Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 2 by Catherine Yang and Version 1 by Elisa Catanzaro.
Cholangiocarcinoma (CCA) is the most common malignancy in patients with primary sclerosing cholangitis (PSC), accounting for 2–8% of cases and being the leading cause of death in these patients. The majority of PSC-associated CCAs (PSC-CCA) develop within the first few years after PSC diagnosis. Older age and male sex, as well as concomitant inflammatory bowel disease (IBD) or high-grade biliary stenosis, are some of the most relevant risk factors. PSC-CCA pathogenesis is characterised by peculiar molecular and genetic features, being a distinct disease from the novo CCA. In recent years, tThere has been a significant push to develop innovative strategies for PSC-CCA early diagnosis and surveillance.
- cholangiocarcinoma
- primary sclerosing cholangitis
- pathogenesis
- diagnosis
1. Introduction
Cholangiocarcinoma (CCA) has always represented a challenging tumor with poor diagnostic and therapeutic tools and associated very low patient survival. Currently, CCA represents around 15% of all primary liver cancers, with an increasing trend during the last decades due to better awareness of the disease and diagnostic advances, accounting for 3% of all gastrointestinal neoplasms [1,2,3][1][2][3]. CCA is a part of a heterogeneous group of tumors, arising from any tract of the biliary tree, characterized by various genetic mutations, histological subtypes, and clinical presentations. Many risk factors could contribute to CCA development, and some of them are strongly associated with specific geographic areas. Flukes infections (Opisthorchis viverrini and Clonorchis sinensis) represent a major risk factors in the South Asian countries, explaining the higher incidence of CCA in this geographical area (6/100,000 cases per year) [1,3,4][1][3][4]. Incidence in western countries is significantly lower (<4/100,000 cases), and CCA is associated with some risk factors such as viral hepatitis (HBV, HCV) and metabolic dysfunction-associated steatotic liver disease (MASLD), but mostly the presence of an underlying biliary disease. Primary sclerosing cholangitis (PSC) represents the main risk factor for CCA in the western countries, followed by other conditions such as congenital bile-duct cysts, Caroli disease, or choledocholithiasis [5,6,7][5][6][7]. PSC is a rare cholestatic disease affecting the biliary tract, with formation of fibrotic biliary strictures and consequent dilatations along the biliary tree [8]. The high risk of developing hepatobiliary malignancies in PSC is well known and also involves other type of neoplasms, such as gallbladder carcinoma (GBC) and hepatocellular carcinoma (HCC). Moreover, the association with inflammatory bowel diseases (IBD) in up to 80% of PSC patients increases their oncological risk, as they are more prone in developing colorectal carcinoma (CRC). Regarding PSC-CCA, the pathogenetic mechanisms are not fully understood; immune-mediated hits and bile stasis might lead to chronic inflammation and cancerogenesis [9]. PSC-associated CCA (PSC-CCA) is characterized by peculiar epidemiological, clinical, molecular, and genetic features, arising the suspicion of PSC-CCA being a distinct disease from de novo CCA.
2. Pathogenesis
The most supported model of biliary carcinogenesis is the “multistep carcinogenesis” model. Presence of chronic inflammation leads to the sequential progression from damaged normal biliary epithelium to low-grade dysplasia (LGD), high-grade dysplasia (HGD), and finally invasive cancer [53,54,55][10][11][12]. A higher frequency of metaplasia and dysplasia was observed in the explanted liver of patients who developed PSC-CCA than PSC alone, suggesting that these histological findings could anticipate CCA development [43,53,54][10][11][13]. Not all types of metaplasia have the same transformative potential, as intestinal metaplasia seems to be more typical of PSC-CCA in comparison with other forms of metaplasia (mucinous, pyloric, etc.) that equally develop in both presence and absence of an underlying PSC. Intestinal metaplasia represents a significant predictor of both dysplasia and CCA, confirming the existence of a multistep process of neoplastic transformation and partially differentiating the pathogenesis of PSC-CCA from CCA alone. This observation could explain the different epidemiology PSC-CCA compared with de novo CCA and the earlier development of PSC-CCA [53][10]. Nevertheless, the role of inflammation in PSC and its progression to neoplastic transformation is still unknown, as well as the pathogenesis of the disease itself. Inflammation in PSC develops on a complicated interplay between immune-mediated mechanisms and external biliary insults from exogenous antigens (infectious, bile etc.), as shown in Figure 1.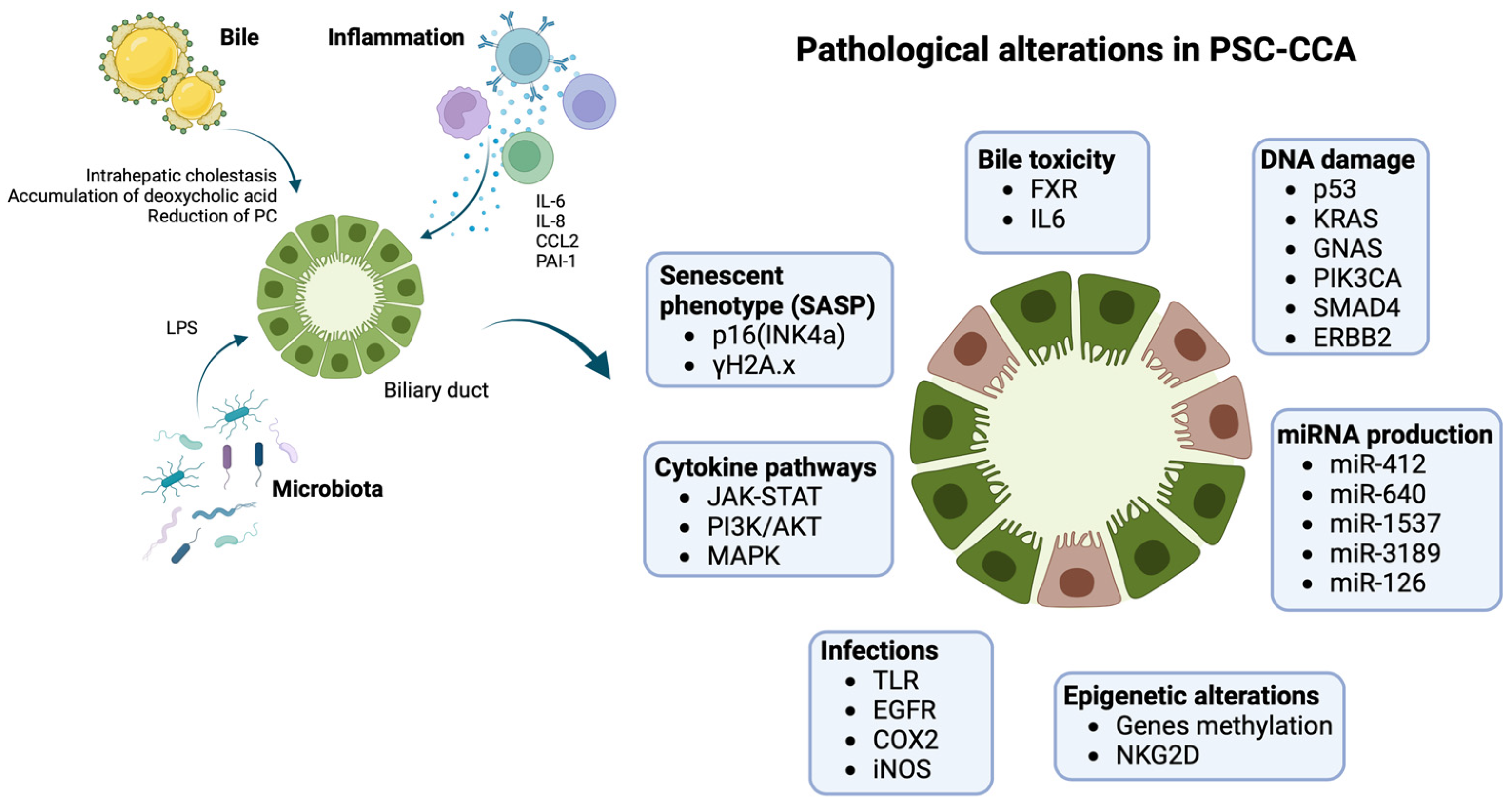
Figure 1. Pathogenesis of PSC-CCA: biliary ducts in PSC are exposed to several pathogenetic triggers, including impaired bile composition, infections, chronic inflammation, ending in damage cholangiocytes on several molecular levels. Inflammation is mediated by different cytokines (IL6, IL8, CCL2) activating pathways such as JAK-STAT, PI3K/AKT, and MAPK, with a proliferative and anti-apoptotic effect on cells. Chronic cholestasis and infections progressively generate up-regulation of iNOS with oxidative stress and stimulation of EGFR and COX2 with increased cellular growth, resistance to apoptosis, and reduction in DNA repair mechanisms. Cholestasis also has transcriptional effects with down-regulation of FXR and reduction of its chemoprotective effect. PSC-CCA is associated with several miRNAs’ dysregulated expression and epigenetic changes that interfere with normal cellular transcription. The effect is the development of an aberrant senescent phenotype. Senescent cholangiocytes are more prone to DNA damage, usually involving oncogenes and oncosuppressors (K-ras, p53), leading to cellular cycle escape and developing of CCA. Images created with BioRender.com, accessed on 18 September 2023.
