwandter, K. Mycorrhizal Technology in Agriculture; From Genes to Bioproducts; Springer: Basel, Switzerland, 2002.
- Oliveira, R.S.; Rocha, I.; Ma, Y.; Vosátka, M.; Freitas, H. Seed coating with arbuscular mycorrhizal fungi as an ecotechnologicalapproach for sustainable agricultural production of common wheat (Triticum aestivum L.). J. Toxicol. Environ. Health Part A 2016, 79, 329–337.
- Berruti, A.; Lumini, E.; Balestrini, R.; Bianciotto, V. Arbuscular Mycorrhizal Fungi as Natural Biofertilizers: Let’s Benefit from Past Successes. Front. Microbiol. 2016, 6, 1559.
- Elliott, A.J.; Daniell, T.J.; Cameron, D.D.; Field, K.J. A commercial arbuscular mycorrhizal inoculum increases root colonization across wheat cultivars but does not increase assimilation of mycorrhiza-acquired nutrients. Plants People Planet 2020, 3, 588–599.
- Reganold, J.P.; Elliott, L.F.; Unger, Y.L. Long-term effects of organic and conventional farming on soil erosion. Nature 1987, 330, 370–372.
- Mäder, P.; Fliessbach, A.; Dubois, D.; Gunst, L.; Fried, P.; Niggli, U. Soil fertility and biodiversity in organic farming. Science 2002, 296, 1694–1697.
- Gattinger, A.; Muller, A.; Haeni, M.; Skinner, C.; Fliessbach, A.; Buchmann, N.; Mader, P.; Stolze, M.; Smith, P.; Scialabba Nel, H.; et al. Enhanced top soil carbon stocks under organic farming. Proc. Natl. Acad. Sci. USA 2012, 109, 18226–18231.
- Eyre, M.D.; Sanderson, R.A.; Shotton, P.N.; Leifert, C. Investigating the effects of crop type, fertility management and crop protection on the activity of beneficial invertebrates in an extensive farm management comparison trial. Ann. Appl. Biol. 2009, 155, 267–276.
- Hadar, Y.; Papadopoulou, K.K. Suppressive composts: Microbial ecology links between abiotic environments and healthy plants. Ann. Rev. Phytopathol. 2012, 50, 133–153.
- Yogev, A.; Laor, Y.; Katan, J.; Hadar, Y.; Cohen, R.; Medina, S.; Raviv, M. Does organic farming increase soil suppression against Fusarium wilt of melon? Org. Agric. 2011, 1, 203–216.
- Giotis, C.; Markelou, E.; Theodoropoulou, A.; Toufexi, E.; Hodson, R.; Shotton, P.; Shiel, R.; Cooper, J.; Leifert, C. Effect of soil amendments and biological control agents (BCAs) on soil-borne root diseases caused by Pyrenochaeta lycopersici and Verticillium alboatrum in organic greenhouse tomato production systems. Eur. J. Plant Pathol. 2009, 123, 387–400.
- Giotis, C.; Theodoropoulou, A.; Cooper, J.; Hodgson, R.; Shotton, P.; Shiel, R.; Eyre, M.; Wilcockson, S.; Markellou, E.; Liopa-Tsakalidis, A.; et al. Effect of variety choice, resistant rootstocks and chitin soil amendments on soil-borne diseases in soil-based, protected tomato production systems. Eur. J. Plant Pathol. 2012, 134, 605–617.
- Tamm, L.; Thürig, B.; Bruns, C.; Fuchs, J.G.; Köpke, U.; Leifert, C.; Mahlberg, N.; Nietlispach, B.; Laustela, M.; Schmidt, C.; et al. Soil type, management history, and soil amendments influence the development of soilborne (Rhizoctonia solani, Pythium ultimum) and airborne (Phytophthora infestans, Hyaloperonospora parasitica) diseases. Eur. J. Plant Pathol. 2010, 127, 465–481.
- Willson, A.; Goltz, M.; Markellou, E.; Volakakis, N.; Leifert, C. Integrating the use of resistant rootstocks/cultivars, suppressive composts and elicitors to improve yields and quality in protected organic cultivation systems. Acta Hortic. 2020, 1268, 155–164.
