Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Sumit Rathod and Version 2 by Jason Zhu.
Neuroinflammation is a complex biological process that typically originates as a protective response in the brain. This inflammatory process is triggered by the release of pro-inflammatory substances like cytokines, prostaglandins, and reactive oxygen and nitrogen species from stimulated endothelial and glial cells, including those with pro-inflammatory functions, in the outer regions. While neuronal inflammation is common in various central nervous system disorders, the specific inflammatory pathways linked with different immune-mediated cell types and the various factors influencing the blood-brain barrier significantly contribute to disease-specific characteristics.
- neuroinflammation
- proinflammatory cytokines
- endocannabinoid system
- microglia
1. Introduction
Over the course of history, cannabis has been used to serve recreational and therapeutic uses. New findings have provided light on the effects of cannabis and its constituents (cannabinoids) on the brain and other regions of the CNS. These psychoactive substances have shown potential in the treatment of neurodegenerative and mental disorders associated with neural inflammation, neurodegeneration, and pain [1][62]. Naturally occurring cannabinoids derived from plants, produced in a laboratory, their target receptors, and the enzymes responsible for their generation and degradation comprise the cannabinoid system [2][63]. Modifications in the cannabis system are connected with variations in concurrent effects and are linked to the inflammatory manifestations of these diseases [3][64]. Non-neuronal cells within the CNS have a particular connection with the onset, perseverance, and alleviation of neurological inflammation, neurological damage, and pain conditions [1][4][5][62,65,66]. This in-depth examination focuses on the locations of receptors that recognize cannabinoids and the ECS in different parts of the brain and the CNS, as well as their potential involvement in neurological modulation [6][67].
The prior genomic and pharmacological study has suggested that there are two distinct kinds of GPCR (CB1 and CB2) that are broadly dispersed throughout the human body. CB1A, an alternative variation of the CB1 receptor, has been effectively found in the brain (Figure 1) [7][68]. This CB1A variation has all of the CB1 receptor’s characteristics but with a somewhat reduced effect [8][69]. However, the practical importance of this splice variation is unknown due to a lack of supporting data for its self-expression, as other groups of investigators have noticed. Although there is no prior proof for CB2 receptor binding, protein presence, or mRNA expression in the brain [9][70], certain functional findings do imply the existence of CB2 receptors in cultured cerebellar granular cells with their involvement in ERK pathways, which will lead to increasing the rate of transcription of mRNA in the granular cells [10][71]. The science of cannabis pharmacology has advanced significantly, culminating in the creation of a variety of specific agonists and antagonists suited for these different receptor subtypes [6][11][67,72].
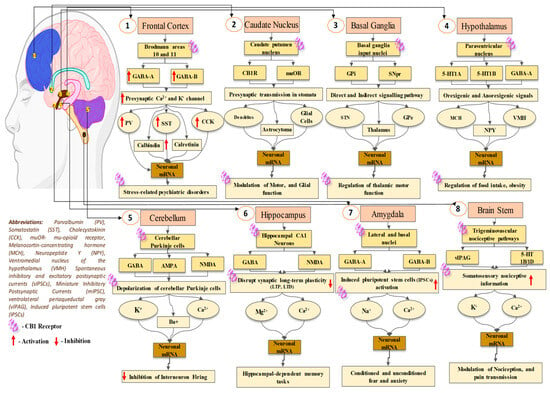
Figure 1.
Location of CB1 receptor in the brain, and its physiological retorts.
2. Establishment of the Cannabinoid Receptor 1 (CB1) in Neuronal Tissues
CB1 receptors have a distinct CNS distribution and are found at greater levels in the mammalian brain compared to other recognized GPCRs. CB1 receptor distribution trends have mostly been examined using radioligand binding and autoradiography, as well as immunohistochemistry approaches. Among the most extensively utilized tritiated cannabinoid receptor ligands in affinity trials or for autoradiography are the CB1-selective [3H]SR141716A, [3H]CP55940, [3H]WIN55212-2, and [3H]HU-243. The last three agonists interact with CB1 and CB2 receptors equally strongly. Cannabinoid receptor abundance in the brain rivals that of glutamate [12][73] and GABA [13][74]. Cannabinoid receptor levels throughout certain areas are equivalent to dopamine receptor levels and much above neuropeptide receptor numbers. The researchers discovered the highest concentrations of binding in the globus pallidus and substantia nigra pars reticulata, as well as the molecular strata of the cerebellum and the dentate gyrus, which are located inside the hippocampus, using automated technology densitometric analyses on film autoradiographs taken from canine, guinea pig, rat, monkey, and human brain samples [14][75]. Previously reported data suggested that additional regions of the hippocampus structure, the cortex of the brain, and the striatum display significant binding.
