Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 2 by Rita Xu and Version 1 by Xin Wang.
Angiosperms are frequently assumed to constitute a monophyletic group. Therefore, the origin of angiosperms is a key question in systematic botany since the answer to this question is hinged with many questions concerned with angiosperm evolution.
- origin
- angiosperms
- misconcept
- fossil
1. An Important but Perplexing Question
The term “angiosperm” was coined in 1690 by the German botanist Paul Hermann (1646–1695) from Greek angeion (vessel) + spermos (seed), to name those plants that have their seeds contained, in contrast to “gymnosperms” that have their seeds exposed. In the current world, there are over 300,000 species of angiosperms, accounting for over 90% of the plant species diversity [1]. This single group of plants provide most of the food and materials necessary for the sustainable development of humans. Without them, the occurrence of humans on Earth would be inconceivable. Needless to say, angiosperms are the single most important plant group for us. It is natural for many botanists to focus their attention on angiosperms and study various aspects of angiosperms. Among these questions concerned with angiosperms, a fundamental question in botany is the origin of angiosperms, namely, from which group the angiosperms were derived. Since angiosperms are frequently taken as a monophyletic group, how the earliest angiosperms looked is a key question in angiosperm systematics.
2. Conflicting Answers for the Same Question
A natural but unpleasant status of current studies is that, rather than reaching a consensus, several different and conflicting answers have been given to the same question. Since it is a mission impossible to give an exhaustive review of all these works in this short paper, I prefer to focus my review on questions of great influence in the past couple of centuries. “Angiosperm” is a well-defined botanical term. From the first day when Paul Herman (1646–1695) coined the term “angiosperm” in 1690, the meaning of the term “angiosperm” has been clear. Per the definition of the term angiosperm, angiosperms are distinguished from gymnosperms by their reproductive parts (flowers) that contain their seeds/ovules, which is not seen in gymnosperms. Therefore, the core question in the origin of angiosperms is how previously naked seeds/ovules became enclosed in the ancestral angiosperms that may have little morphological difference from their gymnosperm peers. As for the provenance of angiosperm flowers, there used to be a confrontation between the Pseudanthial Theory and the Euanthial Theory (also known as the Anthostrobilus Theory). The Euanthial Theory thought that angiosperms were derived from Bennettitales-related groups, unisexual flowers were derived from former bisporangiate strobili, and apocarpous Magnoliales were the most ancestral group among extant angiosperms [2]. While the Pseudanthial Theory thought that angiosperms were derived from Gnetales, and amentiferous-hamamelid orders Casuarinales, Fagales, Myricales, Juglandales, Chloranthaceae, and Piperaceae represented the most ancestral groups in extant angiosperms [3]. These two schools were stalemated around 1900, and the balance between them was historically broken by a slight impetus: the occurrence of fossil evidence of bisexual reproductive organs in Bennettitales [4,5,6][4][5][6]. Although current morphological knowledge of Bennettitales has rejected comparability and phylogenetic relationship between Bennettitales and Magnoliales, bisexuality and spatial deployment of male and female parts in reproductive organs in both groups did break the delicate balance between two confronting schools: Arber and Parkin [2] declared the victory of Euanthial Theory over Pseudanthial Theory. Thereafter Pseudanthial Theory permanently lost its voice in botany, although both theories have now been discarded due to a lack of favoring fossil evidence and emerging molecular systematics. The current mainstream angiosperm system (APG system), however, was more or less born out of and at least initially calibrated by the Euanthial Theory.3. Reasons Underlying the Controversies
