Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 2 by Sunayana Dagar and Version 1 by Sunayana Dagar.
Tunneling nanotubes (TNTs) are thin, hollow plasma membrane projections that directly connect the lumen of one cell to the lumen of another cell, thereby transferring different cargoes between the two connected cells. TNTs have been shown to play very prominent roles in neuronal development and serve as highways for neurodegenerative diseases in the brain.
- tunneling nanotubes
- Rhes
- neurodegenerative diseases
- mutant huntingtin
1. TNT Architecture
TNTs, characterized by their thin, long, and flexible membranous structure, allow for the transfer of organelles, vesicles, and proteins between cells [1][2][3]. Unlike other cellular components, TNTs lack specific markers for identification, and thus, their morphological features, such as their diameter and length, and their ability to grow above the substratum in 2D culture, serve as the primary criteria for recognition. Based on diameter, TNTs can be loosely classified into two groups: those with a diameter of less than 0.7 µm, which are primarily composed of actin and carry small membranous cargoes, and those with a diameter larger than 0.7 µm, which contain both actin and microtubules and are capable of transporting larger cytoplasmic components [4]. The post-translational modifications (PTMs) like acetylation and detyrosination on tubulin in swine testicle cells have been shown to maintain the stability of tunneling nanotubes (TNTs) over longer distances [5]. In 2D culture, TNTs can be observed as structures that appear to “hang” between the connected cells. They are often transient in nature and exhibit varying lifetimes. For example, in neuronal cells, TNTs can last for minutes [6][7], while in normal rat kidney cells, they can persist for several hours [8][9]. While there is still much to be learned, it is generally understood that TNTs can be generated by two distinct mechanisms, growth, and extension, like filopodia, or through the gradual detachment of adhering cells while preserving filamentous connections. TNTs can establish closed-ended or open-ended connections with target cells, but the mechanism for such selections remains unknown [2][10].
A recent study utilized a combination of optical and cryo-electron microscopy techniques to analyze the architecture of TNTs in neuronal Cath. a-differentiated (CAD) and SH-SY5Y cells. Surprisingly, the study revealed that multiple individual TNTs (iTNTs) were bundled together through N-Cadherin-positive connections. These iTNTs contained various membrane-enclosed compartments and mitochondria, which appeared to be transported along parallel F-actin bundles with the assistance of molecular motors [11].
The speed of communication through TNTs varies depending on the type of material being transported. Electrical signal transduction via TNTs occurs within milliseconds, while the transport of organelles relies on motor proteins and is associated with rapid TNT polymerization. The spread of protein aggregates within TNTs is 2–8 times faster compared to other cellular components [12]. For instance, Tau aggregates move at a speed of 2.83 ± 1.99 µm/min in TNTs [13], and the transfer of viruses through TNTs exhibits an average speed of 0.08 ± 0.03 µm/s, which is approximately 2–5 times faster than the movement of the murine leukemia virus along filopodial connections [14]. Endosomes have a velocity of approximately 0.72 µm/min when transported by Rhes-mediated TNTs, which is approximately 150 times slower than the retrograde transportation of cargo observed in neurons. [2][15]. Actin, which is the major constituent of TNTs, represents a crucial focus for TNT imaging[16]. It is hypothesized that F-actin-associated myosin motors play a role in facilitating the transportation of cellular components within TNTs [17][18][19]. Although microtubules have been detected in some TNTs, their exact role in TNTs remains unclear [20][21].
2. TNT-Like Structures in Development
Do TNTs have a role in brain development? According to the research findings, it has been observed that migrating neural crest cells (NCCs) establish and sustain ongoing communication with neighboring NCCs via both short and long filopodia during the process of neurogenesis. These connections play significant roles in guiding the directional movement of NCCs by allowing cells to adjust their course based on the migration of neighboring cells [22][23]. This suggests that these contacts are crucial for coordinating the guidance of NCCs. Interestingly, TNTs have been found to be vital in early neuronal development by facilitating the transfer of calcium ions, which are necessary for regulating the proliferation, migration, and differentiation of neurons [24]. In the initial stages of maturation, developing neurons establish electrical coupling and exchange calcium signals with astrocytes through TNTs [7]. TNTs also enable the bidirectional transmission of electrical signals between NCC cells through the expression of connexin 43 [25]. TNTs can also bridge gaps during neural tube closure and contribute to the closure of the neural tube in the midbrain of mammalian embryos [26]. Thus, it is predicted that TNTs play roles in neuronal development and in the transmission of electrical signals as a critical mechanism in immature neuronal circuits [3]. They may contribute to the ongoing synchronization and coordination of migratory activity among NCCs within expanding tissues during embryogenesis.