Radioligand affinity is modest in the hypothalamus, basal ganglia, solitary tract nucleus, and spinal cord, with slight inter-species differences [14][75]. Earlier immunocytochemical assays have demonstrated that the arrangement pattern of the CB1 receptors closely matches that of cannabinoid sites of binding, as discovered by earlier radioligand experiments. According to numerous research [15][16][17][18][76,77,78,79], these receptors have significantly higher levels of activity in certain locations, including the sense of smell, hippocampus, neocortex, and brainstem. The different arrangement of CB1 receptors in these locations correlates to the regions that are responsible for the different effects of cannabinoid agonists, such as motor control, body position, hypothermia, memory and learning, hunger, vomiting, and alleviation of pain. This finding lends strong credence to the hypothesis that cannabinoids’ activities in the cerebral cortex are primarily mediated by CB1 receptors. CB1 receptor positivity is primarily associated with nerve fiber networks and axon junctions when evaluated at the cellular level, with little activity in neurons and cell nuclei. The above distribution pattern corresponds to the idea of a presynaptic mode of activity.
The positivity of axonal CB1 receptors has been associated with presynaptic nerve terminals of GABAergic neurons, notably ones that are part of cholecystokinin-containing basket cells, in the thoroughly investigated rat hippocampus [19][20][21][80,81,82]. Although there is little proof [15][76] as well as contradictory views [16][19][22][77,80,83] regarding the existence of CB1 receptors on glutamatergic cells, in situ hybridization investigations suggest that they exist, although in far lower amounts than GABAergic neurons [23][84]. Antibodies addressing extrinsic regions of CB1 receptors were used to assess their distribution on cell surfaces, allowing the analysis of their outermost expression inside living cells. Additionally, the immunolabeling of the surface of cells’ CB1 receptors in hippocampal nerve cells is particularly numerous at GABAergic synaptic terminals, but no labeling of glutamatergic neurons as a whole is observed [24][85] (Figure 1).
3. Cannabinoid Receptor 1 (CB1) in the Communication of Neuronal Signaling Pathways
The reduction in cyclic AMP (cAMP) synthesis induced by agonists [25][86] and the inhibition of Ca2+ inward flows mediated by N- and P/Q-type Ca2+ channels [26][87] are two often documented cytoplasmic effects of CB1 receptor activation in neural networks. These characteristics are similar to those seen in other GPCRs that govern synaptic information transmission (like mGlu, muscarinic, and opioid receptors). Furthermore, these Ca2+ channels are mostly found strategically and are required to release neurotransmitters in anticipation of inputs. Cannabis-based compounds also have electrophysiology impacts, such as activating A-type inwardly conducting gates [27][88] and inhibiting M-type K+ channels [28][89]. Specific effects on K+ channels may be caused by variations in cAMP levels [25][27][86,88]. Pre-conditioning cells with pertussis toxin inhibit the effect of cannabis on ion channels, showing the presence of Gi/o proteins. Nevertheless, pertussis toxin treatment reveals a cannabinoid receptor-triggered enhancive impact on cAMP buildup inside cultured striatal neuronal cells, revealing the commencement of a new signaling cascade incorporating a Gs protein [29][90].