3. Surveillance of CCA in PSC
The silent development of this neoplasia, typically asymptomatic in early stages, is in contrast with its aggressive behavior and rapid progression. This characteristic makes it difficult to achieve early diagnosis, losing the therapeutical window for curative treatment and compromising patient outcomes, due to limited systemic treatment options. In PSC patients, the radiological surveillance has a crucial role to detect early CCA and to allow curative treatments, such as liver resection or transplantation in patients who are usually young and fit for surgery. An ideal surveillance strategy should be simple, cheap, accepted by the patient, non-invasive, and with high sensitivity, aiming to increase overall survival. However, no diagnostic tests have those characteristics [33][53], and high-grade dysplasia or early-stage CCA are usually diagnosed using invasive techniques like ERCP, keeping in mind that the risk of complications in ERCP is not justified by the sensitivity of the endoluminal sampling, that is usually low [123][54]. The positive impact of radiological surveillance for PSC patients has already been proven by different studies, reporting better outcomes with a reduction in hepatobiliary cancers-related deaths in patients exposed to at least a routinary imaging technique [21,146][55][56]. Also, the large multi-center study from the IPSCSG proved that scheduled imaging was associated with improved overall survival [33][53]. Magnetic resonance imaging (MRI) with magnetic resonance cholangiopancreatography (MRCP) is the preferred radiologic modality with no radiation and the possibility to supply more detailed and accurate information than abdominal (US) [25][57]. The guidelines suggested yearly MRI/MRCP for CCA surveillance in large-duct PSC patients. Pediatric and sdPSC patients represent low-risk categories, and surveillance is not strictly indicated for them [47,98][58][59]. There is not a consensus on the correct timing of scheduled imaging for CCA surveillance in PSC, and some experts suggested a limited role of imaging surveillance for biliary malignancies [147][60]. Recently, a prospective study from Villard et al., including 512 unselected patients followed for 5 years with annual MRI/MRCP, detected CCA in 2% of patients, associated with previous development of progressive and severe biliary strictures. Although 62% of them were eligible for curative treatment, all of them except one developed tumor recurrence and died during the follow-up, pointing out the inability of annual surveillance to detect cancer early enough to provide long-term survival [32][61]. Moreover, the failing of the guidelines’ strategy has been evident in the group of patients with severe strictures, suggesting how the future direction of research should be the individualization of surveillance, reserving closer MRI/MRCP controls in selected high-risk groups (recent or long-date diagnosis of large-duct PSC, older age, or severe and symptomatic disease). The use of scheduled ERCP associated with MRI/MRCP does not improve the surveillance efficacy [33[53][62],48], and PSC itself represents a risk factor for ERCP-associated complications [148][63]. Scheduled ERCP in PSC could have a beneficial effect in the treatment of high-grade stenosis, regardless of patient symptoms according to recent data [149][64], but does not represent an adequate tool for surveillance. Abdominal US remains the cornerstone for the surveillance of the other types of hepatobiliary neoplasms occurring in PSC; surveillance of GBC requires a 12-month US surveillance, which could be replaced with or added to the annual MRI/MRCP for CCA surveillance if already performed. A 6-month US is mandatory in the presence of cirrhotic evolution for HCC surveillance [47,98][58][59]. The majority of PSC international guidelines discouraged the 6-month use of CA19-9 for CCA surveillance [35,47,98,104][58][59][65][66] because it has a low positive predictive value and a low accuracy [117][67]. Indeed, CA19-9 is highly variable and many other benign or malignant conditions, as well as interpersonal variability, could influence the serum levels of these markers. However, the rapid increase of CA 19-9 has been shown to predict malignancies regardless of the cut-off in use, and some experts have tried to narrow the interval of assessment of the biomarker (every 2 or 3 months) in order to detect the progressive CA19-9 change rather than focus on its absolute value [107][68]. The risk of over-diagnosis associated with this strategy has not been evaluated yet, and the potentially higher rate of false positives could remarkably affect the costs of the surveillance program.4. Future Perspective: Next-Generation Biomarkers for PSC-CCA Diagnosis and Surveillance
The pancreatic autoantibody against glycoprotein 2 (anti-GP2 IgA) had been indicated as a novel marker in large-duct PSC. It represents a prognostic marker associated with a more severe disease and dismal prognosis with a higher risk of developing CCA [150][69]. Another observation on biochemical markers reveals that a high enhanced liver fibrosis (ELF) score correlates with patients developing CCA sporadically or on PSC. A following multivariate analysis also proved that ELF score was correlated to CCA independently of liver disease stage or patients’ age, suggesting this marker to be related to the biological nature of tumor development [151][70]. This follows that some markers could be a useful tool for patient risk stratification, being able to identify patients with a higher risk of developing PSC-CCA. This could guide to a more individualized surveillance strategy in PSC. A recent study from Cuenco et al. analyzed the diagnostic performance of a panel of new biomarkers including pyruvate kinase M2 (PKM2), cytokeratin 19 fragment (CYFRA21.1) and mucin 5AC (MUC5AC) achieving promising results with an high specificity (90%) but mostly a high sensitivity when combined with gamma-glutamyltransferase (GGT) (82%) [152][71]. Also, osteopontin, usually upregulated in CCA tumor cells, was seen to be higher in the serum of patients with CCA compared to healthy controls. During the exploratory analysis of the study from Loosen et al., some 10 PSC patients and 13 healthy controls were compared with 27 patients with CCA, revealing significantly elevated osteopontin levels in CCA patients compared to serum samples from healthy and PSC patients, suggesting a possible role in differentiating among PSC patients those who develop biliary malignancy [153][72]. Unfortunately, no PSC patients were included in the final analysis, and the observation remains limited to the small exploratory cohort. A study on the use of volatile organic compounds (VOCs) measurement in bile showed interesting results, revealing differences in gas concentrations between PSC and PSC-CCA patients [154][73], as already observed in other types of cancer [155][74]. The subsequent study from the same group of Navaneethan et al. also showed accuracy of VOCs in distinguishing CCA from PSC patients when measured in urine. Some VOCs like ethane and 1-octene reached an 80% sensitivity at 100% specificity, and also 2-propanol and acetonitrile showed promising results, representing a potential non-invasive marker. In the future, the possibility of translating VOCs measurement to breath analysis might be tested [156][75]. DNA hypermethylation of gene promoters represents an early event in biliary carcinogenesis. The methylation of CDKN2A (p16) represents a frequent DNA alteration [157][76], but also other loci were found to be hypermethylated in CCA samples [158,159[77][78][79],160], representing potential markers for PSC-CCA detection. However, only a few studies included PSC patients, and data on PSC-CCA are limited. The analysis of methylation status of some candidate genes (cysteine dioxygenase type 1 (CDO1), cannabinoid receptor interacting protein 1 (CNRIP1), septin 9 (SEPT9), and vimentin (VIM)) on biliary brushing of CCA and PSC patients revealed good performances in discriminating malignancy from the benign and inflammatory-related biliary changes in PSC, with high sensitivity (85%) and specificity (98%) [161][80]. A more recent study tried to validate the measurement of methylation level of the same gene panel in bile, using droplet digital PCR (ddPCR) for the analysis [162][81]. Previous studies on bile samples could not find a significative association between hypermethylation of specifical genes (CDKN2A) and PSC-CCA [163][82], or measured potential biomarkers only in non-PSC patients (like CCND2, CDH13, GRIN2B, RUNX3, and TWIST) [164][83]. Instead, the three center-based (Sweden, Norway, Finland) study from Vedelet et al. observed the gene methylation panel including CDO1, CNRIP1, SEPT9, and VIM to be accurate for early PSC-CCA detection, with an AUC of 0.88. The results were even enhanced including only patients with early PSC-CCA diagnosis (≤12 months) and PSC controls with long follow-up (>36 months), reaching a sensitivity and specificity of 100% and 93%, respectively, and an AUC of 0.98. This promising result proved that DNA methylation biomarkers in bile has the potential to complement standard diagnostic modalities for early PSC-CCA detection [162][81]. Prospective validation of this study in larger multicenter sample series is warranted. However, the possibility to include this methylation analysis in