- Bonanomi, G.; Antignani, V.; Capodilupo, M.; Scala, F. Identifying the characteristics of organic soil amendments that suppress soilborne plant diseases. Soil Biol. Biochem. 2010, 42, 136–144.
- Litterick, A.M.; Harrier, L.; Wallace, P.; Watson, C.A.; Wood, M. The role of uncomposted materials, composts, manures, and compost extracts in reducing pest and disease incidence and severity in sustainable temperate agricultural and horticultural production—A review. CRC Crit. Rev. Plant Sci. 2004, 23, 453–479.
- Noble, R.; Coventry, E. Suppression of soil-borne plant diseases with composts: A review. Biocontrol Sci. Technol. 2005, 15, 3–20.
- Brennan, R.J.B.; Glaze-Corcoran, S.; Wick, R.; Hashemi, M. Biofumigation: An alternative strategy for the control of plant parasitic nematodes. J. Integr. Agric. 2020, 19, 1680–1690.
- Ren, H.; Endo, H.; Hayashi, T. Antioxidative and antimutagenic activities and polyphenol content of pesticide-free and organically cultivated green vegetables using water-soluble chitosan as a soil modifier and leaf surface spray. J. Sci. Food Agric. 2001, 81, 1426–1432.
- Ghasemi Pirbalouti, A.; Malekpoor, F.; Salimi, A.; Golparvar, A. Exogenous application of chitosan on biochemical and physiological characteristics, phenolic content and antioxidant activity of two species of basil (Ocimum ciliatum and Ocimum basilicum) under reduced irrigation. Sci. Hortic. 2017, 217, 114–122.
- Rahman, M.; Mukta, J.A.; Sabir, A.A.; Gupta, D.R.; Mohi-Ud-Din, M.; Hasanuzzaman, M.; Miah, M.G.; Rahman, M.; Islam, M.T. Chitosan biopolymer promotes yield and stimulates accumulation of antioxidants in strawberry fruit. PLoS ONE 2018, 13, 0203769.
- Ben-Issa, R.; Gomez, L.; Gautier, H. Companion Plants for Aphid Pest Management. Insects 2017, 8, 112.
- Eyre, M.D.; Leifert, C. Crop and field boundry influences on the activity of a wide range of beneficial invertebrate groups on a split conventional/organic farm in northern England. Bull. Entomol. Res. 2011, 101, 135–144.
- Eyre, M.D.; Labanowska-Bury, D.; White, R.; Leifert, C. Relationship between beneficial invertebrates, field margin vegetation, and thrip damage in organic leek fields in eastern England. Org. Agric. 2011, 1, 45–54.
- Eyre, M.D.; Luff, M.L.; Leifert, C. Crop, field boundary, productivity and disturbance influences on ground beetles (Coleoptera, Carabidae) in the agroecosystem. Agric. Ecosyst. Environ. 2013, 165, 60–67.
- MacLeod, A.; Wratten, S.D.; Sotherton, N.W.; Thomas, M.B. ‘Beetle banks’ as refuges for beneficial arthropods in farmland: Long-term changes in predator communities and habitat. Agric. For. Entomol. 2004, 6, 147–154.
- Bengtsson, J.; Ahnström, J.; Weibull, A.-C. The Effects of Organic Agriculture on Biodiversity and Abundance: A Meta-Analysis. J. Appl. Ecol. 2005, 42, 261–269.
- Gabriel, D.; Tscharntke, T. Insect pollinated plants benefit from organic farming. Agric. Ecosyst. Environ. 2007, 118, 43–48.
- Muneret, L.; Auriol, A.; Bonnard, O.; Richart-Cervera, S.; Thiéry, D.; Rusch, A. Organic farming expansion drives natural enemy abundance but not diversity in vineyard-dominated landscapes. Ecol. Evol. 2019, 9, 13532–13542.
- Birkhofer, K.; Arvidsson, F.; Ehlers, D.; Mader, V.L.; Bengtsson, J.; Smith, H.G. Organic farming affects the biological control of hemipteran pests and yields in spring barley independent of landscape complexity. Landsc. Ecol. 2016, 31, 567–579.
- Kasselaki, A.M.; Malathrakis, N.E.; Goumas, D.E.; Leifert, C. Effect of alternative treatments on seed borne Didymella lycopersici in tomato. J. Appl. Microbiol. 2008, 105, 36–41.