3. TNTs in Glial Cells
Effective intercellular communication is essential between neurons and glial cells for the proper functioning and development of the brain [27]. TNTs have been reported to play a pivotal role in the brain’s detoxification process by removing toxic misfolded proteins via glial cells [28]. For example, TNTs have been shown to transport fibrillar α-synuclein (α-syn) from overloaded astrocytes to microglia, where it undergoes degradation [29]. Moreover, overloaded microglia can form TNTs, both in vitro and in vivo, to release excessive α-syn aggregates to neighboring microglia, which help in their clearance [29]. During such a process, TNTs can prevent cell death and inflammation by transferring the mitochondria from naïve microglia to affected microglia [29]. Astrocytes and microglia can employ TNTs to facilitate the transfer of α-syn and mitochondria between astrocytes [30][31]. In another study, the authors demonstrate the occurrence of TNTs between human neuronal and microglial cells [30]. In this study, microglia showed a preference for transferring microglia to neuronal cells burdened with α-syn. These studies indicate that glial–TNT communication plays a critical role in decreasing the burden of accumulated protein aggregates and in promoting the mitochondria-mediated rescue mechanisms in neurodegenerative disease [30][31].
4. TNTs May Serve as Highways for Neurodegenerative Diseases in the Brain
TNTs are likely to have a crucial function in facilitating the transportation of different cargoes between brain cells. These cargoes include organelles, signaling molecules, pathogenic proteins, and microbes [10][28][30][32][33]. The transmission facilitated by TNT has the potential to influence both the physiological and pathological conditions of an organism. In particular, the intercellular transfer of various misfolded protein aggregates through TNTs holds significant implications for cellular communication and the development and progression of neurological disorders.
Neurodegenerative diseases such as Creutzfeldt JD, AD, PDakob disease (CJD), Alzheimer's disease (AD), Parkinson's disease (PD), and HDuntington's disease (HD) share a common characteristic of protein aggregates that accumulate in the central nervous system, leading to selective neuronal cell death [34]. Recent research suggests that the progression of these diseases is associated with the spreading of these protein assemblies, which can influence the misfolding of other cellular proteins [13][34][35]. Specifically, there is evidence indicating that disease proteins, such as prion protein (PrP), amyloid beta (Aβ), tau, alpha-synuclein (α-syn), and huntingtin, can be transferred between cells through TNTs [30][34][36][37] (Figure 1).