CCA surveillance in PSC patients remains controversial due to the invasive nature of bile sampling. A few studies also performed DNA methylation analysis on blood samples, discovering some potential biomarkers, already tested in tissue samples [165][84] (SHOX2, SEPT9, OPCML, HOXD9, etc.), but unfortunately no PSC patients had been included [166,167][85][86]. The analysis of micro ribonucleic acid (miRNAs) was highly investigated. miRNAs represent non-coding small nucleic acids capable of binding to specifical messenger RNA (mRNAs) and regulating gene expression [168,169][87][88]. Several miRNAs share an important role in chronic cholestasis, interfering with several cellular processes including proliferation and apoptosis. Impaired levels of miRNAs could result in dysregulating cellular cycle generating clonal expansion and eventually cancer. Interestingly, miRNAs expression is not directly altered by cholestasis; indeed, presence of high levels of bilirubin does not influence the result of miRNAs analysis [93][52]. As already explained above, bile levels of some miRNAs (miR-640, miR-3189, miR-1537, and miR-412) are significantly higher in PSC-CCA patients compared with PSC-alone patients, representing a potential diagnostic tool in early diagnosis. Another study from Li et al. aimed to investigate potential biomarkers for CCA diagnosis regardless of PSC, showing the diagnostic value of an miRNA panel specifically associated with CCA with good sensitivity of 67% and high specificity of 96%. The study included a few PSC patients in the control group, suggesting their possible role in differentiating the miRNA pattern [170][89]. This aspect needs to be considered in future studies. Some differences were found between miRNAs levels of PSC and CCA-alone patients (miR-1281, miR-126, miR-26a, miR-30b, and miR-122). Nevertheless, a consistent overlapping expression of most miRNAs in PSC and CCA patients was observed. Only miR-126 gave considering results, but only reached a specificity of 93% with poor sensitivity, comparable with brushing cytology [93][52]. However, another study of miRNA in serum showed a difference in the expression of miR-222 and miR-483-5p in CCA versus PSC controls [171][90]. Other miRNAs (miR-21, miR-221, miR-122, miR-192, miR-29b, and miR-155) were demonstrated to be associated with CCA in terms of both diagnosis and prognosis; however, control groups of these studies count only a few or no PSC patients [172,173][91][92]. Future research is required to investigate on serum miRNAs detection, being less invasive compared to bile sampling. Combined strategies should be implemented to include more information from both serum and bile and integrate them with other biomarkers. Neoplastic cells are characterized by high metabolism with production of several potential biomarkers. Indeed, metabolic alteration could reflect on impaired concentration of circulating metabolites in the presence of cancer. Specific alteration at the metabolome analysis had been already described in other neoplastic diseases [174,175][93][94]. According to that, the group of Banales et al. performed a metabolomic analysis of 424 different metabolites in serum to identify a significant concentration of certain circulating metabolites in patients who developed hepatic malignancy, in particular iCCA or hepatocellular carcinoma (HCC) compared with PSC. Some metabolites were significantly different in the PSC and iCCA group and the algorithms combining some of them, histidine and PC(34:3), could accurately differentiate between the two diagnoses [176][95]. This suggests a different metabolic profile of CCA and PSC that could represent a starting point for further analysis on PSC-CCA patients. Promising results also came from proteomic analysis. In particular, urine proteomic analysis reveals the capacity to differentiate CCA from PSC [177][96]. The use of urine proteomic analysis showed better results when associated with biliary proteomic analysis, which also provides good discrimination capability in the field [178][97]. The combination of bile and urine proteomic analysis had been explored in a retrospective study and then applied to a prospective analysis, improving sensitivity of CCA diagnosis, compared with standard diagnostic approach, from 72% to 94%. A large prospective trial on patients with progressive cholestasis during surveillance is required to confirm the accuracy of this method for the early diagnosis of CCA [179][98]. In serum, a glycomic and proteomic analysis on a particular set of proteins was able to identify some differences in the expression of certain glycans in patients with CCA. Among them, fucosylated fetuin A (fc-fetuin A) was capable of differentiating CCA from those with PSC, suggesting a possible role of this analysis in the surveillance of PSC patients [180][99]. An early field of liquid biopsy-based research is the analysis of extracellular vesicles (EVs) and their proteomic profile in serum samples. EVs contain biomolecules of various nature, like lipids, proteins, and nucleic acids, and different cells, including malignant cholangiocytes, release EVs with specific possible biomarkers [181,182][100][101]. First evidence of specifical EVs proteomic patterns came from the analysis of tissue samples, defining several CCA-derived EVs, not in common with PSC, that may reflect tumor features and give prognostic information [183][102]. Lapitz et al. performed an analysis of EVs’ transcriptomic profile in urine and serum of CCA, showing a specific expression of messenger RNAs (mRNA) and non-coding RNAs (ncRNAs) in CCA patients compared to a mixed group of patients with PSC, UC, and healthy controls. In urine, in the partial analysis including only CCA (n = 23) vs. PSC (n = 5) patients, a higher expression of some specific mRNAs like CLIP3, VCAM1, and TRIM33 in CCA group (AUC 0.965) was reported, as well as a significant different expression of other transcripts like ATP5EP2, LOC100134713, and SNORA8 [184][103]. The same analysis in serum (CCA n = 12 vs. PSC n = 6) reveals mRNAs, PON1, ATF4, and PHGDH were the best candidate biomarkers for the differential diagnosis of CCA and PSC (AUCs 1.00), as well as a high accuracy for the identification of CCA vs. PSC of the transcripts MALAT1, LOC100190986, and SNORA11B (AUCs: 1.00) [184][103]. Subsequent gene ontology analysis indicated that most of the commonly altered mRNAs participate in carcinogenic pathways. Among EVs biomarkers, proteins also have a differential expression in CCA or PSC (FIBG, A1AG1, S10A8) according recent studies, and interestingly some of them have higher diagnostic values in early-stage CCA (I-II) (FCN2, ITIH4 and FIBG) [183][102]. A recent proteomic analysis of EVs in serum from the same study group individuates the combination of serum biomarkers CRP/FIBRINOGEN/FRIL as a valid tool for diagnosis of early CCA developing in PSC patients. Other biomarkers were associated with a prognostic role, predicting overall survival. Moreover, the integration of serum PIGR with CRP/FIBRINOGEN/FRIL showed the capacity for predicting CCA development in PSC patients with sensitivity and specificity of 64% and 93%, respectively (AUC 0.91), suggesting its potential use in surveillance settings in the future [185][104]. Anyway, further studies are needed to validate their effective role and the potential therapeutic strategies associated with their measurement. Next-generation sequencing (NGS) provides a multigene analysis with high analytical sensitivity and recently grew up with the advances in the characterization of the mutational profile of biliary tumors. Thanks to NGS, many recurrent mutations had been already identified in CCA [186][105], and some studies already tried to apply NGS for the research of novel biomarkers in de novo CCA diagnosis. A prospective study from Singhi et al. evaluated the use of NGS (a 28-gene panel renamed BiliSeq) in bile duct brushing and tissue sample obtained during ERCP. There was an improvement of sensitivity compared to standard clinical and pathological evaluation (73% vs. 48%). Notably, sensitivity of the BiliSeq panel was considerably higher (83%) in patients with underlying diagnosis of PSC [187][106]. More recently Arechederra et al. evaluated the use of NGS for the mutational analysis of bile cell-free DNA (cfDNA). The panel in use, called the “Bilemut assay”, achieved a sensitivity of 96.4% and a specificity of 69.2% [188][107]. The use of bile cfDNA could provide the advantage of obtaining nucleic acids released from all the CCA cells along the biliary tract, overcoming the limitation of the tissue sample and improving sensitivity. In general, the use of NGS for the identification of biomarkers could be useful not only for the early detection of malignancy but also for its prognostic role, being able to detect mutations that could become potential targets of oncological therapies. Further research will fill the gap made by the absence of specifical studies on PSC-CCA patients, testing the diagnostic and prognostic performances of NGS biomarkers panels in this group of high-risk patients.References
- Izquierdo-Sanchez, L.; Lamarca, A.; La Casta, A.; Buettner, S.; Utpatel, K.; Klümpen, H.-J.; Adeva, J.; Vogel, A.; Lleo, A.; Fabris, L.; et al. Cholangiocarcinoma Landscape in Europe: Diagnostic, Prognostic and Therapeutic Insights from the ENSCCA Registry. J. Hepatol. 2022, 76, 1109–1121.