- Kasselaki, A.-M.; Goumas, D.; Tamm, L.; Fuchs, J.; Cooper, C.; Leifert, C. Effect of alternative strategies for the disinfection of tomato seed infected with bacterial canker (Clavibacter michiganensis subsp. michiganensis). NJAS-Wagen. J. Life Sci. 2011, 58, 145–147.
- Giotis, C.; Markellou, E.; Theodoropoulou, A.; Kostoulas, G.; Wilcockson, S.; Leifert, C. The effects of different biological control agents (BCAs) and plant defence elicitors on cucumber powdery mildew (Podosphera xanthii). Org. Agric. 2012, 2, 89–101.
- Dafermos, N.G.; Kasselaki, A.M.; Goumas, D.E.; Eyre, M.D.; Spandidakis, K.; Leifert, C. Integration of Elicitors and Less Susceptible Hybrids for the Control of Powdery Mildew in Organic Tomato Crops. Plant Dis. 2012, 96, 1506–1512.
- Margaritopoulou, T.; Kizis, D.; Kotopoulis, D.; Papadakis, I.E.; Anagnostopoulos, C.; Baira, E.; Termentzi, A.; VichouLeifert, C.; Markellou, E. Enriched HeK4me3 marks at Pm-0 resistance-related genes prime courgette against Podosphaera xanthii. Plant Physiol. 2021, kiab453.
- Barratt, B.I.P.; Moran, V.C.; Bigler, F.; van Lenteren, J.C. The status of biological control and recommendations for improving uptake for the future. Biocontrol 2018, 63, 155–167.
- Thambugala, K.M.; Daranagama, D.A.; Phillips, A.J.L.; Kannangara, S.D.; Itthayakor, P. Fungi vs. Fungi in Biocontrol: An Overview of Fungal Antagonists Applied Against Fungal Plant Pathogens. Front. Cell. Infect. Microbiol. 2020, 10, 604923.
- Köhl, J.; Kolnaar, R.; Ravensberg, W.J. Mode of Action of Microbial Biological Control Agents Against Plant Diseases: Relevance Beyond Efficacy. Front. Plant Sci. 2019, 10, 00845.
- Pesticide Action Network. Is Organic Better? 2017. Available online: www.pan-uk.org/organic/ (accessed on 1 June 2021).
- Kuesel, R.; Scott Hicks, D.; Archer, K.; Sciligo, A.; Bessin, R.; Gonthier, D. Effects of Fine-Mesh Exclusion Netting on Pests of Blackberry. Insects 2019, 10, 249.
- Gogo, E.O.; Saidi, M.; Itulya, F.M.; Martin, T.; Mathieu Ngouajio, M. Eco-Friendly Nets and Floating Row Covers Reduce Pest Infestation and Improve Tomato (Solanum lycopersicum L.) Yields for Smallholder Farmers in Kenya. Agronomy 2014, 4, 1–12.
- Mahmood, A.; Hu, Y.; Tanny, J.; Asante, E.A. Effects of shading and insect-proof screens on crop microclimate and production: A review of recent advances. Sci. Hortic. 2018, 241, 241–251.
- Savoldelli, S.; Trematerra, P. Mass-trapping, mating-disruption and attracticide methods for managing stored-product insects: Success stories and research needs. Stewart Postharvest Rev. 2011, 7, 1–8.
- Volakakis, N.; Kabourakis, E.; Leifert, C. Conventional and organic cultivation and their effect on the functional composition of olive oil. In Olives and Olive Oil as Functional Foods; Kiritsakis, A., Shahidi, F., Eds.; Wiley: Oxford, UK, 2017; pp. 35–44.
- Volakakis, N. Development of Strategies to Improve the Quality and Productivity of Organic and ‘Low Input’ Olive Production Systems in Semi-Arid Mediterranean Regions. Ph.D. Thesis, Newcastle University, Newcastle Upon Tyne, UK, 2010.
- Nikolouli, K.; Sassù, F.; Mouton, L.; Stauffer, C.; Bourtzis, K. Combining sterile and incompatible insect techniques for the population suppression of Drosophila suzukii. J. Pest Sci. 2020, 93, 647–661.
- Baker, T.C.; Zhu, J.J.; Millar, J.G. Delivering on the promise of pheromones. J. Chem. Ecol. 2016, 42, 553–556.