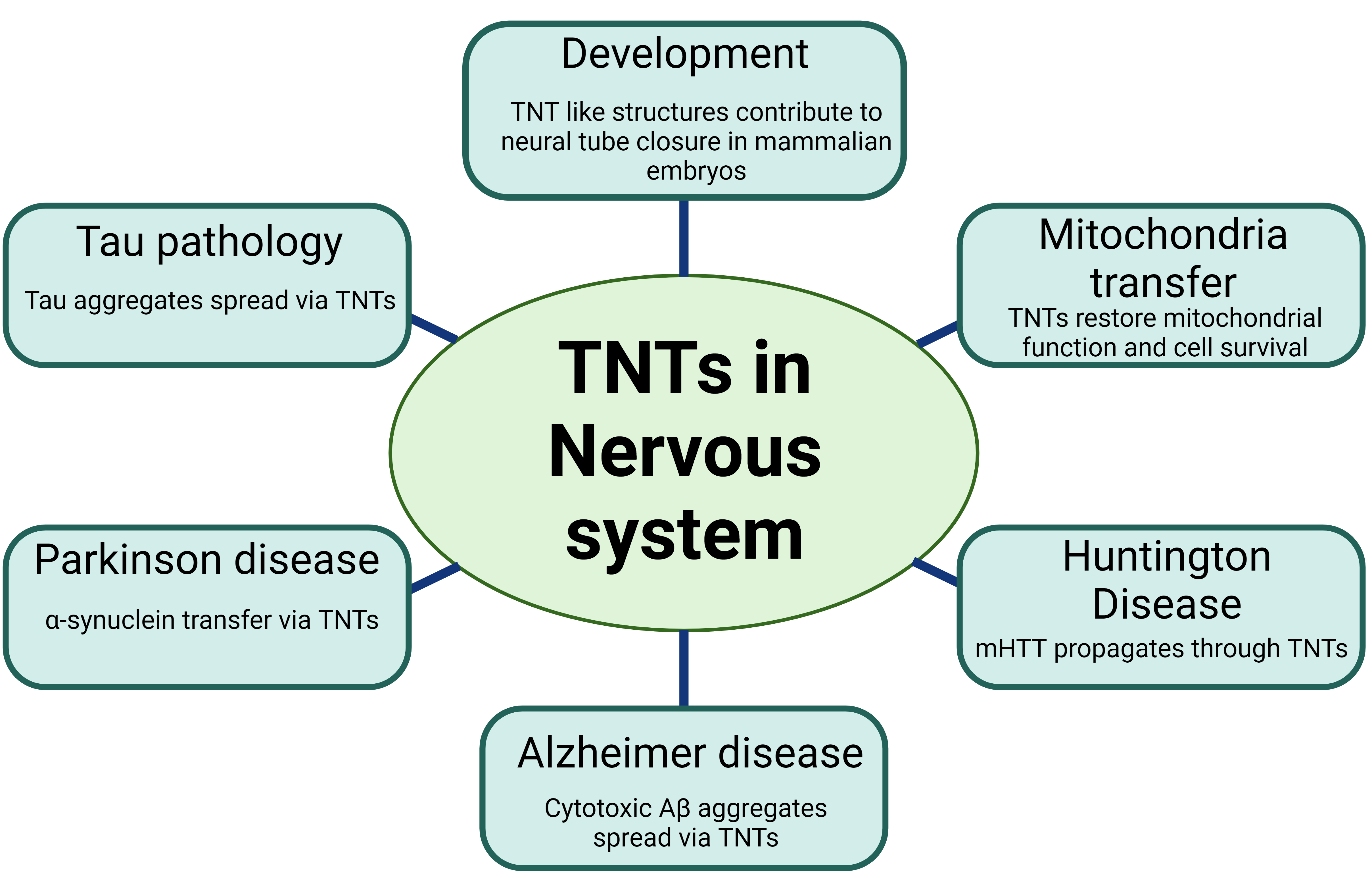
Figure 1. Various roles of Tunneling Nanotubes (TNTs) in the nervous system
Studies suggest that prion protein can be transported through TNTs in two ways in CJD [38]. Firstly, due to the GPI-anchored nature of PrP, it can migrate along the surface of TNTs. Secondly, PrP transfer within TNTs may involve specific organelles or vesicles such as specific endosomal compartments [35][38][39][40]. Prions can also induce the formation of TNTs to facilitate their own intercellular transmission via endosomal vesicles [38]. Similarly, in rat hippocampal astrocytes and neurons, TNT-like structures are involved in the transfer of PrP and Aβ aggregates that are associated with AD [41][42]. Aβ is known to have toxic effects on neuronal cells. Cytotoxic Aβ aggregates can utilize TNTs as a means of spreading, thereby causing neurotoxicity [36][41]. Recent research suggests that oxidative stress and serum deprivation can unidirectionally transport extracellular Aβ through TNTs in rat hippocampal astrocytes and neurons. Interestingly, it was demonstrated that the transport speed of cytotoxic Aβ via TNTs is faster than that of cellular organelles [36][41], suggesting that TNTs can serve as superhighways for Aβ spreading and potential disease progression.
Like Aβ, the α-syn fibrils also spread through TNTs between various cells, including neurons, human neuronal precursor cells (hNPCs), and astrocytes, involving the Wnt/Ca2+ pathway [30][43][44][45]. Thus, the cell-contact-dependent transfer appears to be a more efficient cell–cell transfer method for disease-relevant proteins. α-syn fibrils are shuttled predominantly inside lysosomal vesicles through TNTs involving ROS production [46][47]. Such transfer may also involve astrocyte intermediates, which further drive α-syn transmission to healthy astrocytes via TNTs, which could deliver mitochondria back to stressed astrocytes as two-way traffic [30][48]. Additionally, astrocytes can mediate the intercellular transfer of α-syn/MHC-II deposits through TNTs, indicating their involvement in the spreading of inflammation [49]. Thus, TNTs likely play multiple roles in the pathophysiological progression of PD.