- Banales, J.M.; Marin, J.J.G.; Lamarca, A.; Rodrigues, P.M.; Khan, S.A.; Roberts, L.R.; Cardinale, V.; Carpino, G.; Andersen, J.B.; Braconi, C.; et al. Cholangiocarcinoma 2020: The next Horizon in Mechanisms and Management. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 557–588.
- Bertuccio, P.; Malvezzi, M.; Carioli, G.; Hashim, D.; Boffetta, P.; El-Serag, H.B.; La Vecchia, C.; Negri, E. Global Trends in Mortality from Intrahepatic and Extrahepatic Cholangiocarcinoma. J. Hepatol. 2019, 71, 104–114.
- Kaewpitoon, N. Opisthorchis Viverrini: The Carcinogenic Human Liver Fluke. WJG 2008, 14, 666.
- Clements, O.; Eliahoo, J.; Kim, J.U.; Taylor-Robinson, S.D.; Khan, S.A. Risk Factors for Intrahepatic and Extrahepatic Cholangiocarcinoma: A Systematic Review and Meta-Analysis. J. Hepatol. 2020, 72, 95–103.
- Wongjarupong, N.; Assavapongpaiboon, B.; Susantitaphong, P.; Cheungpasitporn, W.; Treeprasertsuk, S.; Rerknimitr, R.; Chaiteerakij, R. Non-Alcoholic Fatty Liver Disease as a Risk Factor for Cholangiocarcinoma: A Systematic Review and Meta-Analysis. BMC Gastroenterol. 2017, 17, 149.
- Tyson, G.L.; El-Serag, H.B. Risk Factors for Cholangiocarcinoma. Hepatology 2011, 54, 173–184.
- Karlsen, T.H.; Folseraas, T.; Thorburn, D.; Vesterhus, M. Primary Sclerosing Cholangitis—A Comprehensive Review. J. Hepatol. 2017, 67, 1298–1323.
- Khan, S.A.; Tavolari, S.; Brandi, G. Cholangiocarcinoma: Epidemiology and Risk Factors. Liver Int. 2019, 39, 19–31.
- Lewis, J.T.; Talwalkar, J.A.; Rosen, C.B.; Smyrk, T.C.; Abraham, S.C. Precancerous Bile Duct Pathology in End-Stage Primary Sclerosing Cholangitis, with and without Cholangiocarcinoma. Am. J. Surg. Pathol. 2010, 34, 27–34.
- Fleming, K.A.; Boberg, K.M.; Glaumann, H.; Bergquist, A.; Smith, D.; Clausen, O.P.F. Biliary Dysplasia as a Marker of Cholangiocarcinoma in Primary Sclerosing Cholangitis. J. Hepatol. 2001, 34, 360–365.
- Bergquist, A.; Glaumann, H.; Stal, P.; Wang, G.-S.; Broome, U. Biliary Dysplasia, Cell Proliferation and Nuclear DNA-Fragmentation in Primary Sclerosing Cholangitis with and without Cholangiocarcinoma. J. Intern. Med. 2001, 249, 69–75.
- Chung, B.K.; Karlsen, T.H.; Folseraas, T. Cholangiocytes in the Pathogenesis of Primary Sclerosing Cholangitis and Development of Cholangiocarcinoma. Biochim. Biophys. Acta (BBA)—Mol. Basis Dis. 2018, 1864, 1390–1400.
- de Vries, A.B. Distinctive Inflammatory Bowel Disease Phenotype in Primary Sclerosing Cholangitis. WJG 2015, 21, 1956.
- Hirschfield, G.M.; Karlsen, T.H.; Lindor, K.D.; Adams, D.H. Primary Sclerosing Cholangitis. Lancet 2013, 382, 1587–1599.
- Özdirik, B.; Müller, T.; Wree, A.; Tacke, F.; Sigal, M. The Role of Microbiota in Primary Sclerosing Cholangitis and Related Biliary Malignancies. Int. J. Mol. Sci. 2021, 22, 6975.
- Saab, M.; Mestivier, D.; Sohrabi, M.; Rodriguez, C.; Khonsari, M.R.; Faraji, A.; Sobhani, I. Characterization of Biliary Microbiota Dysbiosis in Extrahepatic Cholangiocarcinoma. PLoS ONE 2021, 16, e0247798.
- Jia, X.; Lu, S.; Zeng, Z.; Liu, Q.; Dong, Z.; Chen, Y.; Zhu, Z.; Hong, Z.; Zhang, T.; Du, G.; et al. Characterization of Gut Microbiota, Bile Acid Metabolism, and Cytokines in Intrahepatic Cholangiocarcinoma. Hepatology 2020, 71, 893–906.
- Reich, M.; Spomer, L.; Klindt, C.; Fuchs, K.; Stindt, J.; Deutschmann, K.; Höhne, J.; Liaskou, E.; Hov, J.R.; Karlsen, T.H.; et al. Downregulation of TGR5 (GPBAR1) in Biliary Epithelial Cells Contributes to the Pathogenesis of Sclerosing Cholangitis. J. Hepatol. 2021, 75, 634–646.
- Hohenester, S.; Maillette de Buy Wenniger, L.; Paulusma, C.C.; van Vliet, S.J.; Jefferson, D.M.; Oude Elferink, R.P.; Beuers, U. A Biliary HCO3− Umbrella Constitutes a Protective Mechanism against Bile Acid-Induced Injury in Human Cholangiocytes. Hepatology 2012, 55, 173–183.
- Gauss, A. Biliary Phosphatidylcholine and Lysophosphatidylcholine Profiles in Sclerosing Cholangitis. WJG 2013, 19, 5454.