In the debates on the nature of carpels, botanists in both schools have been working hard to convince the other with their own evidence, but neither succeeded. Why? Because both schools have found and shown evidence favoring their own ideas in extant plants that are all from the same time plane. The aims of these scholars were to reconstruct the deep history before extant plants, but the great diversity of extant angiosperms does not disfavor any hypotheses, just as the infinite number of straight lines go across a single point. Without time information embodied only in fossils, cherry-picking favoring evidence among diverse gynoecia and carpels in extant angiosperms is an easy job and allows any saying to go. Focusing on a favoring feature of selected groups allows one easily to be blind to unpleasant evidence and alternatives from the others. With valuable time information that is lacking in extant plants, plant fossils provide crucial independent data testing and sifting a valid hypothesis out of many proposed candidates.4. False Evidence Favoring the Euanthial Theory
The dominance of the Euanthial Theory is not a random aftermath. Many publications favored this theory, and these publications (especially those by famous authors) persuaded many otherwise hesitant botanists and consolidated the foundation for the theory. The Euanthial Theory started its dominance in the war against the Pseudanthial Theory when Arber and Parkin published their paper titled “On the origin of angiosperms” in 1907, taking advantage of then just published bisporangiate strobili of Bennettitales [6]. The bisexuality and proximal arrangement of androecium and gynoecium in Bennettitales were compared to those in Magnoliales and interpreted as evidence favoring the Euanthial Theory. This conclusion was further repeatedly strengthened by several ensuing publications of influence published in the past century, including anatomic work of Canright [9][7], studies of fossil early angiosperm Archaeanthus by Dilcher and Crane [10][8], Archaefructus by Sun et al. [11][9], and Monetianthus by Friis et al. [12][10]). These works initially were interpreted as favoring the Euanthial Theory. However, more detailed examinations indicate that these favorings are all spurious (see below).5. Emerging Inconsistencies and Lacunae in Proof
If all evidence favored the Euanthial Theory, then any new angiosperm system, including the APG system, would not occur. As time passed, increasing lacunae in the proof of Euanthial Theory popped up. The earliest and alarming one is from Parkin (one of the only two authors of Arber and Parkin [2]) himself: Parkin admitted in 1925 (Arber passed away in 1918 [13][11]) that their conclusion made eighteen years before had no fossil evidence support [14][12]. In the fossil record, at least as early as 1984 some negative evidence started to emerge. Friis [15][13] reported over 100 angiosperm fossil taxa from the Late Cretaceous of Sweden. It is noteworthy that, despite the abundant and diverse taxa reported in the fossil flora, no trace of Magnoliaceae was found. This lack of Magnoliaceae in the fossil flora