In the case of HD, mutant huntingtin (mHTT) aggregates in neuronal cells and primary neurons increase the number of TNTs and promote their own spread through intercellular TNT-mediated communication [15]. Our recent research findings suggest that the Rhes protein, functioning as a multifunctional GTPase resembling SUMO E3 ligase, plays a vital role in the formation of TNTs and the facilitation of mHTT transportation between striatal medium spiny neurons (MSNs) and the cortex; the brain circuit is known to be highly vulnerable to degeneration in HD [2][15][50][51]. This transport of mHTT via Rhes is partly dependent on the SUMO posttranslational modification of mHTT [2][15][50][52][53]. An additional investigation is necessary to clarify the precise molecular mechanisms behind the propagation of mHTT through Rhes-TNTS and its possible contribution to the stereotypical degeneration observed in HD [54]. Similarly, misfolded α-synuclein aggregates, implicated in PD, are efficiently transferred through TNTs in primary neurons, CAD cells, SH-SY5Y cells, primary human brain pericytes derived from postmortem PD brains, and patient monocyte-derived microglia [29][43][45][46][55]. These aggregates often travel inside lysosomal vesicles and can seed soluble α-synuclein aggregation in the recipient cell cytosol [55].
The transfer of tau proteins across neuronal cells has been found to be facilitated by TNTs in astrocytes and neurons [13][41]. Both extracellular monomeric and fibrillar forms of Tau have been found to induce the formation of TNTs and promote their transfer between neuronal cells [13][32][56]. These observations support the idea that TNTs could serve as a means of spreading proteins that are involved across various neurodegenerative diseases. Therefore, further research is necessary to investigate the underlying mechanisms and pathways involved in the TNT-mediated mechanisms in neurodegenerative disease to explore avenues for effective therapies.
References
- Kimura, S.; Hase, K.; Ohno, H. Tunneling nanotubes: Emerging view of their molecular components and formation mechanisms. Exp. Cell Res. 2012, 318, 1699–1706.
- Subramaniam, S. Rhes Tunnels: A Radical New Way of Communication in the Brain’s Striatum? Bioessays 2020, 42, e1900231.
- Zurzolo, C. Tunneling nanotubes: Reshaping connectivity. Curr. Opin. Cell Biol. 2021, 71, 139–147.
- Drab, M.; Stopar, D.; Kralj-Iglič, V.; Iglič, A. Inception Mechanisms of Tunneling Nanotubes. Cells 2019, 8, 626.
- Jansens, R.J.J.; Van den Broeck, W.; De Pelsmaeker, S.; Lamote, J.A.S.; Van Waesberghe, C.; Couck, L.; Favoreel, H.W. Pseudorabies Virus US3-Induced Tunneling Nanotubes Contain Stabilized Microtubules, Interact with Neighboring Cells via Cadherins, and Allow Intercellular Molecular Communication. J. Virol. 2017, 91, 10-1128.
- Gerdes, H.H.; Rustom, A.; Wang, X. Tunneling nanotubes, an emerging intercellular communication route in development. Mech. Dev. 2013, 130, 381–387.
- Wang, X.; Bukoreshtliev, N.V.; Gerdes, H.H. Developing neurons form transient nanotubes facilitating electrical coupling and calcium signaling with distant astrocytes. PLoS ONE 2012, 7, e47429.
- Gurke, S.; Barroso, J.F.; Hodneland, E.; Bukoreshtliev, N.V.; Schlicker, O.; Gerdes, H.H. Tunneling nanotube (TNT)-like structures facilitate a constitutive, actomyosin-dependent exchange of endocytic organelles between normal rat kidney cells. Exp. Cell Res. 2008, 314, 3669–3683.
- Gurke, S.; Barroso, J.F.; Gerdes, H.H. The art of cellular communication: Tunneling nanotubes bridge the divide. Histochem. Cell Biol. 2008, 129, 539–550.