- Henriksen, E.K.K.; Viken, M.K.; Wittig, M.; Holm, K.; Folseraas, T.; Mucha, S.; Melum, E.; Hov, J.R.; Lazaridis, K.N.; Juran, B.D.; et al. HLA Haplotypes in Primary Sclerosing Cholangitis Patients of Admixed and Non-European Ancestry. HLA 2017, 90, 228–233.
- The International IBD Genetics Consortium (IIBDGC); International Genetics of Ankylosing Spondylitis Consortium (IGAS); International PSC Study Group (IPSCSG); Genetic Analysis of Psoriasis Consortium (GAPC); Psoriasis Association Genetics Extension (PAGE); Ellinghaus, D.; Jostins, L.; Spain, S.L.; Cortes, A.; Bethune, J.; et al. Analysis of Five Chronic Inflammatory Diseases Identifies 27 New Associations and Highlights Disease-Specific Patterns at Shared Loci. Nat. Genet. 2016, 48, 510–518.
- Folseraas, T.; Liaskou, E.; Anderson, C.A.; Karlsen, T.H. Genetics in PSC: What Do the “Risk Genes” Teach Us? Clin. Rev. Allergy Immunol. 2015, 48, 154–164.
- The UK-PSCSC Consortium; The International IBD Genetics Consortium; The International PSC Study Group; Liu, J.Z.; Hov, J.R.; Folseraas, T.; Ellinghaus, E.; Rushbrook, S.M.; Doncheva, N.T.; Andreassen, O.A.; et al. Dense Genotyping of Immune-Related Disease Regions Identifies Nine New Risk Loci for Primary Sclerosing Cholangitis. Nat. Genet. 2013, 45, 670–675.
- Alvaro, D.; Gigliozzi, A.; Attili, A.F. Regulation and Deregulation of Cholangiocyte Proliferation. J. Hepatol. 2000, 33, 333–340.
- Raven, A.; Lu, W.-Y.; Man, T.Y.; Ferreira-Gonzalez, S.; O’Duibhir, E.; Dwyer, B.J.; Thomson, J.P.; Meehan, R.R.; Bogorad, R.; Koteliansky, V.; et al. Cholangiocytes Act as Facultative Liver Stem Cells during Impaired Hepatocyte Regeneration. Nature 2017, 547, 350–354.
- Carpino, G.; Cardinale, V.; Renzi, A.; Hov, J.R.; Berloco, P.B.; Rossi, M.; Karlsen, T.H.; Alvaro, D.; Gaudio, E. Activation of Biliary Tree Stem Cells within Peribiliary Glands in Primary Sclerosing Cholangitis. J. Hepatol. 2015, 63, 1220–1228.
- DiPaola, F.; Shivakumar, P.; Pfister, J.; Walters, S.; Sabla, G.; Bezerra, J.A. Identification of Intramural Epithelial Networks Linked to Peribiliary Glands That Express Progenitor Cell Markers and Proliferate after Injury in Mice. Hepatology 2013, 58, 1486–1496.
- Guicciardi, M.E.; Trussoni, C.E.; LaRusso, N.F.; Gores, G.J. The Spectrum of Reactive Cholangiocytes in Primary Sclerosing Cholangitis. Hepatology 2020, 71, 741–748.
- Carpino, G.; Nevi, L.; Overi, D.; Cardinale, V.; Lu, W.; Di Matteo, S.; Safarikia, S.; Berloco, P.B.; Venere, R.; Onori, P.; et al. Peribiliary Gland Niche Participates in Biliary Tree Regeneration in Mouse and in Human Primary Sclerosing Cholangitis. Hepatology 2020, 71, 972–989.
- Clerbaux, L.-A.; Manco, R.; Van Hul, N.; Bouzin, C.; Sciarra, A.; Sempoux, C.; Theise, N.D.; Leclercq, I.A. Invasive Ductular Reaction Operates Hepatobiliary Junctions upon Hepatocellular Injury in Rodents and Humans. Am. J. Pathol. 2019, 189, 1569–1581.
- Sato, K.; Marzioni, M.; Meng, F.; Francis, H.; Glaser, S.; Alpini, G. Ductular Reaction in Liver Diseases: Pathological Mechanisms and Translational Significances. Hepatology 2019, 69, 420–430.
- Mederacke, I.; Hsu, C.C.; Troeger, J.S.; Huebener, P.; Mu, X.; Dapito, D.H.; Pradere, J.-P.; Schwabe, R.F. Fate Tracing Reveals Hepatic Stellate Cells as Dominant Contributors to Liver Fibrosis Independent of Its Aetiology. Nat. Commun. 2013, 4, 2823.
- Volckaert, T.; De Langhe, S. Lung Epithelial Stem Cells and Their Niches: Fgf10 Takes Center Stage. Fibrogenes. Tissue Repair. 2014, 7, 8.
- Tabibian, J.H.; O’Hara, S.P.; Splinter, P.L.; Trussoni, C.E.; LaRusso, N.F. Cholangiocyte Senescence by Way of N-Ras Activation Is a Characteristic of Primary Sclerosing Cholangitis: Hepatology. Hepatology 2014, 59, 2263–2275.
- Coppé, J.-P.; Desprez, P.-Y.; Krtolica, A.; Campisi, J. The Senescence-Associated Secretory Phenotype: The Dark Side of Tumor Suppression. Annu. Rev. Pathol. Mech. Dis. 2010, 5, 99–118.
- O’Hara, S.P.; Splinter, P.L.; Trussoni, C.E.; Guicciardi, M.E.; Splinter, N.P.; Al Suraih, M.S.; Nasser-Ghodsi, N.; Stollenwerk, D.; Gores, G.J.; LaRusso, N.F. The Transcription Factor ETS1 Promotes Apoptosis Resistance of Senescent Cholangiocytes by Epigenetically Up-Regulating the Apoptosis Suppressor BCL2L1. J. Biol. Chem. 2019, 294, 18698–18713.
- Boberg, K.M.; Schrumpf, E.; Bergquist, A.; Broomé, U.; Pares, A.; Remotti, H.; Schjölberg, A.; Spurkland, A.; Clausen, O.P.F. Cholangiocarcinoma in Primary Sclerosing Cholangitis: K-Ras Mutations and Tp53 Dysfunction Are Implicated in the Neoplastic Development. J. Hepatol. 2000, 32, 374–380.
- Kamp, E.J.; Dinjens, W.N.; Doukas, M.; Van Marion, R.; Verheij, J.; Ponsioen, C.Y.; Bruno, M.J.; Groot Koerkamp, B.; Trivedi, P.J.; Peppelenbosch, M.P.; et al. Genetic Alterations during the Neoplastic Cascade towards Cholangiocarcinoma in Primary Sclerosing Cholangitis. J. Pathol. 2022, 258, 227–235.
- Goeppert, B.; Folseraas, T.; Roessler, S.; Kloor, M.; Volckmar, A.; Endris, V.; Buchhalter, I.; Stenzinger, A.; Grzyb, K.; Grimsrud, M.M.; et al. Genomic Characterization of Cholangiocarcinoma in Primary Sclerosing Cholangitis Reveals Therapeutic Opportunities. Hepatology 2020, 72, 1253–1266.
- Ahrendt, S. Diagnosis and Management of Cholangiocarcinoma in Primary Sclerosing Cholangitis. J. Gastrointest. Surg. 1999, 3, 357–368.
- Yamada, D.; Rizvi, S.; Razumilava, N.; Bronk, S.F.; Davila, J.I.; Champion, M.D.; Borad, M.J.; Bezerra, J.A.; Chen, X.; Gores, G.J. IL-33 Facilitates Oncogene-induced Cholangiocarcinoma in Mice by an Interleukin-6-sensitive Mechanism. Hepatology 2015, 61, 1627–1642.
- Meng, F.; Yamagiwa, Y.; Ueno, Y.; Patel, T. Over-Expression of Interleukin-6 Enhances Cell Survival and Transformed Cell Growth in Human Malignant Cholangiocytes. J. Hepatol. 2006, 44, 1055–1065.