Cannabinoids stimulate intrinsic signaling networks in the CNS, including the mitogen-activated protein (MAP) kinase pathway [30][91], and maybe the c-Jun N-terminal kinase via a phosphoinositide 3’-kinase-dependent process [31][92]. In addition, earlier research has shown that CB1 receptors can interact with phospholipase C in cerebellar granule cells, causing intracellular calcium to be released [32][93]. The CB1 receptor, like many other GPCRs, has a reduced response after being exposed to agonists, a condition known as agonist-induced desensitization [33][34][35][36][94,95,96,97]. This method involves G-protein disengagement after modification by G-protein-coupled receptor kinases (GRKs), resulting in receptor internalization [35][96]. The phosphorylation of receptors by GRKs causes beta-arrestin binding, which isolates receptors from heterotrimeric G-proteins and drives CB1 receptors into clathrin-coated vesicles [34][35][95,96]. CB1 receptor internalization has been examined with transgenic Chinese hamster ovary or AtT20 cell lines [33][34][94,95], and native receptors generated in cultured neurons in hippocampal regions or F11 cells [36][97].
4. Role of Cannabinoid Receptor 1 (CB1) in the Modulation of Neuronal Physiology
Cannabinoid impacts can be divided into two categories: those influenced primarily by cannabinoid receptors and those impacted indirectly by competing mechanisms [34][95]. Cannabinoid CB1 receptor stimulus influences the release of many neurotransmitters inside the CNS, including major neurotransmitters that are excitatory and inhibitory like glutamate and GABA [37][98]. Cannabinoids also influence the discharge of slower-acting neurotransmitters and neurological modulators like opiates, acetylcholine, dopamine, and noradrenaline [38][39][40][41][42][43][99,100,101,102,103,104]. The processes that are driving these consequences at the cellular level are mostly linked to presynaptic Ca2+ and K+ channel regulation. Cannabinoids modulate GABA transmission in various CNS areas, notably the cortex, the basal ganglia, the cerebellum, and the hypothalamus. However, research has demonstrated that ∆9-THC has an inhibitory impact on GABA absorption in the cerebral cortex and basal ganglia [44][45][105,106], and electrophysiology investigations on GABAergic transmission through synapses do not support this influence.
Morphological studies indicate a close connection between high amounts of CB1 receptor immunoreactivity and mRNA and GABAergic nerve cells [16][21][46][77,82,107]. Several neurochemical research [21][82] and electrophysiology testing [20][47][48][81,108,109] in the hippocampus have demonstrated cannabis’ ability to modulate GABA input from a particular group of inhibitory neurons. Previously reported studies explored that cannabis-based compounds inhibit GABA emission via a presynaptic process in the regions of basal ganglia [49][50][51][52][53][110,111,112,113,114], brainstem [54][55][115,116], and cerebellum [56][57][117,118]. The bulk of data suggest that cannabis regulates GABA distribution mostly through blocking synaptic Ca2+ channels (particularly N or N/PQ channels) [40][48][101,109], while in other cases, they may directly impact the releasing pathway [24][40][55][85,101,116]. Cannabinoids additionally affect glutamate transport via many crucial mechanisms. Previous investigations have shown that CB1 receptors were present in excitatory synapses of cultured cerebellar granule cells [58][119].
Cannabinoid receptor activation inhibits glutamate release at the junction among Purkinje cells and longitudinal fibers in cerebellar slices, according to substantial electrophysiological data [59][60][61][120,121,122]. Also, CB1 receptor stimulation inhibits glutamate release via the cerebral nerve in the striatum [62][123]. In addition, earlier reported studies explored that cannabinoids decrease the glutamatergic transmission of synapses in the middle brain region specifically from periaqueductal grey [55][116]. Studies also suggested that cannabis-based compounds could impact the discharge of glutamate from a glutamatergic cerebral afferent nerve in the nucleus accumbens, incorporating terminal K+ channels [63][124]. There is uncertainty about excitatory synaptic communication in the hippocampus, with some studies indicating inhibition by cannabis [64][65][125,126] and others indicating no impact [44][66][105,127] (Figure 2).