- Dagar, S.; Pathak, D.; Oza, H.V.; Mylavarapu, S.V.S. Tunneling nanotubes and related structures: Molecular mechanisms of formation and function. Biochem. J. 2021, 478, 3977–3998.
- Sartori-Rupp, A.; Cordero Cervantes, D.; Pepe, A.; Gousset, K.; Delage, E.; Corroyer-Dulmont, S.; Schmitt, C.; Krijnse-Locker, J.; Zurzolo, C. Correlative cryo-electron microscopy reveals the structure of TNTs in neuronal cells. Nat. Commun. 2019, 10, 342.
- Wang, Y.; Cui, J.; Sun, X.; Zhang, Y. Tunneling-nanotube development in astrocytes depends on p53 activation. Cell Death Differ. 2011, 18, 732–742.
- Tardivel, M.; Begard, S.; Bousset, L.; Dujardin, S.; Coens, A.; Melki, R.; Buee, L.; Colin, M. Tunneling nanotube (TNT)-mediated neuron-to neuron transfer of pathological Tau protein assemblies. Acta Neuropathol. Commun. 2016, 4, 117.
- Sowinski, S.; Jolly, C.; Berninghausen, O.; Purbhoo, M.A.; Chauveau, A.; Köhler, K.; Oddos, S.; Eissmann, P.; Brodsky, F.M.; Hopkins, C.; et al. Membrane nanotubes physically connect T cells over long distances presenting a novel route for HIV-1 transmission. Nat. Cell Biol. 2008, 10, 211–219.
- Sharma, M.; Subramaniam, S. Rhes travels from cell to cell and transports Huntington disease protein via TNT-like protrusion. J. Cell Biol. 2019, 218, 1972–1993.
- Dagar, S.; Pushpa, K.; Pathak, D.; Samaddar, S.; Saxena, A.; Banerjee, S.; Mylavarapu, S.V.S. Nucleolin regulates 14-3-3ζ mRNA and promotes cofilin phosphorylation to induce tunneling nanotube formation. FASEB J 2021, 35(1), e21199.
- Gousset, K.; Marzo, L.; Commere, P.H.; Zurzolo, C. Myo10 is a key regulator of TNT formation in neuronal cells. J. Cell Sci. 2013, 126, 4424–4435.
- Sun, Y.Y.; Yang, Y.F.; Keller, K.E. Myosin-X Silencing in the Trabecular Meshwork Suggests a Role for Tunneling Nanotubes in Outflow Regulation. Investig. Ophthalmol. Vis. Sci. 2019, 60, 843–851.
- Uhl, J.; Gujarathi, S.; Waheed, A.A.; Gordon, A.; Freed, E.O.; Gousset, K. Myosin-X is essential to the intercellular spread of HIV-1 Nef through tunneling nanotubes. J. Cell Commun. Signal 2019, 13, 209–224.
- Resnik, N.; Erman, A.; Veranič, P.; Kreft, M.E. Triple labelling of actin filaments, intermediate filaments and microtubules for broad application in cell biology: Uncovering the cytoskeletal composition in tunneling nanotubes. Histochem. Cell Biol. 2019, 152, 311–317.
- Resnik, N.; Prezelj, T.; De Luca, G.M.R.; Manders, E.; Polishchuk, R.; Veranič, P.; Kreft, M.E. Helical organization of microtubules occurs in a minority of tunneling membrane nanotubes in normal and cancer urothelial cells. Sci. Rep. 2018, 8, 17133.
- Tarasiuk, O.; Scuteri, A. Role of Tunneling Nanotubes in the Nervous System. Int. J. Mol. Sci. 2022, 23, 12545.
- Teddy, J.M.; Kulesa, P.M. In vivo evidence for short- and long-range cell communication in cranial neural crest cells. Development 2004, 131, 6141–6151.
- Spitzer, N.C. Electrical activity in early neuronal development. Nature 2006, 444, 707–712.
- Wang, X.; Veruki, M.L.; Bukoreshtliev, N.V.; Hartveit, E.; Gerdes, H.H. Animal cells connected by nanotubes can be electrically coupled through interposed gap-junction channels. Proc. Natl. Acad. Sci. USA 2010, 107, 17194–17199.