- Frampton, G.; Invernizzi, P.; Bernuzzi, F.; Pae, H.Y.; Quinn, M.; Horvat, D.; Galindo, C.; Huang, L.; McMillin, M.; Cooper, B.; et al. Interleukin-6-Driven Progranulin Expression Increases Cholangiocarcinoma Growth by an Akt-Dependent Mechanism. Gut 2012, 61, 268–277.
- Jaiswal, M.; LaRusso, N.F.; Burgart, L.J.; Gores, G.J. Inflammatory Cytokines Induce DNA Damage and Inhibit DNA Repair in Cholangiocarcinoma Cells by a Nitric Oxide-Dependent Mechanism. Cancer Res. 2000, 60, 184–190.
- Ehlken, H.; Schramm, C. Primary Sclerosing Cholangitis and Cholangiocarcinoma: Pathogenesis and Modes of Diagnostics. Dig. Dis. 2013, 31, 118–125.
- Finzi, L.; Shao, M.X.G.; Paye, F.; Housset, C.; Nadel, J.A. Lipopolysaccharide Initiates a Positive Feedback of Epidermal Growth Factor Receptor Signaling by Prostaglandin E2 in Human Biliary Carcinoma Cells. J. Immunol. 2009, 182, 2269–2276.
- Lozano, E.; Sanchez-Vicente, L.; Monte, M.J.; Herraez, E.; Briz, O.; Banales, J.M.; Marin, J.J.G.; Macias, R.I.R. Cocarcinogenic Effects of Intrahepatic Bile Acid Accumulation in Cholangiocarcinoma Development. Mol. Cancer Res. 2014, 12, 91–100.
- Melum, E.; Karlsen, T.H.; Schrumpf, E.; Bergquist, A.; Thorsby, E.; Boberg, K.M.; Lie, B.A. Cholangiocarcinoma in Primary Sclerosing Cholangitis Is Associated with NKG2D Polymorphisms. Hepatology 2007, 47, 90–96.
- Yang, H.; Li, T.W.H.; Peng, J.; Tang, X.; Ko, K.S.; Xia, M.; Aller, M. A Mouse Model of Cholestasis-Associated Cholangiocarcinoma and Transcription Factors Involved in Progression. Gastroenterology 2011, 141, 378–388.e4.
- Voigtländer, T.; Gupta, S.K.; Thum, S.; Fendrich, J.; Manns, M.P.; Lankisch, T.O.; Thum, T. MicroRNAs in Serum and Bile of Patients with Primary Sclerosing Cholangitis and/or Cholangiocarcinoma. PLoS ONE 2015, 10, e0139305.
- Bergquist, A.; Weismüller, T.J.; Levy, C.; Rupp, C.; Joshi, D.; Nayagam, J.S.; Montano-Loza, A.J.; Lytvyak, E.; Wunsch, E.; Milkiewicz, P.; et al. Impact on Follow-up Strategies in Patients with Primary Sclerosing Cholangitis. Liver Int. 2023, 43, 127–138.
- Majeed, A.; Castedal, M.; Arnelo, U.; Söderdahl, G.; Bergquist, A.; Said, K. Optimizing the Detection of Biliary Dysplasia in Primary Sclerosing Cholangitis before Liver Transplantation. Scand. J. Gastroenterol. 2018, 53, 56–63.
- Trivedi, P.J.; Crothers, H.; Mytton, J.; Bosch, S.; Iqbal, T.; Ferguson, J.; Hirschfield, G.M. Effects of Primary Sclerosing Cholangitis on Risks of Cancer and Death in People with Inflammatory Bowel Disease, Based on Sex, Race, and Age. Gastroenterology 2020, 159, 915–928.
- Ali, A.H.; Tabibian, J.H.; Nasser-Ghodsi, N.; Lennon, R.J.; DeLeon, T.; Borad, M.J.; Hilscher, M.; Silveira, M.G.; Carey, E.J.; Lindor, K.D. Surveillance for Hepatobiliary Cancers in Patients with Primary Sclerosing Cholangitis. Hepatology 2018, 67, 2338–2351.
- Eaton, J.E.; Welle, C.L.; Bakhshi, Z.; Sheedy, S.P.; Idilman, I.S.; Gores, G.J.; Rosen, C.B.; Heimbach, J.K.; Taner, T.; Harnois, D.M.; et al. Early Cholangiocarcinoma Detection with Magnetic Resonance Imaging versus Ultrasound in Primary Sclerosing Cholangitis. Hepatology 2021, 73, 1868–1881.
- Chazouilleres, O.; Beuers, U.; Bergquist, A.; Karlsen, T.H.; Levy, C.; Samyn, M.; Schramm, C.; Trauner, M. EASL Clinical Practice Guidelines on Sclerosing Cholangitis. J. Hepatol. 2022, 77, 761–806.
- Bowlus, C.L.; Arrivé, L.; Bergquist, A.; Deneau, M.; Forman, L.; Ilyas, S.I.; Lunsford, K.E.; Martinez, M.; Sapisochin, G.; Shroff, R.; et al. AASLD Practice Guidance on Primary Sclerosing Cholangitis and Cholangiocarcinoma. Hepatology 2023, 77, 659–702.
- Nicoletti, A.; Maurice, J.B.; Thorburn, D. Guideline Review: British Society of Gastroenterology/UK-PSC Guidelines for the Diagnosis and Management of Primary Sclerosing Cholangitis. Frontline Gastroenterol. 2021, 12, 62–66.
- Villard, C.; Friis-Liby, I.; Rorsman, F.; Said, K.; Warnqvist, A.; Cornillet, M.; Kechagias, S.; Nyhlin, N.; Werner, M.; Janczewska, I.; et al. Prospective Surveillance for Cholangiocarcinoma in Unselected Individuals with Primary Sclerosing Cholangitis. J. Hepatol. 2023, 78, 604–613.
- Aabakken, L.; Karlsen, T.H.; Albert, J.; Arvanitakis, M.; Chazouilleres, O.; Dumonceau, J.-M.; Färkkilä, M.; Fickert, P.; Hirschfield, G.M.; Laghi, A.; et al. Role of Endoscopy in Primary Sclerosing Cholangitis: European Society of Gastrointestinal Endoscopy (ESGE) and European Association for the Study of the Liver (EASL) Clinical Guideline. J. Hepatol. 2017, 66, 1265–1281.
- von Seth, E.; Arnelo, U.; Enochsson, L.; Bergquist, A. Primary Sclerosing Cholangitis Increases the Risk for Pancreatitis after Endoscopic Retrograde Cholangiopancreatography. Liver Int. 2015, 35, 254–262.
- Rupp, C.; Hippchen, T.; Bruckner, T.; Klöters-Plachky, P.; Schaible, A.; Koschny, R.; Stiehl, A.; Gotthardt, D.N.; Sauer, P. Effect of Scheduled Endoscopic Dilatation of Dominant Strictures on Outcome in Patients with Primary Sclerosing Cholangitis. Gut 2019, 68, 2170–2178.
- Bowlus, C.L.; Lim, J.K.; Lindor, K.D. AGA Clinical Practice Update on Surveillance for Hepatobiliary Cancers in Patients with Primary Sclerosing Cholangitis: Expert Review. Clin. Gastroenterol. Hepatol. 2019, 17, 2416–2422.
- Chapman, R.; Fevery, J.; Kalloo, A.; Nagorney, D.M.; Boberg, K.M.; Shneider, B.; Gores, G.J. Diagnosis and Management of Primary Sclerosing Cholangitis. Hepatology 2010, 51, 660–678.
- Taghavi, S.A.; Eshraghian, A.; Niknam, R.; Sivandzadeh, G.R.; Bagheri Lankarani, K. Diagnosis of Cholangiocarcinoma in Primary Sclerosing Cholangitis. Expert Rev. Gastroenterol. Hepatol. 2018, 12, 575–584.
