Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Charalampos Proestos and Version 2 by Peter Tang.
Polyphenols are a diverse group of compounds possessing various health-promoting properties that are of utmost importance for many wine sensory attributes. Apart from genetic and environmental parameters, the implementation of specific oenological practices as well as the subsequent storage conditions deeply affect the content and nature of the polyphenols present in wine.
- polyphenols
- wine authenticity
- metabolomics
- oenological practices
- biomarkers
1. Introduction
Polyphenols constitute a diverse group of bioactive compounds occurring in both grapes and wines [1]. In plants, they have been found to exhibit key roles in growth, fertility, and reproduction. They present protective properties against abiotic stress conditions such as UV-light and biotic stresses such as pathogen and predator attacks [2][3][2,3]. Polyphenols exhibit a significant role in modern food technology and human nutrition [4][5][4,5] and are frequently key ingredients in functional foods. The benefits derived from the moderated wine consumption for human health have been well elaborated [6][7][6,7] with several groups of phenolic compounds including stilbenes [8], flavonols [9], and proanthocyanidins [10] found to exert various health-promoting properties [6].
Wine production is regulated by OIV (International Organization of Vine and Wine), global wine policies, and national governments. Provision of authentic wines to the market has always been a prerequisite meaning that the declarations on the wine label should mirror the composition and provenance of this intriguing product [11]. However multiple cases of intentional or unintentional wine mislabeling [12][13][14][15][12,13,14,15] have been recorded, alarming wine consumers who demand for strict controls safeguarding wine transparency.
Polyphenols play a fundamental role in the determination of wine sensory attributes contributing greatly to its color, flavor, and taste. In wine trade, the organoleptic characteristics are the major determinants of wine consumers’ acceptance and preference. It is generally accepted that high-quality red wines exhibit sensory properties that reflect their chemical composition.
2. Structural Information
From a structural aspect, polyphenols can be divided into two main categories, the flavonoids that bear a common C6-C3-C6 skeleton and the non-flavonoids. Their name comes from the Greek (Ancient) word polus (that means “much, many”) and phenol, which refers to a chemical structure formed by the attachment of a hydroxyl (-OH) group to an aromatic benzenoid (phenyl) ring in a similar way to that found in alcohols. This section highlights the basic polyphenolic structures in relation to wine authenticity, however, it does not provide a detailed presentation of the polyphenolic profile of grapes and wine as this has been extensively described [16][17][18][19][17,18,19,20].2.1. Non-Flavonoid Polyphenols
Essentially non-flavonoid polyphenols are located in the grape pulp with the main classes comprising of phenolic acids and stilbenes as well as their derivatives. In wine, oak-derived non-flavonoids have also been detected including the classes of hydrolysable tannins (gallotannins and elagitannins), coumarins, and lignans [20][21][21,22].2.1.1. Phenolic Acids and Derivatives
Phenolic acids have been successfully employed for white wine authentication purposes [22][16]. They are divided into two groups, the hydroxybenzoic (HBA) and the hydroxycinnamic (HCA) acids (phenolic acids). HBAs share a common C6-C1 structure, referring to a benzene ring with one carbon aliphatic chain substituent. Different acids are formed by the substitution of the benzene ring, with p-hydroxybenzoic, protocatechuic, syringic, vanillic, and gallic acid regarded as the most abundant compounds in this sub-class Figure 1 [16][17]. HCAs have been detected and identified in both grapes and wines [23]. They are present in lower concentrations in their free form (ferulic, caffeic, p-coumaric, and sinapic acid) and as esters of tartaric acid (fertaric, caftaric, and coutaric acids) [24][25][24,25]. HCAs are the principal phenolic compounds of free-run juice and white wines [26]. High concentration of HCAs may provoke a slightly astringent mouthfeel [27]. Esters of gallic acid (gallotannins) and ellagic acid (ellagitannins) with glucose or related sugars are oak-derived non-flavonoids occurring in wine [28]. Ethyl esters [24] as well as various glycosides of other HBAs and HCAs have also been reported [29].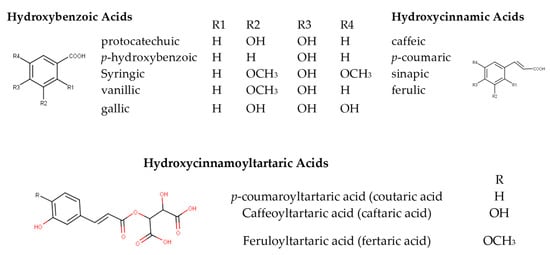
Figure 1.
Basic structures of phenolic acids in wine.