- Marliac, G.; Penvern, S.; Barbier, J.M.; Lescourret, F.; Capowiez, Y. Impact of crop protection strategies on natural enemies in organic apple production. Agron. Sustain. Dev. 2015, 35, 803–813.
- van Loenen, M.C.A.; Turbett, Y.; Mullins, C.E.; Feilden, N.E.H.; Wilson, M.J.; Leifert, C.; Seel, W. Low temperature-short duration steaming of soils kills soil-borne pathogens, nematodes and weeds. Eur. J. Plant Pathol. 2003, 109, 993–1002.
- Williams, J.S.; Cooper, R.M. The oldest fungicide and newest phytoalexin—A reappraisal of the fungitoxicity of elemental sulphur. Plant Pathol. 2004, 53, 263–279.
- Komárek, M.; Čadková, E.; Chrastný, V.; Bordas, F.; Bollinger, J.-C. Contamination of vineyard soils with fungicides: A review of environmental and toxicological aspects. Environ. Int. 2010, 36, 138–151.
- Ndakidemi, B.; Mtei, K.; Ndakidemi, P.A. Impacts of Synthetic and Botanical Pesticides on Beneficial Insects. Agric. Sci. 2016, 7, 364–372.
- Zanuncio, J.C.; Abreu Mourão, S.; Martínez, L.C.; Wilcken, C.F.; Ramalho, F.S.; Plata-Rueda, A.; Alvarenga Soares, M.; Serrão, J.E. Toxic effects of the neem oil (Azadirachta indica) formulation on the stink bug predator, Podisus nigrispinus (Heteroptera: Pentatomidae). Sci. Rep. 2016, 6, 30261.
- EFSA. Peer review of the pesticide risk assessment of the active substance spinosad. EFSA J. 2018, 16, 5252.
- Chrustek, A.; Hołyńska-Iwan, I.; Dziembowska, I.; Bogusiewicz, J.; Wróblewski, M.; Cwynar, A.; Olszewska-Słonina, D. Current research on the safety of pyrethroids used as insecticides. Medicina 2018, 54, 61.
- EFSA. The 2018 European Union Report on Pesticide Residues in Food. EFSA J. 2020, 18, 6057. Available online: https://efsa.onlinelibrary.wiley.com/doi/10.2903/j.efsa.2020.6057 (accessed on 4 February 2021).
- Markantonis, M.; van der Velde-Koerts, T.; Graven, C.; te Biesebeek, J.D.; Zeilmaker, M.; Rietveld, A.G.; Ossendorp, B.C. Assessment of occupational and dietary exposure to pesticide residues. EFSA J. 2018, 16, 16087.
- Bjørling-Poulsen, M.; Andersen, H.R.; Grandjean, P. Potential developmental neurotoxicity of pesticides used in Europe. Environ. Health 2008, 7, 50.
- EFSA. Evaluation of the data on clothianidin, imidacloprid and thiamethoxam for the updated risk assessment to bees for seed treatments and granules in the EU. EFSA Support. Publ. 2018, 15, 1378.
- Schug, T.T.; Janesick, A.; Blumberg, B.; Heindel, J.J. Endocrine disrupting chemicals and disease susceptibility. J. Steroid Biochem. Mol. Biol. 2011, 12, 204–215.
- Costas, L.; Infante-Rivard, C.; Zock, J.P.; Van Tongeren, M.; Boffetta, P.; Cusson, A.; Robles, C.; Casabonne, D.; Benavente, Y.; Becker, N.; et al. Occupational exposure to endocrine disruptors and lymphoma risk in a multi-centric European study. Br. J. Cancer 2015, 112, 1251–1256.
- Bonde, J.P.; Meulengracht Flachs, E.; Rimborg, S.; Glaser, C.H.; Giwercman, A.; Ramlau-Hansen, C.H.; Sørig Hougaard, K.; Høyer, B.B.; Keglberg Hærvig, K.; Bondo Petersen, S.; et al. The epidemiologic evidence linking prenatal and postnatal exposure to endocrine disrupting chemicals with male reproductive disorders: A systematic review and meta-analysis. Hum. Reprod. Update 2017, 23, 104–125.