was quite conspicuous and alarming, especially when the Euanthial Theory was still of great influence. However, both of the above important alarming signals were either downplayed or largely ignored, (intentionally?) successfully. One cannot help asking, “Are these publications peerless, singular exceptional exceptions? Should they be reasonably ignored or downplayed, considering more other publications favoring the Euanthial Theory?” Reviewing these “publications favoring the Euanthial Theory” indicates that at least some of the other favoring-appearing publications have been involved in various forms of data misinterpretation or even falsification, and their corresponding supposed favor for the Euanthial Theory should either be discarded, discounted, or at least re-evaluated. According to the Euanthial Theory, the ovules in Magnoliaceae should be borne on the ventral margin of carpels, as ovules are supposed to be borne on the margins of megasporophylls. This expectation was met by Canright using his anatomic work: Canright [9][7] provided some specious anatomic evidence favoring the theory, based on his anatomic study of magnoliaceous fruits/carpels. Indeed, he appeared to have succeeded, as no one (at least the reviewers of his paper did not) realized that Canright had altered the data about the connection of ovule-bearing vascular bundle to meet the theoretical expectation of the Euanthial Theory. In the only picture demonstrating ovule-bearing vascular bundle provenance in Magnolia salicifolia Maxim. (his Figure 8), the ovule was connected to the “cort. b.” (cortical bundle), but the same ovule was drawn to be connected to the “v. b.” (ventral bundle) in his Figure 17 [9][7]. It appeared that his subtle change escaped the attention of the reviewers, who probably also expected the situation in his Figure 17. This case is not peerless. Instead similar cases also occurred in some studies of fossil angiosperms: Dilcher and Crane [10][8] drew ovules of Archaeanthus on the ventral margins of carpels in their Figure 60h (just as the Euanthial Theory expected), their own Figure 24 indicated that at least some of the ovules were borne on the dorsal fruit margin [10][8]. In addition, the famous early angiosperm Archaefructus was initially described as bearing “follicles”, implying that ovules were borne on the ventral margin of carpels in Archaefructus [11][9]. This “rational” implication congruous with the Euanthial Theory was later found groundless as the ovules were later found borne along the dorsal margin of the carpels/fruits [16][14]. In both of the above two cases, how these “correct” and “rational” mistakes had escaped the supposed careful examinations of the reviewers is a mystery. A rational guestimation is that the presented information was compatible with pre-existing one, which has long been imbibed in the mind of reviewers beforehand. In addition, the almost universally expected ventral position of ovules in carpels has been disfavored by Endress’Brasenia ovule borne on the dorsal (rather than ventral) vascular bundle [17][15].6. “Sporophyll” and the Euanthial Theory