2.1.2. Stilbenes
Stilbenes are polyphenols featuring diverse biological properties and a complex structure that exhibits a limited but heterogeneous distribution in the plant kingdom [30]. Grapes are one of the richest sources of stilbenes that have also been detected in wines and oak wood. They share a common C6-C2-C6 skeleton, containing two benzene rings, usually bonded by an ethylene, or ethane chain. Resveratrol as well as specific oligomers collectively mentioned as “inducible” phytoalexins are produced by the grapevine as a defense response against pathogens [31] (Figure 2). Conversely “metabolized” viniferins are formed by enzymes released from pathogens in an attempt to eliminate toxic compounds [32]. Stilbenes are present as mono-, di-, and trimers and very complex oligomers. Resveratrol is probably the most studied compound of this subclass as it exhibits various therapeutic effects [33][34][35][36][37][33,34,35,36,37]. Studies have shown that resveratrol mimics effects of caloric restriction, exerts antioxidative anti-inflammatory properties, and is linked with the initiation and progression of many diseases through several mechanisms [37]. However, the major drawback as reported in most of the clinical trials is its poor bioavailability [37]. Resveratrol content is regulated by genetic and environmental factors. Viticultural practices aiming at lower yields as well as soft vinification protocols can promote/maintain the resveratrol content in wine [31]. Most monomer stilbenes in Vitis species are found as glycosylated or methoxylated derivatives of piceatannol and resveratrol [38]. Apart from trans-resveratrol they include trans and cis piceid (3,4’-5-Trihydroxystilbene-3-beta-D-glucopyranoside), pterostilbene (trans-3,5-dimethoxy-4’-hydroxystilbene), piceatannol (3,4,3′,5’-tetrahydroxy-trans-stilbene), and astringin (piceatannol 3-O-beta-D-glucoside). Dimer stilbenes usually contain a monomer resveratrol moety with the most commonly occurring compound being ε-viniferin. Figure 2 depicts a scheme of the formation of resveratrol oligomers and viniferins, initially reported from Bavaresco et al. (2016) [31]. Flamini et al. (2016) reported a significant increase of resveratrol (trans monomer, dimers, and two tetramers) and a simultaneous decrease in glycoside derivatives on Aspergillus. Carbonarious inoculated Negro Amaro cv grapes [32]. Pentamers of resveratrol have been found present in other plants but not in the Vitis vinifera [38]. Papastamoulis et al. (2014) reported the presence of the resveratrol hexamer Viniphenol A in vine stalks of V. vinifera [39].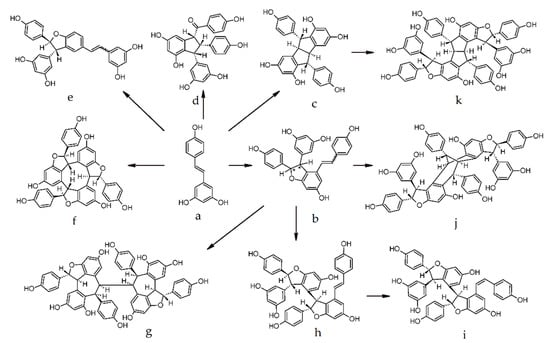
Figure 2. Scheme of formation of resveratrol oligomers and viniferins: (a) trans-resveratrol; (b) (E and Z) ε-viniferin/ ω-viniferin; (c) pallidol; (d) caraphenol B; (e) δ-viniferin (E and Z); (f) α-viniferin; (g) isohopeaphenol; (h) E-miyabenol C; (i) Z-miyabenol C; (j) vaticanol C isomer; and (k) ampelopsin H [31].