- Nicolopoulou-Stamati, P.; Maipas, S.; Kotampasi, C.; Stamatis, P.; Hens, L. Chemical pesticides and human health: The urgent need for a new concept in agriculture. Front Public Health 2016, 4, 148.
- Burns, C.J.; McIntosh, L.J.; Mink, P.J.; Jurek, A.M.; Li, A.A. Pesticide exposure and neurodevelopmental outcomes: Review of the epidemiologic and animal studies. J. Toxicol. Environ. Health B Crit. Rev. 2013, 16, 127–283.
- Smith-Spangler, C.; Brandeau, M.L.; Hunter, G.E.; Clay Bavinger, J.; Pearson, M.; Eschbach, P.J.; Sundaram, V.; Liu, H.; Schirmer, P.; Stave, C.; et al. Are organic foods safer or healthier than conventional alternatives? A systematic review. Ann. Intern. Med. 2012, 157, 348–366.
- Mie, A.; Andersen, H.R.; Gunnarsson, S.; Kahl, J.; Kesse-Guyot, E.; Rembiałkowska, E.; Quaglio, G.; Grandjean, P. Human health implications of organic food and organic agriculture: A comprehensive review. Environ. Health 2017, 16, 111.
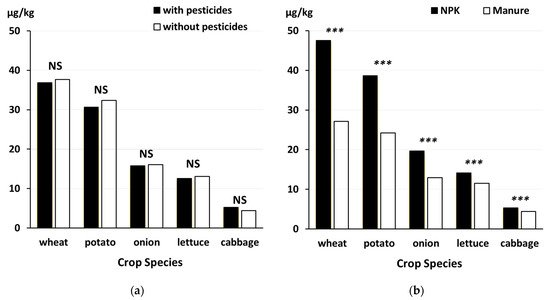
- Rempelos, L.; Wang, J.; Baranski, M.; Watson, A.; Volakakis, N.; Hoppe, H.-W.; Kühn-Velten, W.N.; Hadall, C.; Hasanaliyeva, G.; Chatzidimitriou, E.; et al. Diet and food type affect urinary pesticide residue excretion profiles in healthy individuals: Results of a randomized controlled dietary intervention trial. Am. J. Clin. Nutr. 2021, nqab308.
- Wang, J.; Hasanalieva, G.; Wood, L.; Anagnostopoulos, C.; Ampadogiannis, G.; Bempelou, E.; Kiousi, M.; Markellou, E.; Iversen, P.O.; Seal, C.; et al. Effect of wheat species (Triticum aestivum vs T. spelta), farming system (organic vs conventional) and flour type (wholegrain vs white) on composition of wheat flour—Results of a retail survey in the UK and Germany—3. Pesticide residue content. Food Chem. X 2020, 6, 100089.
- Średnicka-Tober, D.; Baranski, M.; Gromadzka-Ostrowska, J.; Skwarło-Sońta, K.; Rembiałkowska, E.; Hajslova, J.; Schulzova, V.; Cakmak, I.; Ozturk, L.; Krolikowski, T.; et al. Effect of crop protection and fertilization regimes used in organic and conventional production systems on feed composition and selected physiological parameters in rats. J. Agric. Food Chem. 2013, 61, 1017–1029.
- Baranski, M.; Średnicka-Tober, D.; Rempelos, L.; Hasanaliyeva, G.; Gromadzka-Ostrowska, J.; Skwarło-Sońta, K.; Królikowski, T.; Rembiałkowska, E.; Hajslova, J.; Schulzova, V.; et al. Feed Composition Differences Resulting from Organic and Conventional Farming Practices Affect Physiological Parameters in Wistar Rats—Results from a Factorial, Two-Generation Dietary Intervention Trial. Nutrients 2021, 13, 377.
- Rahman, A. Effect of soil organic matter on the phytotoxicity of soil applied herbicides—Glasshouse studies. N. Z. J. Exp. Agric. 1976, 4, 85–88.
- Jones, R.L.; Norris, F.A. Factors affecting degradation of aldicarb and ethoprop. J. Nematol. 1998, 30, 45–55.
- Kirchmann, H.; Haberhauer, G.; Kandeler, E.; Sessitsch, A.; Gerzabek, M.H. Effects of level and quality of organic matter input on carbon storage and biological activity in soil: Synthesis of a long-term experiment. Global Biogeochem. Cycles 2004, 18, GB4011.