- Wannhoff, A.; Brune, M.; Knierim, J.; Weiss, K.H.; Rupp, C.; Gotthardt, D.N. Longitudinal Analysis of CA19-9 Reveals Individualised Normal Range and Early Changes before Development of Biliary Tract Cancer in Patients with Primary Sclerosing Cholangitis. Aliment. Pharmacol. Ther. 2019, 49, 769–778.
- Jendrek, S.T.; Gotthardt, D.; Nitzsche, T.; Widmann, L.; Korf, T.; Michaels, M.A.; Weiss, K.-H.; Liaskou, E.; Vesterhus, M.; Karlsen, T.H.; et al. Anti-GP2 IgA Autoantibodies Are Associated with Poor Survival and Cholangiocarcinoma in Primary Sclerosing Cholangitis. Gut 2017, 66, 137–144.
- Saffioti, F.; Roccarina, D.; Vesterhus, M.; Hov, J.R.; Rosenberg, W.; Pinzani, M.; Pereira, S.P.; Boberg, K.M.; Thorburn, D. Cholangiocarcinoma Is Associated with a Raised Enhanced Liver Fibrosis Score Independent of Primary Sclerosing Cholangitis. Eur. J. Clin. Investig. 2019, 49, e13088.
- Cuenco, J.; Wehnert, N.; Blyuss, O.; Kazarian, A.; Whitwell, H.J.; Menon, U.; Dawnay, A.; Manns, M.P.; Pereira, S.P.; Timms, J.F. Identification of a Serum Biomarker Panel for the Differential Diagnosis of Cholangiocarcinoma and Primary Sclerosing Cholangitis. Oncotarget 2018, 9, 17430–17442.
- Loosen, S.H.; Roderburg, C.; Kauertz, K.L.; Pombeiro, I.; Leyh, C.; Benz, F.; Vucur, M.; Longerich, T.; Koch, A.; Braunschweig, T.; et al. Elevated Levels of Circulating Osteopontin Are Associated with a Poor Survival after Resection of Cholangiocarcinoma. J. Hepatol. 2017, 67, 749–757.
- Navaneethan, U.; Parsi, M.A.; Lourdusamy, V.; Bhatt, A.; Gutierrez, N.G.; Grove, D.; Sanaka, M.R.; Hammel, J.P.; Stevens, T.; Vargo, J.J.; et al. Volatile Organic Compounds in Bile for Early Diagnosis of Cholangiocarcinoma in Patients with Primary Sclerosing Cholangitis: A Pilot Study. Gastrointest. Endosc. 2015, 81, 943–949.e1.
- Navaneethan, U.; Parsi, M.A.; Gutierrez, N.G.; Bhatt, A.; Venkatesh, P.G.K.; Lourdusamy, D.; Grove, D.; Hammel, J.P.; Jang, S.; Sanaka, M.R.; et al. Volatile Organic Compounds in Bile Can Diagnose Malignant Biliary Strictures in the Setting of Pancreatic Cancer: A Preliminary Observation. Gastrointest. Endosc. 2014, 80, 1038–1045.
- Navaneethan, U.; Parsi, M.A.; Lourdusamy, D.; Grove, D.; Sanaka, M.R.; Hammel, J.P.; Vargo, J.J.; Dweik, R.A. Volatile Organic Compounds in Urine for Noninvasive Diagnosis of Malignant Biliary Strictures: A Pilot Study. Dig. Dis. Sci. 2015, 60, 2150–2157.
- Ishikawa, A.; Sasaki, M.; Sato, Y.; Ohira, S.; Chen, M.-F.; Huang, S.-F.; Oda, K.; Nimura, Y.; Nakanuma, Y. Frequent P16ink4a Inactivation Is an Early and Frequent Event of Intraductal Papillary Neoplasm of the Liver Arising in Hepatolithiasis. Human. Pathol. 2004, 35, 1505–1514.
- Nakamoto, S.; Kumamoto, Y.; Igarashi, K.; Fujiyama, Y.; Nishizawa, N.; Ei, S.; Tajima, H.; Kaizu, T.; Watanabe, M.; Yamashita, K. Methylated Promoter DNA of CDO1 Gene and Preoperative Serum CA19-9 Are Prognostic Biomarkers in Primary Extrahepatic Cholangiocarcinoma. PLoS ONE 2018, 13, e0205864.
- Amornpisutt, R.; Proungvitaya, S.; Jearanaikoon, P.; Limpaiboon, T. DNA Methylation Level of OPCML and SFRP1: A Potential Diagnostic Biomarker of Cholangiocarcinoma. Tumor Biol. 2015, 36, 4973–4978.
- Kim, B.; Cho, N.-Y.; Shin, S.H.; Kwon, H.-J.; Jang, J.J.; Kang, G.H. CpG Island Hypermethylation and Repetitive DNA Hypomethylation in Premalignant Lesion of Extrahepatic Cholangiocarcinoma. Virchows Arch. 2009, 455, 343–351.
- Andresen, K.; Boberg, K.M.; Vedeld, H.M.; Honne, H.; Jebsen, P.; Hektoen, M.; Wadsworth, C.A.; Clausen, O.P.; Lundin, K.E.A.; Paulsen, V.; et al. Four DNA Methylation Biomarkers in Biliary Brush Samples Accurately Identify the Presence of Cholangiocarcinoma. Hepatology 2015, 61, 1651–1659.
- Vedeld, H.M.; Grimsrud, M.M.; Andresen, K.; Pharo, H.D.; von Seth, E.; Karlsen, T.H.; Honne, H.; Paulsen, V.; Färkkilä, M.A.; Bergquist, A.; et al. Early and Accurate Detection of Cholangiocarcinoma in Patients with Primary Sclerosing Cholangitis by Methylation Markers in Bile. Hepatology 2022, 75, 59–73.
- Klump, B.; Hsieh, C.-J.; Dette, S.; Holzmann, K.; Kiebetalich, R.; Jung, M.; Sinn, U.; Ortner, M.; Porschen, R.; Gregor, M. Promoter Methylation of INK4a/ARF as Detected in Bile-Significance for the Differential Diagnosis in Biliary Disease. Clin. Cancer Res. 2003, 9, 1773–1778.
- Shin, S.-H.; Lee, K.; Kim, B.-H.; Cho, N.-Y.; Jang, J.-Y.; Kim, Y.-T.; Kim, D.; Jang, J.J.; Kang, G.H. Bile-Based Detection of Extrahepatic Cholangiocarcinoma with Quantitative DNA Methylation Markers and Its High Sensitivity. J. Mol. Diagn. 2012, 14, 256–263.
- Sriraksa, R.; Zeller, C.; El-Bahrawy, M.A.; Dai, W.; Daduang, J.; Jearanaikoon, P.; Chau-in, S.; Brown, R.; Limpaiboon, T. CpG-Island Methylation Study of Liver Fluke-Related Cholangiocarcinoma. Br. J. Cancer 2011, 104, 1313–1318.
- Wasenang, W.; Chaiyarit, P.; Proungvitaya, S.; Limpaiboon, T. Serum Cell-Free DNA Methylation of OPCML and HOXD9 as a Biomarker That May Aid in Differential Diagnosis between Cholangiocarcinoma and Other Biliary Diseases. Clin. Epigenet. 2019, 11, 39.
- Branchi, V.; Schaefer, P.; Semaan, A.; Kania, A.; Lingohr, P.; Kalff, J.C.; Schäfer, N.; Kristiansen, G.; Dietrich, D.; Matthaei, H. Promoter Hypermethylation of SHOX2 and SEPT9 Is a Potential Biomarker for Minimally Invasive Diagnosis in Adenocarcinomas of the Biliary Tract. Clin. Epigenet. 2016, 8, 133.
- Letelier, P.; Riquelme, I.; Hernández, A.; Guzmán, N.; Farías, J.; Roa, J. Circulating MicroRNAs as Biomarkers in Biliary Tract Cancers. Int. J. Mol. Sci. 2016, 17, 791.