2.1.3. Lignans
Lignans are an important class of non-flavonoid secondary metabolites occurring in various plants. They are synthesized from shikimic acid through the phenylpropanoid pathway. The amount of lignans in wine has been found to vary on average from 0.22 mg/L for the whites to 0.80 mg/L for the reds constituting wine as an important source of lignans in comparison to other alcoholic beverages [20][21]. Selected compounds including some lignan and neolignan derivatives from lariciresinol, isolariciresinol, and secoisolariciresinol have been previously identified in both white and red wines [21][40][22,40].2.2. Flavonoid Compounds
Flavonoids are secondary metabolites occurring in a wide variety of natural products such as vegetables, fruits, stems, cocoa, tea, grapes, and wine. Nowadays flavonoids are considered essential ingredients for various medicinal, nutraceutical, pharmaceutical, and cosmetic applications. This is due to their potent anti-oxidative, anti-carcinogenic, anti-inflammatory, and anti-mutagenic properties combined with the ability to modulate significant enzyme functions [41][42][43][41,42,43]. In plants they are involved in a series of processes related to defense against pathogens and pests, protection from ultraviolet (UV) radiation, allelopathy, pollen fertilization, auxin transport regulation, and pigmentation [44][45][46][47][44,45,46,47]. In wine, flavonoids have a fundamental role in the determination of its sensory attributes specifically wine color, flavor, astringency, and bitterness [27][48][27,48]. Total phenolic content in red wine ranges from 1200 to 1800 mg gallic acid equivalents/L, which is six to nine times more than the corresponding content in white wines [49]. Chemically, they belong to a class displaying a diphenylpropane skeleton structure, essentially, two aromatic rings (rings A and B) linked by a three-carbon chain forming a closed pyran ring (heterocyclic ring containing oxygen, the C ring) with the A aromatic ring (Figure 3). In this C6-C3-C6 structure, usually the B aromatic ring is attached to position 2 of C ring and on fewer occasions, it is found attached to positions 3 or 4 [7][16][7,17]. The above possible combinations, the patterns of glycosylation and/or hydroxylation of the aromatic rings, the structural features of the rings, along with the degree of unsaturation and oxidation of the C ring account for the large diversity of the flavonoid compounds.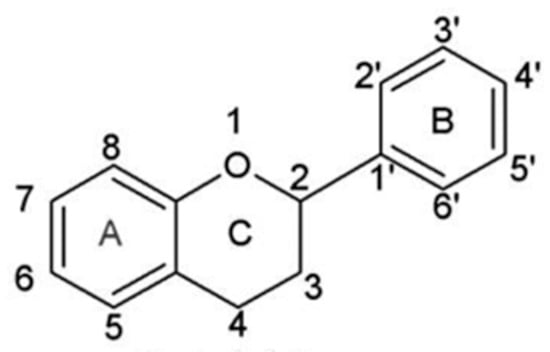
Figure 3.
Basic flavonoid structure.
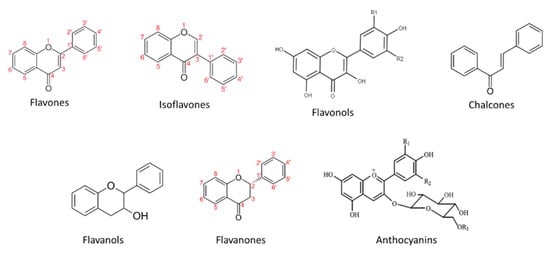
Figure 4.
Main flavonoid groups.
2.2.1. Flavones
Flavones display a double bond between positions 2 and 3 of the central ring and ketone group at position 4 (Figure 4). They are considered as an important group of various fruits and vegetables. Flavones are present as aglycons and glucosides, however, they are not found abundant in grapes except for luteolin derivatives [50]. Luteolin has been found to exhibit significant anti-oxidative, anti-tumor, cardioprotective, and anti-inflammatory properties [51]. Isoflavones are isomers of flavones where ring B is attached in position 3 of the central ring. Genistein and daidzein are compounds of this subclass found present in wine [52].2.2.2. Flavanones
Flavanones, also called dihydroflavones, display a saturated carbon chain meaning that the double bond between positions 2 and 3 is saturated in comparison to flavones. Eriodictyol, naringenin, and hesperitin are the main compounds identified in grapes and wine from this subclass [16][25][53][54][55][17,25,53,54,55].2.2.3. Flavonols
Flavonols are a diverse group of flavonoids present in the majority of higher plants. They are mainly localized in the outer epidermis of the grape berry skins and accumulate after flowering and during ripening [9][44][9,44]. They exhibit photo-protection and copigmentation properties [9], have been employed for chemotaxonomical studies [56][57][58][56,57,58], and feature various health-promoting properties [6]. Flavonols are considered as the building blocks for anthocyanins. Structurally, they are characterized by the presence of a double bond in positions 2 and 3 and a ketone group at position 4 of the central ring as well 2 hydroxyl groups at position 5 and 7 of Ring A. Various compounds arise from the different methylation, hydroxylation, and glycosylation patterns observed, constituting