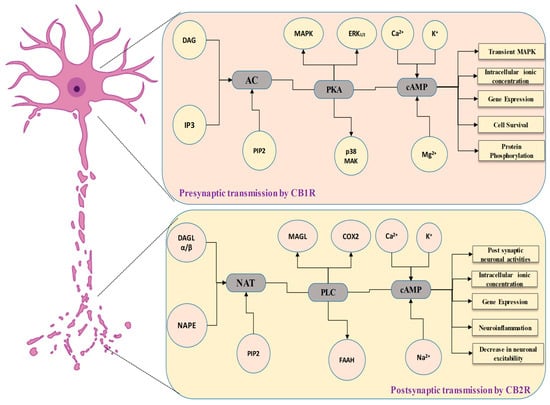
Figure 2.
Synaptic transmission by CB1 and CB2 receptors in the neurons, and its physiological retorts.
5. The Involvement of the Cannabinoid Receptor 1 (CB1) in Synaptic Modulation
Cannabinoids impede multiple types of plasticity of synapses in the hippocampus, including long-term potentiation (LTP) [65][67][68][126,128,129] and long-term depression (LTD) [65][126], via a CB1 receptor-dependent process. Studies also evidence that cannabinoids inhibit synaptic plasticity in the brain [69][130] and striatum [70][131]. The particular mechanism beyond such impacts is yet unknown. Previous data also suggested that cannabinoids modulate hippocampus neural plasticity by decreasing the release of glutamate [65][126]. This blockage can be overcome as the neuronal development of NMDA receptors rises, as demonstrated in Mg2+-depleted circumstances, or in cases where the postsynaptic cell membrane depolarizes via LTP or LTD induction methods [66][127].
Because cannabis influences just a specific population of interneurons, the delicate relationship between synaptic plasticity and its effect in inhibiting the transmission of synaptic information is expected to be subtle. This nuance might explain why, contrary to the expected result of a general decrease in inhibitory signaling, cannabis does not increase the plasticity of synapses. The effect of cannabis in inhibiting synaptic communication has the potential to impact the intensity of network oscillations, which are important for activities like memory and learning [20][81]. According to studies conducted on mice with deficient CB1 receptors, endocannabinoids have a consistent inhibitory effect on synaptic remodeling. This suggests that anandamide may work as an intrinsic messenger, controlling cognitive-linked activities.
6. Cannabinoid Receptor 1 (CB1) Gene Expression in the Brain
CB1 receptors are the possible therapeutic targets in a variety of illnesses. They nevertheless contribute to the euphoric properties of marijuana, as well as weakening motor abilities and cognition. Such features limit its therapeutic application, prompting a further investigation to uncover the cellular mechanisms driving such cannabinoid-associated behaviors [71][132]. Understanding these pathways is critical for developing measures to mitigate such unfavorable effects. Adaptation to the recurrent treatment of cannabis agonists is being linked to CB1 receptor desensitization and decreased levels (receptor ablation). Differences in CB1 receptor desensitization and decreased levels have been discovered in rats exposed to THC or synthetic cannabinoids [72][133]. Identical regional differences in CB1 receptor-reduced levels were identified in the human cerebral cortex [73][74][134,135].
Additionally, investigations also demonstrated the presence of CB1 receptors across the brain regions, with significant levels in the hippocampus, cerebral cortex, and cerebellar molecular layers. The hypothalamus, periaqueductal grey, and basolateral amygdala have fewer degrees of expression. The aforementioned distribution pattern corresponds to the cannabinoids’ immediate consequences, including antinociception, catalepsy, hyperlocomotion, hypothermia, and cognitive impairment [75][76][136,137]. A line of research has revealed that transcription factors that may be triggered under particular situations naturally exist in the brain and exhibit species-specific changes in activity. Although there has not been an immediate comparison of basal CB1 receptors expression, a more indirect assessment may be conducted, utilizing the BrainStars (B*) database, which comprises DNA microarray information collected from the brain of a mouse [77][138].