- Marin, J.J.G.; Bujanda, L.; Banales, J.M. MicroRNAs and Cholestatic Liver Diseases. Curr. Opin. Gastroenterol. 2014, 30, 303–309.
- Li, L.; Masica, D.; Ishida, M.; Tomuleasa, C.; Umegaki, S.; Kalloo, A.N.; Georgiades, C.; Singh, V.K.; Khashab, M.; Amateau, S.; et al. Human Bile Contains MicroRNA-Laden Extracellular Vesicles That Can Be Used for Cholangiocarcinoma Diagnosis. Hepatology 2014, 60, 896–907.
- Bernuzzi, F.; Marabita, F.; Lleo, A.; Carbone, M.; Mirolo, M.; Marzioni, M.; Alpini, G.; Alvaro, D.; Boberg, K.M.; Locati, M.; et al. Serum microRNAs as Novel Biomarkers for Primary Sclerosing Cholangitis and Cholangiocarcinoma. Clin. Exp. Immunol. 2016, 185, 61–71.
- Loosen, S.H.; Lurje, G.; Wiltberger, G.; Vucur, M.; Koch, A.; Kather, J.N.; Paffenholz, P.; Tacke, F.; Ulmer, F.T.; Trautwein, C.; et al. Serum Levels of miR-29, miR-122, miR-155 and miR-192 Are Elevated in Patients with Cholangiocarcinoma. PLoS ONE 2019, 14, e0210944.
- Correa-Gallego, C.; Maddalo, D.; Doussot, A.; Kemeny, N.; Kingham, T.P.; Allen, P.J.; D’Angelica, M.I.; DeMatteo, R.P.; Betel, D.; Klimstra, D.; et al. Circulating Plasma Levels of MicroRNA-21 and MicroRNA-221 Are Potential Diagnostic Markers for Primary Intrahepatic Cholangiocarcinoma. PLoS ONE 2016, 11, e0163699.
- Miolo, G.; Muraro, E.; Caruso, D.; Crivellari, D.; Ash, A.; Scalone, S.; Lombardi, D.; Rizzolio, F.; Giordano, A.; Corona, G. Pharmacometabolomics Study Identifies Circulating Spermidine and Tryptophan as Potential Biomarkers Associated with the Complete Pathological Response to Trastuzumab-Paclitaxel Neoadjuvant Therapy in HER-2 Positive Breast Cancer. Oncotarget 2016, 7, 39809–39822.
- Hensley, C.T.; Faubert, B.; Yuan, Q.; Lev-Cohain, N.; Jin, E.; Kim, J.; Jiang, L.; Ko, B.; Skelton, R.; Loudat, L.; et al. Metabolic Heterogeneity in Human Lung Tumors. Cell 2016, 164, 681–694.
- Banales, J.M.; Iñarrairaegui, M.; Arbelaiz, A.; Milkiewicz, P.; Muntané, J.; Muñoz-Bellvis, L.; La Casta, A.; Gonzalez, L.M.; Arretxe, E.; Alonso, C.; et al. Serum Metabolites as Diagnostic Biomarkers for Cholangiocarcinoma, Hepatocellular Carcinoma, and Primary Sclerosing Cholangitis. Hepatology 2019, 70, 547–562.
- Metzger, J.; Negm, A.A.; Plentz, R.R.; Weismüller, T.J.; Wedemeyer, J.; Karlsen, T.H.; Dakna, M.; Mullen, W.; Mischak, H.; Manns, M.P.; et al. Urine Proteomic Analysis Differentiates Cholangiocarcinoma from Primary Sclerosing Cholangitis and Other Benign Biliary Disorders. Gut 2013, 62, 122–130.
- Lankisch, T.O.; Metzger, J.; Negm, A.A.; Voβkuhl, K.; Siwy, J.; Weismüller, T.J.; Schneider, A.S.; Thedieck, K.; Baumeister, R.; Zürbig, P.; et al. Bile Proteomic Profiles Differentiate Cholangiocarcinoma from Primary Sclerosing Cholangitis and Choledocholithiasis. Hepatology 2011, 53, 875–884.
- Voigtländer, T.; Metzger, J.; Schönemeier, B.; Jäger, M.; Mischak, H.; Manns, M.P.; Lankisch, T.O. A Combined Bile and Urine Proteomic Test for Cholangiocarcinoma Diagnosis in Patients with Biliary Strictures of Unknown Origin. United Eur. Gastroenterol. J. 2017, 5, 668–676.
- Betesh, L.; Comunale, M.A.; Wang, M.; Liang, H.; Hafner, J.; Karabudak, A.; Giama, N.H.; Moser, C.D.; Miyoshi, E.; Roberts, L.R.; et al. Identification of Fucosylated Fetuin-A as a Potential Biomarker for Cholangiocarcinoma. Prot. Clin. Appl. 2017, 11, 1600141.
- Lapitz, A.; Arbelaiz, A.; Olaizola, P.; Aranburu, A.; Bujanda, L.; Perugorria, M.J.; Banales, J.M. Extracellular Vesicles in Hepatobiliary Malignancies. Front. Immunol. 2018, 9, 2270.
- Hirsova, P.; Ibrahim, S.H.; Verma, V.K.; Morton, L.A.; Shah, V.H.; LaRusso, N.F.; Gores, G.J.; Malhi, H. Extracellular Vesicles in Liver Pathobiology: Small Particles with Big Impact. Hepatology 2016, 64, 2219–2233.
- Arbelaiz, A.; Azkargorta, M.; Krawczyk, M.; Santos-Laso, A.; Lapitz, A.; Perugorria, M.J.; Erice, O.; Gonzalez, E.; Jimenez-Agüero, R.; Lacasta, A.; et al. Serum Extracellular Vesicles Contain Protein Biomarkers for Primary Sclerosing Cholangitis and Cholangiocarcinoma. Hepatology 2017, 66, 1125–1143.
- Lapitz, A.; Arbelaiz, A.; O’Rourke, C.J.; Lavin, J.L.; Casta, A.L.; Ibarra, C.; Jimeno, J.P.; Santos-Laso, A.; Izquierdo-Sanchez, L.; Krawczyk, M.; et al. Patients with Cholangiocarcinoma Present Specific RNA Profiles in Serum and Urine Extracellular Vesicles Mirroring the Tumor Expression: Novel Liquid Biopsy Biomarkers for Disease Diagnosis. Cells 2020, 9, 721.
- Lapitz, A.; Azkargorta, M.; Milkiewicz, P.; Olaizola, P.; Zhuravleva, E.; Grimsrud, M.M.; Schramm, C.; Arbelaiz, A.; O’Rourke, C.J.; La Casta, A.; et al. Liquid Biopsy-Based Protein Biomarkers for Risk Prediction, Early Diagnosis, and Prognostication of Cholangiocarcinoma. J. Hepatol. 2023, 79, 93–108.
- Valle, J.W.; Kelley, R.K.; Nervi, B.; Oh, D.-Y.; Zhu, A.X. Biliary Tract Cancer. Lancet 2021, 397, 428–444.
- Singhi, A.D.; Nikiforova, M.N.; Chennat, J.; Papachristou, G.I.; Khalid, A.; Rabinovitz, M.; Das, R.; Sarkaria, S.; Ayasso, M.S.; Wald, A.I.; et al. Integrating Next-Generation Sequencing to Endoscopic Retrograde Cholangiopancreatography (ERCP)-Obtained Biliary Specimens Improves the Detection and Management of Patients with Malignant Bile Duct Strictures. Gut 2020, 69, 52–61.
- Arechederra, M.; Rullán, M.; Amat, I.; Oyon, D.; Zabalza, L.; Elizalde, M.; Latasa, M.U.; Mercado, M.R.; Ruiz-Clavijo, D.; Saldaña, C.; et al. Next-Generation Sequencing of Bile Cell-Free DNA for the Early Detection of Patients with Malignant Biliary Strictures. Gut 2022, 71, 1141–1151.
More