“Sporophyll” is a frequently and widely used term for seed plants in botany textbooks. It has a far-reaching influence on almost all botanists. To many, reproductive organs of seed plants cannot be themselves without “sporophylls”. All sporangioids (sporangia, pollen sacs, and ovules) in ferns, gymnosperms, or angiosperms were interpreted as borne on “sporophylls” in currently available botany textbooks. “Sporophyll” is one of the few founding concepts underlying the Euanthial Theory. In the Euanthial Theory, a carpel was interpreted as a “sporophyll” bearing ovules along its margins and enclosing the ovules through enrolling along its longitudinal axis. “Sporophyll” was so popular in botany for reasons. According to Arber and Parkin [2], they interpreted a carpel as derived from a leaf, at least partially, because of Goethe’s dictum “All is leaf”. Goethe (an amateur botanist [20][16]) is not the inventor of this idea. Far before Goethe, botanists Grew and Wolff had made similar statements in 1672 and 1768, respectively [21][17]. The influence of these botanists appeared little in botany. Celebrity and appealing expression won over professionalism, even among the assumed scientific professional botanical circle: Goethe had more heavily influenced Arber and Parkin, who won the debate against the Pseudanthial Theory and initiated the dominance of the Euanthial Theory while citing the dictum of celebrity Goethe [2]. The validity of Goethe’s statement was recently challenged by evidence from several lines. First, the most leaf-like female reproductive parts, “megasporophylls”, in all living plants, of Cycas were proven, by a developmental experiment, to be leaf-like due to mechanical pressure but not genes [22][18]. Second, vascular bundles in at least some cycad “megasporophylls” furcate in three dimensions, rather than in a single plane as expected for a typical leaf [23,24][19][20]. Third, in the earliest cycad fossil “megasporophyll”, of Permian Primocycas [25][21], the ovules were not borne along the supposed leaf margins. Fourth, the currently available fossil evidence [26,27][22][23] indicates that all sporangia in all early land plants are borne on the termini of branches rather than on any leaf margins. Since ovules and pollen sacs are all derived from these terminal sporangia, the supposed marginal position of ovules, if indeed so, must be a consequence of long-time evolution rather than an ancestral status in seed plants. Fifth, Crane has repeatedly stated that, at least in some Mesozoic fossil plants, the ovules are borne on the termini of branches rather than leaf margins [28,29][24][25]. Sixth, the carpels of Michelia (Magnoliaceae) have been proven synorganizations of a placenta and its subtending leaf (Figure 1), rather than metamorphs of leaves [30][26]. It appears that Goethe’s conception has misled almost all botanists over centuries [31][27], and it is time to clean it out of botany, at least that of seed plants. The term “sporophyll” is a misconception that should be discarded in the botany of seed plants, despite its wide usage and popularity. The removal of “megasporophylls” undermines the validity of the Euanthial Theory, as the former is a core concept founding the latter.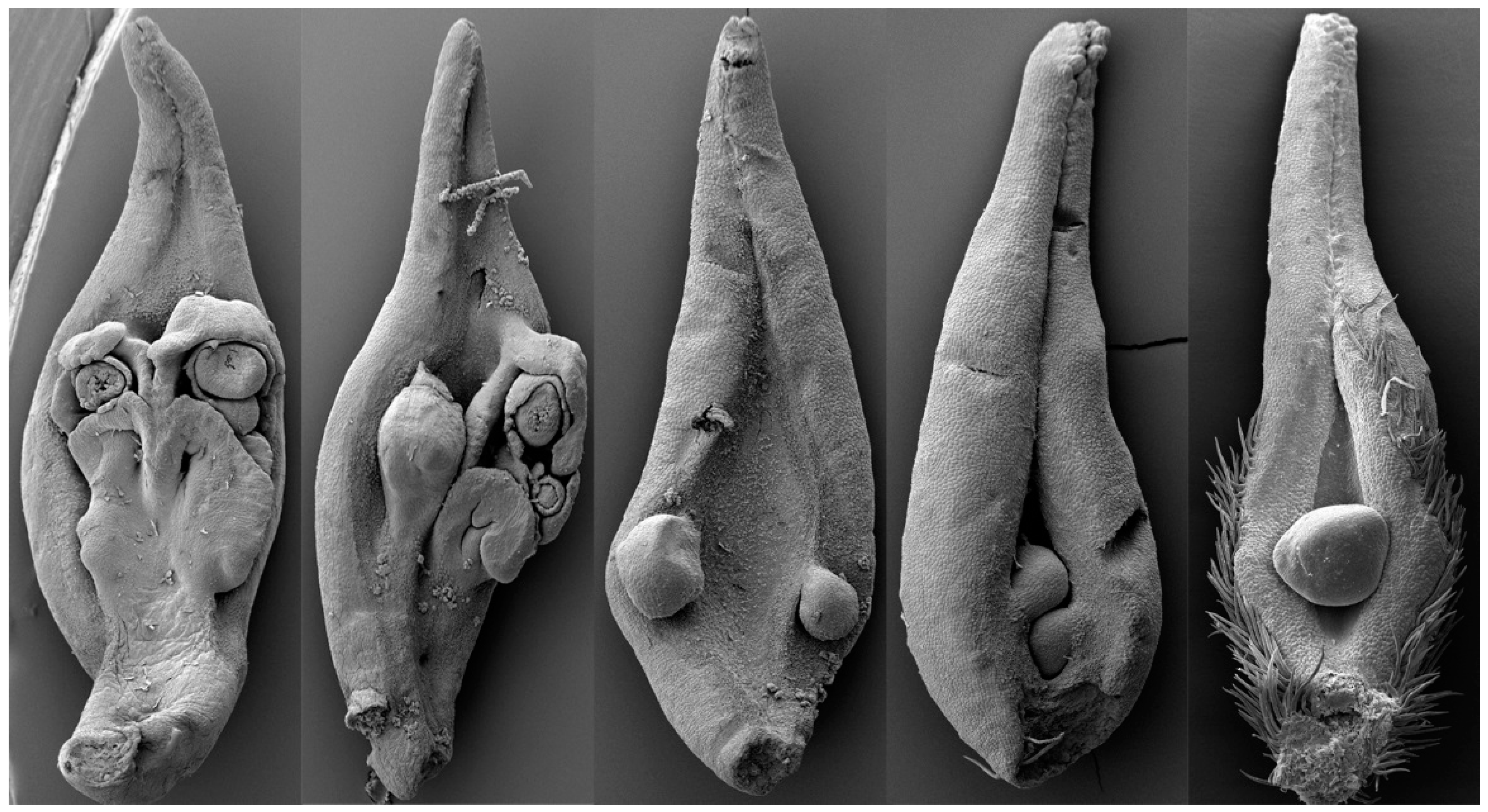
References
- Crepet, W.L.; Niklas, K.J. Darwin’s second “abominable mystery”: Why are there so many angiosperm species? Am. J. Bot. 2009, 96, 366–381.
- Arber, E.A.N.; Parkin, J. On the origin of angiosperms. J. Linn. Soc. Lond. Bot. 1907, 38, 29–80.
- von Wettstein, R.R. Handbuck der Systematischen Botanik. II. Band; Franz Deuticke: Leipzig, Germany; Wien, Austria, 1907; Available online: https://books.google.com.sg/books?hl=en&lr=&id=4nwaAAAAYAAJ&oi=fnd&pg=PA1&dq=Handbuch+der+Systematischen+Botanik&ots=1lIjHfxprZ&sig=JCaCwpEnkSCR2LmGgRtpoOX7KZI#v=onepage&q=Handbuch%20der%20Systematischen%20Botanik&f=false (accessed on 1 October 2023).
- Wieland, G.R. A study of some American fossil cycads. Parts III. The female fructification of Cycadeoidea. Am. J. Sci. 1899, 7, 383–391.
- Wieland, G.R. A study of some American fossil cycads. Part IV. On the microsporangiate fructifications of Cycadeoidea. Am. J. Sci. 1901, 11, 423–436.