In addition, reverse transcription polymerase chain reaction (RT-PCR) has been employed to evaluate the expression of messenger RNA (mRNA) that encodes the cannabinoid CB1 receptor. The investigation included the post-mortem brain tissue from people who did not have neurological conditions as well as persons who died from Parkinson’s disease or related conditions [75][136]. CB1 receptor mRNA was found in the striatum as well as extra striatal areas (such as the area of the globus pallidus and substantia nigra). CB1 receptor mRNA levels were shown to be lower in affected neurons in the caudate nucleus, anterior dorsal putamen, and external section of the globus pallidus. However, no significant changes in CB1 receptor mRNA levels were noted in the other examined brain regions. The results suggest that alterations in CB1 receptor mRNA expression developed in Parkinson’s disease; nevertheless, the impact of treatment cannot be ignored. Furthermore, these findings show a relationship between changes in dopaminergic pathways and changes in CB1 receptor mRNA activity [77][138].
7. Establishment of the Cannabinoid Receptor 2 (CB2) in Neuronal Tissues
According to investigations in the field of neurological conditions, CB2 principally works in immunological regulation. Brain tissue data from disorders such as Alzheimer’s, Multiple Sclerosis, and ALS show a distinct and particular existence of CB2 in microglial cells [78][139]. A separate investigation found that CB2 inhibits the production of pro-inflammatory cytokines via stimulated microglia in Alzheimer’s disease [79][140]. In addition, CB2 stimulation, like CB1, promotes the creation of new synapses in adults, and there is accumulating proof suggesting it may be involved in modulating the permeation of the BBB [80][81][141,142]. CB2 receptors were considered to be lacking in the brain in the early stages due to the failure to identify CB2 mRNA transcripts using different methods [82][83][84][85][143,144,145,146]. These observations contributed to the designation of CB2 receptors as “peripheral” cannabinoid receptors [86][87][147,148]. Nevertheless, the theory has been challenged by the discovery of CB2 receptors across the CNS [82][86][143,147]. CB2 receptors in the brain differ from CB1 receptors in that they express themselves at a lesser degree than CB1 receptors, hinting that CB2 receptors may not be linked to cannabis-related effects in normal physiological circumstances.
Because CB2 receptors activity in the brain is very susceptible to induction, there is a fast rise in CB2 receptors activity inside the brain in some clinical settings (including addiction, inflammation, and anxiety) [88][149]. The evidence shows a link between CB2 receptors regulation and the incidence of many mental and neurological conditions. CB2 receptors are distributed differently in the brain. Because they are mostly present within postsynaptic neuronal somatodendritic areas, triggering them may have different consequences than CB1 receptors [89][150]. Despite this, CB1 receptors, for example, are mostly found on nerve terminals, namely presynaptic GABAergic endings and dopamine neurons of ventral tegmental area (VTA) [90][151]. Whenever CB1 receptors are active, they reduce GABA distribution onto dopamine neurons, resulting in a spike in dopamine neuronal activity via a disengagement process. CB2 receptors, on the other hand, are mostly found in postsynaptic somatodendritic areas, and their activation reduces the rate of firing and the responsiveness of VTA and dopamine neurons in the brain [91][152]. CB2 receptors tend to be the key for underpinning protective effects on neurons, and addressing CB2 receptors could provide a unique therapeutic method for addressing neuropsychiatric and brain-related conditions, avoiding the normal CB1 receptor-mediated negative consequences [92][93][153,154] (Figure 3).
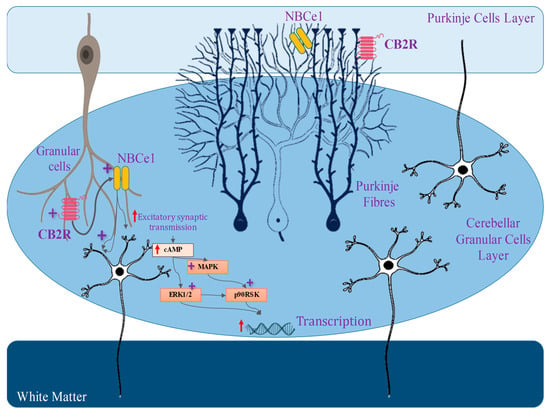
Figure 3.
Location of CB2 receptor in the brain, and its physiological retorts.