- Pyrgaki, C.; Trainor, P.; Hadjantonakis, A.K.; Niswander, L. Dynamic imaging of mammalian neural tube closure. Dev. Biol. 2010, 344, 941–947.
- Tanriover, G.; Sozen, B.; Seker, A.; Kilic, T.; Gunel, M.; Demir, N. Ultrastructural analysis of vascular features in cerebral cavernous malformations. Clin. Neurol. Neurosurg. 2013, 115, 438–444.
- Lagalwar, S. Mechanisms of tunneling nanotube-based propagation of neurodegenerative disease proteins. Front. Mol. Neurosci. 2022, 15, 957067.
- Scheiblich, H.; Dansokho, C.; Mercan, D.; Schmidt, S.V.; Bousset, L.; Wischhof, L.; Eikens, F.; Odainic, A.; Spitzer, J.; Griep, A.; et al. Microglia jointly degrade fibrillar alpha-synuclein cargo by distribution through tunneling nanotubes. Cell 2021, 184, 5089–5106.e21.
- Chakraborty, R.; Nonaka, T.; Hasegawa, M.; Zurzolo, C. Tunnelling nanotubes between neuronal and microglial cells allow bi-directional transfer of α-Synuclein and mitochondria. Cell Death Dis. 2023, 14, 329.
- Rostami, J.; Holmqvist, S.; Lindström, V.; Sigvardson, J.; Westermark, G.T.; Ingelsson, M.; Bergström, J.; Roybon, L.; Erlandsson, A. Human Astrocytes Transfer Aggregated Alpha-Synuclein via Tunneling Nanotubes. J. Neurosci. 2017, 37, 11835–11853.
- Abounit, S.; Wu, J.W.; Duff, K.; Victoria, G.S.; Zurzolo, C. Tunneling nanotubes: A possible highway in the spreading of tau and other prion-like proteins in neurodegenerative diseases. Prion 2016, 10, 344–351.
- Padmanabhan, S.; Manjithaya, R. Leaderless secretory proteins of the neurodegenerative diseases via TNTs: A structure-function perspective. Front. Mol. Neurosci. 2023, 16, 983108.
- Tiwari, V.; Koganti, R.; Russell, G.; Sharma, A.; Shukla, D. Role of Tunneling Nanotubes in Viral Infection, Neurodegenerative Disease, and Cancer. Front. Immunol. 2021, 12, 680891.
- Victoria, G.S.; Zurzolo, C. The spread of prion-like proteins by lysosomes and tunneling nanotubes: Implications for neurodegenerative diseases. J. Cell Biol. 2017, 216, 2633–2644.
- Dilna, A.; Deepak, K.V.; Damodaran, N.; Kielkopf, C.S.; Kagedal, K.; Ollinger, K.; Nath, S. Amyloid-β induced membrane damage instigates tunneling nanotube-like conduits by p21-activated kinase dependent actin remodulation. Biochim. Biophys. Acta Mol. Basis Dis. 2021, 1867, 166246.
- Wang, X.T.; Sun, H.; Chen, N.H.; Yuan, Y.H. Tunneling nanotubes: A novel pharmacological target for neurodegenerative diseases? Pharmacol. Res. 2021, 170, 105541.
- Zhu, S.; Victoria, G.S.; Marzo, L.; Ghosh, R.; Zurzolo, C. Prion aggregates transfer through tunneling nanotubes in endocytic vesicles. Prion 2015, 9, 125–135.
- Kuznetsov, I.A.; Kuznetsov, A.V. A two population model of prion transport through a tunnelling nanotube. Comput. Methods Biomech. Biomed. Eng. 2014, 17, 1705–1715.
- Victoria, G.S.; Arkhipenko, A.; Zhu, S.; Syan, S.; Zurzolo, C. Astrocyte-to-neuron intercellular prion transfer is mediated by cell-cell contact. Sci. Rep. 2016, 6, 20762.
- Zhang, K.; Sun, Z.; Chen, X.; Zhang, Y.; Guo, A.; Zhang, Y. Intercellular transport of Tau protein and β-amyloid mediated by tunneling nanotubes. Am. J. Transl. Res. 2021, 13, 12509–12522.