- Lori, M.; Symnaczik, S.; Mader, P.; De Deyn, G.; Gattinger, A. Organic farming enhances soil microbial abundance and activity-A meta-analysis and meta-regression. PLoS ONE 2017, 12, 0180442.
- Harkes, P.; Suleiman, A.K.A.; van den Elsen, S.J.J.; de Haan, J.J.; Holterman, M.; Kuramae, E.; Helder, J. Conventional and organic soil management as divergent drivers of resident and active fractions of major soil food web constituents. Sci. Rep. 2019, 9, 13521.
- Saladin, G.; Clément, C. Physiological side effects of pesticides on non-target plants. In Agriculture and Soil Pollution; Livingston, J.V., Ed.; Nova Science Publishers Inc.: Hauppauge, NY, USA, 2005; pp. 53–86.
- McCurdy, J.D.; McElroy, J.S.; Kopsell, D.A.; Sams, C.E.; Sorochan, J.C. Effects of Mesotrione on Perennial Ryegrass (Lolium perenne L.) Carotenoid Concentrations under Varying Environmental Conditions. J. Agric. Food Chem. 2008, 56, 9133–9139.
- Marcelo, P.; Gomes, E.; Smedbol, A.; Chalifour, L.; Hénault-Ethier, M.; Labrecque, L.; Lepage, M.; Lucotte, P.J. Alteration of plant physiology by glyphosate and its by-product aminomethylphosphonic acid: An overview. J. Exp. Bot. 2014, 65, 4691–4703.
- Kanissery, R.; Gairhe, B.; Kadyampakeni, D.; Batuman, O.; Alferez, F. Glyphosate: Its Environmental Persistence and Impact on Crop Health and Nutrition. Plants 2019, 8, 499.
- Cakmak, I.; Yazici, A.; Tutus, Y.; Ozturk, L. Glyphosate reduced seed and leaf concentrations of calcium, manganese, magnesium, and iron in non-glyphosate resistant soybean. Eur. J. Agron. 2009, 31, 114–119.
- Bott, S.; Tesfamariam, T.; Candan, H.; Cakmak, I.; Römheld, V.; Neumann, G. Glyphosate-induced impairment of plant growth and micronutrient status in glyphosate-resistant soybean (Glycine max L.). Plant Soil 2008, 312, 185–194.
- Martinez, D.A.; Loening, U.E.; Graham, M.C. Impacts of glyphosate-based herbicides on disease resistance and health of crops: A review. Environ. Sci. Eur. 2018, 30, 2.
- Speiser, B.; Tamm, L.; Amsler, T.; Lambion, J.; Bertrand, C.; Hermansen, A.; Ruissen, M.A.; Haaland, P.; Zarb, J.; Santos, J.; et al. Field tests of blight control methods for organic farming: Tolerant varieties and copper fungicides. Biol. Agric. Hortic. 2006, 23, 393–412.
- Wolfe, M.S.; Baresel, J.P.; Desclaux, D.; Goldringer, I.; Hoad, S. Developments in breeding cereals for organic agriculture. Euphytica 2008, 163, 323.
- Lammerts van Bueren, E.T.; Jones, S.S.; Tamm, L.; Murphy, K.M.; Myers, J.R.; Leifert, C.; Messmer, M.M. The need to breed crop varieties suitable for organic farming, using wheat, tomato and broccoli as examples: A review. NJAS-Wagen. J. Life Sci. 2010, 58, 193–205.
- Crespo-Herrera, L.A.; Ortiz, R. Plant breeding for organic agriculture: Something new? Agric. Food Secur. 2015, 4, 1–7.
- Lammerts van Bueren, E.T.; Thorup-Kristensen, K.; Leifert, C.; Cooper, J.M.; Becker, H.C. Breeding for nitrogen efficiency; concepts, methods, and case studies. Euphytica 2014, 199, 1–2.
- Nuijten, E.; Messmer, M.M.; Lammerts van Bueren, E.T. Concepts and Strategies of Organic Plant Breeding in Light of Novel Breeding Techniques. Sustainability 2017, 9, 18.
- Lammerts van Bueren, E.T.; Myers, J.R. Organic Crop Breeding; Wiley-Blackwell: Hoboken, NY, USA, 2012.