8. Cannabinoid Receptor 2 (CB2) in the Communication of Neuronal Signaling Pathways
As CB1 receptors are functioning, they reduce GABA discharge from presynaptic terminals, thereby reducing GABAergic neurons’ detrimental influence on the postsynaptic neurons. As a consequence of this disengagement process, postsynaptic neurons are activated. CB2 receptors, on the other hand, are mostly found in the cell nuclei of postsynaptic nerve cells [94][95][155,156]. As a result, the enrolment of such downstream CB2 receptors often result in a membrane potential hyperpolarization, which inhibits postsynaptic neuronal firing. Because of this discrepancy in distribution, CB1 and CB2 receptors activation produces opposite results. CB2 receptors expression reduces neuronal excitability via various methods. CB2 receptors expression reduces the excitability of neurons via a variety of methods. The CB2 receptors triggering, for example, operates for VTA dopamine neurons via modulating the K+ channel performance [94][155]. Intracellular CB2 receptors in frontal cortex neurons have connections to the Gq11-PLC-IP3 pathways, which initiates the development of Ca2+-dependent Cl- channels.
As a consequence, the cell membrane becomes hyperpolarized, resulting in neural blocking [96][157]. Studies also evidence that CB2 receptor expression activates the Na+/Bicarbonate co-transporter in the hippocampus, specifically in the CA3/CA2 neuronal cells, resulting in a sustained synaptic hyperpolarization. Interestingly, CB2 receptors activation is autonomous, changing the input and output behavior of CA3 pyramidal neurons and modulating gamma oscillations in vivo [95][156]. The increased expression of CB2 receptors in midbrain dopamine neuronal cells suggests that they may have an impact on a variety of key behaviors connected to dopamine activity [97][158]. CB2 receptors have been linked to the regulation of hunger, managing weight [98][99][100][101][159,160,161,162], depression [102][163], stress [103][164], and schizophrenia-like behaviors [104][165]. Several studies have demonstrated how CB2 receptors in the brain have an important role in reducing drug dependence—mainly alcohol, nicotine, and psychotropic substances [105][106][107][166,167,168]. These results imply that CB2 receptors have a considerable influence on the mesocorticolimbic system and perform essential functions in a wide range of brain processes, including psychiatric, cognitive abilities, and neurological tasks.
9. Role of Cannabinoid Receptor 2 (CB2) in the Modulation of Neuronal Physiology
Previous data suggested that CB2 receptors have been found in immune cell populations trafficking in the circulation, the spleen, and macrophage-derived cells, including cartilage cells and hepatocellular Kupffer cells [85][108][146,169]. Unlike CB1 receptors, which are present throughout the CNS, CB2 receptors are generally located in the brainstem and hippocampus CA2/3 pyramidal cells under normal physiological circumstances [95][109][156,170]. Nevertheless, if there is an inflammatory condition or destruction, the level of expression of CB2 receptors in activated microglial cells inside the CNS can be dramatically elevated [80][95][110][111][112][113][114][115][116][117][141,156,171,172,173,174,175,176,177,178]. These cells, which comprise indigenous microglia, perivascular microglia, astrocytes, and oligodendrocytes, account for more than 70% of the total cell population in the brain and spinal cord. They serve as the initial defense against inflammation and other forms of damage [118][119][179,180]. Microglia are the brain’s innate immune system cells and play a crucial part in both health and disease [80][141].
Microglia play an important role in initiating and maintaining synaptic remodeling inside neuronal cells under normal physiological circumstances. This is accomplished by changes in the immediate environment and changes in synapse architecture [120][121][122][123][124][181,182,183,184,185]. In cases of neurological pain, activated microglia in the nerve roots express CB2 mRNA [120][181]. Particularly, CB2 receptor activity is increased in the dorsal horn in a variety of neurological pain scenarios, involving regional damage to neurons, chemotherapy-induced neurological pain, and persistent post-ischemia pain. This increased expression is associated with microglia that have been activated [114][120][125][126][127][128][175,181,186,187,188,189]. CB2 receptors have been identified in the autopsy brain tissue of Alzheimer’s patients [129][130][190,191]. Additionally, receptors for CB2 were found in high concentrations along with high specificity in microglial cells linked to neuritic plaques [110][171]. Furthermore, CB2 receptors are upregulated in activated microglia cells from mouse models of numerous diseases, including Alzheimer’s disease, Huntington’s condition, and multiple neurological disorders [131][192]. A negative feedback process helps to restore homeostasis in numerous physiological stress circumstances, like wound healing. The Toll-like receptor (TLR) stimulation inside microglial cells triggers a release of suppression systems’ cytokine signaling molecules [132][193], while members of the tumor necrosis factor-alpha (TNFα)-induced protein 8 families are known to control responses to inflammation [133][194].