- Wieland, G.R. American Fossil Cycads; The Wilkens Sheiry Printing Co.: Washington, DC, USA, 1906; Volume 34, p. 295.
- Canright, J.E. The comparative morphology and relationships of the Magnoliaceae. III. Carpels. Am. J. Bot. 1960, 47, 145–155.
- Dilcher, D.L.; Crane, P.R. Archaeanthus: An early angiosperm from the Cenomanian of the Western Interior of North America. Ann. Mo. Bot. Gard. 1984, 71, 351–383.
- Sun, G.; Dilcher, D.L.; Zheng, S.; Zhou, Z. In search of the first flower: A Jurassic angiosperm, Archaefructus, from Northeast China. Science 1998, 282, 1692–1695.
- Friis, E.M.; Pedersen, K.R.; von Balthazar, M.; Grimm, G.W.; Crane, P.R. Monetianthus mirus gen. et sp. nov., a nymphaealean flower from the Early Cretaceous of Portugal. Int. J. Plant Sci. 2009, 170, 1086–1101.
- Seward, A.C. Dr.E.A. Newell Arber. Nature 1918, 101, 328–329.
- Parkin, J. The phylogenetic classification of flowering plants. Nature 1925, 115, 385–387.
- Friis, E.M. Preliminary report of Upper Cretaceous angiosperm reproductive organs from Sweden and their level of organization. Ann. Mo. Bot. Gard. 1984, 71, 403–418.
- Ji, Q.; Li, H.; Bowe, M.; Liu, Y.; Taylor, D.W. Early Cretaceous Archaefructus eoflora sp. nov. with bisexual flowers from Beipiao, Western Liaoning, China. Acta Geol. Sin. (Engl. Ed.) 2004, 78, 883–896.
- Endress, P.K. Carpels of Brasenia (Cabombaceae) are completely ascidiate despite a long stigmatic crest. Ann. Bot. 2005, 96, 209–215.
- Arber, A. The Natural Philosophy of Plant Form; University Press: Cambridge, UK, 1950.
- Arber, A. Introduction to Goethe’s botany. Chron. Bot. 1946, 10, 63–87.
- Wang, X.; Luo, B. Mechanical pressure, not genes, makes ovulate parts leaf-like in Cycas. Am. J. Plant Sci. 2013, 4, 53–57.
- Stevenson, D.W. Morphology and systematics of the Cycadales. Mem. N. Y. Bot. Gard. 1990, 57, 8–55.
- Hu, Z.; Liu, W.; Cai, X.; Wu, P.; Ning, X. Plant Anatomy; Higher Education Press: Beijing, China, 2010; p. 355.
- Zhu, J.N.; Du, X.M. A new cycad—Primocycas chinensis gen. et sp. nov. discovered from the Lower Permian in Shanxi, China and its significance. Acta Bot. Sin. 1981, 23, 401–404.
- Edwards, D. Embryophytic sporophytes in the Rhynie and Windyeld cherts. Trans. R. Soc. Edinb. Earth Sci. 2003, 94, 397–410.
- Hao, S.; Xue, J. The Early Devonian Posongchong Flora of Yunnan; Science Press: Beijing, China, 2013; p. 366.
- Shi, G.; Leslie, A.B.; Herendeen, P.S.; Herrera, F.; Ichinnorov, N.; Takahashi, M.; Knopf, P.; Crane, P.R. Early Cretaceous Umkomasia from Mongolia: Implications for homology of corystosperm cupules. New Phytol. 2016, 210, 1418–1429.
- Crane, P.R.; Herendeen, P.S.; Herrera, F.; Shi, G. Diversity and homologies of corystosperm seed-bearing structures from the Early Cretaceous of Mongolia and China. In Proceedings of the 10th European Palaeobotany and Palynology Conference, Dublin, Ireland, 12–19 August 2018; p. 88.
- Zhang, X.; Liu, W.; Wang, X. How the ovules get enclosed in magnoliaceous carpels. PLoS ONE 2017, 12, e0174955.
- Wang, X.; Liu, Z.-J.; Liu, W.; Liao, W.; Zhang, X.; Liu, Z.; Hu, G.; Guo, X.; Wang, Y. Stepping out of the shadow of Goethe: For a more scientific plant systematics. Chin. Bull. Bot. 2020, 55, 505–512.
More