- Miyazawa, K.; Kipkorir, T.; Tittman, S.; Manuelidis, L. Continuous production of prions after infectious particles are eliminated: Implications for Alzheimer’s disease. PLoS ONE 2012, 7, e35471.
- Dieriks, B.V.; Park, T.I.; Fourie, C.; Faull, R.L.; Dragunow, M.; Curtis, M.A. α-synuclein transfer through tunneling nanotubes occurs in SH-SY5Y cells and primary brain pericytes from Parkinson’s disease patients. Sci. Rep. 2017, 7, 42984.
- Vargas, J.Y.; Loria, F.; Wu, Y.J.; Córdova, G.; Nonaka, T.; Bellow, S.; Syan, S.; Hasegawa, M.; van Woerden, G.M.; Trollet, C.; et al. The Wnt/Ca2+ pathway is involved in interneuronal communication mediated by tunneling nanotubes. EMBO J. 2019, 38, e101230.
- Dilsizoglu Senol, A.; Samarani, M.; Syan, S.; Guardia, C.M.; Nonaka, T.; Liv, N.; Latour-Lambert, P.; Hasegawa, M.; Klumperman, J.; Bonifacino, J.S.; et al. α-Synuclein fibrils subvert lysosome structure and function for the propagation of protein misfolding between cells through tunneling nanotubes. PLoS Biol. 2021, 19, e3001287.
- Okuda, S.; Uemura, N.; Takahashi, R. Alpha-synuclein fibrils propagate through tunneling nanotubes. Mov. Disord. 2017, 32, 394.
- Freeman, D.; Cedillos, R.; Choyke, S.; Lukic, Z.; McGuire, K.; Marvin, S.; Burrage, A.M.; Sudholt, S.; Rana, A.; O’Connor, C.; et al. Alpha-synuclein induces lysosomal rupture and cathepsin dependent reactive oxygen species following endocytosis. PLoS ONE 2013, 8, e62143.
- Valdinocci, D.; Kovarova, J.; Neuzil, J.; Pountney, D.L. Alpha-Synuclein Aggregates Associated with Mitochondria in Tunnelling Nanotubes. Neurotox. Res. 2021, 39, 429–443.
- Valdinocci, D.; Radford, R.A.; Siow, S.M.; Chung, R.S.; Pountney, D.L. Potential Modes of Intercellular α-Synuclein Transmission. Int. J. Mol. Sci. 2017, 18, 469.
- Subramaniam, S.; Boregowda, S. Curbing Rhes Actions: Mechanism-based Molecular Target for Huntington’s Disease and Tauopathies. CNS Neurol. Disord.—Drug Targets 2024, 23, 21–29.
- Ramirez-Jarquin, U.N.; Sharma, M.; Shahani, N.; Li, Y.; Boregowda, S.; Subramaniam, S. Rhes protein transits from neuron to neuron and facilitates mutant huntingtin spreading in the brain. Sci. Adv. 2022, 8, eabm3877.
- Subramaniam, S. Striatal Induction and Spread of the Huntington’s Disease Protein: A Novel Rhes Route. J. Huntington’s Dis. 2022, 11, 281–290.
- Ramirez-Jarquin, U.N.; Sharma, M.; Zhou, W.; Shahani, N.; Subramaniam, S. Deletion of SUMO1 attenuates behavioral and anatomical deficits by regulating autophagic activities in Huntington disease. Proc. Natl. Acad. Sci. USA 2022, 119, e2107187119.
- Del Rey, N.L.; Garcia-Cabezas, M.A. Cytology, architecture, development, and connections of the primate striatum: Hints for human pathology. Neurobiol. Dis. 2023, 176, 105945.
- Abounit, S.; Bousset, L.; Loria, F.; Zhu, S.; de Chaumont, F.; Pieri, L.; Olivo-Marin, J.C.; Melki, R.; Zurzolo, C. Tunneling nanotubes spread fibrillar α-synuclein by intercellular trafficking of lysosomes. EMBO J. 2016, 35, 2120–2138.
- Annadurai, N.; De Sanctis, J.B.; Hajdúch, M.; Das, V. Tau secretion and propagation: Perspectives for potential preventive interventions in Alzheimer’s disease and other tauopathies. Exp. Neurol. 2021, 343, 113756.
More