Enhanced CB2 receptor manifestation and expression could indicate an aggressive approach to limit or modulate the inflammatory process. The analogy might be related to the way tumor suppressor genes limit cell proliferation and how ephrin receptors in the brain can have different actions depending on the conditions [134][195]. Several studies have shown that CB2 receptor stimulation helps to reduce short-term inflammation [135][136][137][138][139][140][196,197,198,199,200,201]. Whenever the CB2 receptor system is activated, it suppresses neuroinflammatory signaling pathways and restores glial activity to normal—transitioning from a pro-inflammatory to an anti-inflammatory condition. CB2 receptors additionally influence the triggering of ERK1/2 (extracellular signal-regulated protein kinases) [141][202]. Earlier studies also evidence that toll-like receptor 2 (TLR2) and TLR4 have been linked to ERK1/2 amplification [142][203]. Experimental evidence shows that anandamide, which is an endocannabinoid, works via the MAPK pathway inside the CNS immune system. This strategy attempts to reduce the extent of inflammation and limit immunological responses associated with neurotoxicity [141][202].
A prior investigation has shown that using a particular CB2 agonist named 1-(3-benzyl-3-methyl-2,3-dihydro-1-benzofuran-6-yl) carbonyl) piperidine (MDA7) [143][204] affects the expression of gene modulation in the neuroinflammatory state generated by paclitaxel. This is evidenced by the considerably reduced expression of TLR2, CB2 receptors, and ERK1/2 [144][205]. In addition, the use of MDA7 has been linked to changes in cerebral glutamatergic communication [144][205] (Figure 4).
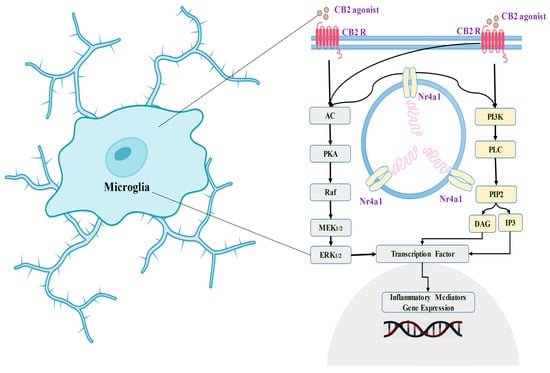
Figure 4.
Location of CB2 receptor in the microglia, and its physiological retorts.
10. The Involvement of the Cannabinoid Receptor 2 (CB2) in Synaptic Modulation
CB2 was first identified as a receptor found in distal macrophages [108][169]. Its existence in the brain was later validated by methods such as RT-PCR, in situ hybridization, and immunofluorescence studies. Regardless of the fact that CB2 levels throughout the brain are substantially less than those in the immune system [145][206], the receptor itself is found in microglial cells but not in astrocytes [146][207]. CB2 transcription is significantly increased in the face of prolonged pain [120][147][148][181,208,209]. CB2 was discovered within both astrocytes linked to neuritic deposits and microglia [110][171]. A new investigation identified that CB2 in the brainstem is biologically linked to vomiting control in collaboration with CB1 [149][210]. Previous research, nevertheless, demonstrated that CB2 mRNA was not found in the brain using methods such as RT-PCR and immunoblotting procedures [150][211].
A number of investigations have found neuronal CB2 activity in various brain locations [10][150][151][71,211,212], with CB2 being found in cell bodies of neurons and dendrites but not in synaptic endpoints [150][211]. Researchers effectively identified the existence of CB1 and CB2 mRNAs in the cerebral cortex of rats and mice using particular probes and primers. Researchers also provided evidence demonstrating the existence of CB2 receptors in numerous areas of the adult rat brain. The researchers used multiple antibodies from various sources, all engineered to target a distinct epitope on the CB2 receptor. When these antibodies were used to analyze the lymph nodes and other brain areas of rodents, they produced very comparable marking characteristics. The analogous antibody staining features seen in the lymph nodes and cerebellum provide evidence of this relationship. CB2-positive activity was also found in neurons optimistic for neuronal-specific enolase (NSE) in basic hippocampal preparations. The Western blot tests using extracts from the rodent’s spleen and brain revealed similar lines at the predicted molecular mass, which corresponded to the anticipated magnitude of the CB2 receptor. Immunohistochemical examinations revealed strong CB2 antibody staining across multiple neurons as well as glial cells throughout the brain.
11. Cannabinoid Receptor 2 (CB2) Gene Expression in the Brain
Earlier findings postulated that CB2 is located on chromosome 4 in rodents and on 1p36 in humans. Its genomic organization is straightforward, with a single translating exon. CB2 belongs to the GPCR receptor class and is made up of a single polypeptide chain with seven transmembrane α-helices. A glycosylated N-terminus is outside the cell, whereas an uninvolved C-terminus is inside. The CB2 region has only 44% nucleotide sequence analogy to the CB1 receptor. Whereas CB1 receptors are found in the brain, there is insufficient proof of CB2 binding to receptors, proteins, or mRNA in neurons [152][46]. Nonetheless, considerable quantitative evidence reveals that CB2 receptors are present in cultivated cerebral cell granules [153][213]. Advancement in cannabis pharmacotherapy has resulted in the discovery of several specialized antagonists and agonists that target these receptor categories [154][214]. The samples of proteins comprising rodent brain and spleen lysates revealed a conspicuous band at around 53 kDa, as well as further identifiable bands at roughly 37 kDa and 75 kDa. This discovery is consistent with current research [149][210]. It was additionally demonstrated that the location of CB2 receptors is particular by using three unique anti-CB2 antibodies, which were isolated according to their specificity, and targeting various amino acid endings, leading to equivalent marking trends.
The stains demonstrating the existence of CB2 receptors were ubiquitous throughout the cerebral cortex segments studied, including the orbital cortex, cerebral cortex, visual motor cortex, and auditory cortex. In layers III and V, the cell soma and top sections of pyramid-shaped neurons are heavily stained. In pyramid-shaped neurons in the hippocampus allocortex, an intermediate to significant CB2 immunostaining trend was seen. Such marking is prominent in the hippocampal CA2 and CA3 areas whereas it was less visible in the subiculum [147][149][208,210]. CB2 immunoreactivity was additionally detected in synapses situated in the stratum oriens and stratum radiatum. In addition, numerous glial cells showed immunostaining. Distinct neuronal soma-like and process-like CB2 staining patterns have been identified in the thalamus. The bulk of thalamic nuclei exhibited staining similar to cell bodies. Notably, well-defined processes were labeled within the reticular thalamic nucleus, featuring a dense network of fibers with CB2-like immunoreactivity. Nonetheless, CB2-positive labeling was seen in the cell nuclei of the ventral posterior thalamus region [155][215]. These structures are around 10–20 m in size.
The lateral posterior thalamic nucleus, the posterior thalamic nuclear category, including the paracentral thalamic nucleus all showed significant CB2 cell body marking. Cell bodies with fairly abundant CB2 immunopositivity were seen in the periaqueductal grey area. The greatest degree of staining was detected in the para-trochlear nucleus, the paralemniscal nucleus, and the red nuclei. The pontine nucleus constituted the most significant CB2-positive area inside the pons [156][216]. The pontine nucleus was defined by CB2-positive staining. The highly stained pathway seems to be an astrocyte or microglial process. CB2-like immunoreactivity was seen in substantia nigra pars reticulata, with clear labeling in the cell body, and some neuronal processes were noticeable. CB2-positive interneurons are larger than 20 m in size. The CB2-positive labeling appeared across the cerebellum lobes. Purkinje cell bodies were highly tarnished, but dendrites in the cellular tier were just faintly labeled. Numerous heavily stained fine puncta were seen in the molecular layer [145][206].