flavonols as one of the most important and diverse subclasses of phenolic compounds present in grapes and wine. In grapes they are commonly found as glycosides of the six main aglycons namely quercetin, kaempferol, isorhamnetin, myricetin, laricitrin, and syringetin (Figure 5). Castillo-Muñoz et al. (2010) reported that in the white grape varieties studied, flavonols were only found present as 3-O-glycoside derivatives, comprising of the series of 3-O-glucosides, 3-O-galactosides, and 3-O-glucuronides of quercetin and kaempferol along with 3-O-glucosides and 3-O-galactosides of isorhamnetin. In the same study, rutin was also detected as well as scarce traces of isorhamnetin 3-O-glucuronide, while myricetin, laricitrin, syringetin, and their derivatives were absent in all samples [56]. The latter absence has been attributed to the lack of expression of the enzyme flavonoid 3’,5’-hydroxylase in white grapes [9][59][9,59]. Mattivi et al. reported that quercetin derivatives were by far the predominant flavonols in white and light rose grape varieties, while myricetin was the most abundant flavonol in the red varieties studied [59]. However, Blancquaert et al. (2019) reported that the major flavonol glucosides of Cabernet Sauvignon grapes from Stellenbosch University vineyard were quercetin- 3-O-glucoside and quercetin-3-O-glucuronide and not myricetin derivatives as previously described. The same authors reported that flavonol accumulation was greatly influenced by light quality [44], which was also verified from Martínez-Lüscher et al. (2019) who in hand indicated that flavonol profile is a reliable indicator to assess canopy architecture and exposure of red wines to solar radiation [60]. Favre’ et al. (2018) employing HRMS (high-resolution mass spectrometry) metabolomics identified new acetylated and p-coumaroylated derivatives of the flavonol 3-O-glucosides of isorhamnetin, laricitrin, and syringetin in Vitis vinifera grape skins and wines [61].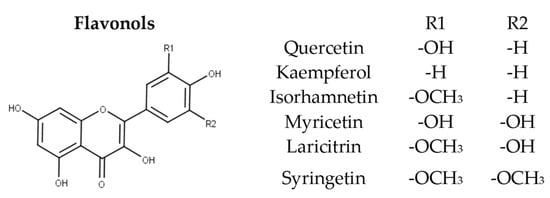
Figure 5.
Structure of the six main flavonol aglycons.
2.2.4. Flavan-3-ols
Flavan-3-ols, also named as flavanols or catechins, are mainly located in the solid parts of the berry (stem, skin, seeds), with seeds presenting the highest concentration [62]. Structurally monomeric flavan-3-ols are benzopyrans displaying a carbon chain between C2 and C3, a hydroxyl group at position three of the central ring, and absence of carbonyl moiety in position 4 [16][17]. Flavanols possess two chiral carbons, able to form four diastereoisomers for each of them. In Vitis vinifera grape varieties, (+)-catechin and (−)-epicatechin are the principal compounds followed by (+)-gallocatechin, (−)-epigallocatechin, and the galloylated form of (−)-epicatechin (i.e., (−)-epicatechin gallate), which is found at lower concentrations [17][63][18,63] (Figure 6). Flavan-3-ols are responsible for influencing the mouthfeel of wines imparting astringency. This tactile sensation results from flavanol interaction with salivary proteins and glycoproteins causing a loss of lubricating power loss of the saliva or with the glycoproteins of the mouth epithylium [64]. It has been reported that flavanol bitterness decreases from the monomer to trimer flavan-3-ols.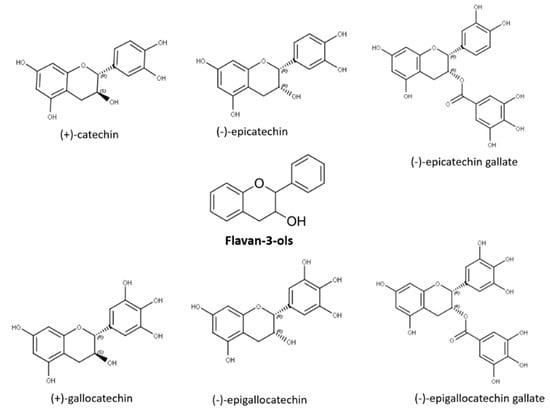
Figure 6.
Flavan-3-ol monomeric forms.
2.2.5. Anthocyanins
Anthocyanins present an essential role in red wines as they impact on wine color as well as on reactions affecting the stability and longevity of red wines [67][70]. More than 540 anthocyanin pigments have been found in nature [67][70]. They exhibit a wide range of biological activities including antioxidant, anti-inflammatory, antimicrobial, antimutagenic, and anticarcinogenic properties [68][71]. In V. vinifera grape varieties, they are mainly localized on red berry skins where they are synthesized from veraison until full ripening. Their content is significantly affected by vintage and overall vineyard management practices [9]. They have been also found present on the skins of some pink-skinned varieties such as Gewurtztraminer and Muscat Rouge de Madere [59] and in the pulp of the so called “teinturer” V. vinifera (i.e., Alicante Bouchet) and non V. vinifera grape varieties [69][70][71][72,73,74]. Malvidin, delphinidin, peonidin, cyanidin, petunidin, and lately pelargonidin [72][73][75,76] are regarded as the most important anthocyanidins in wine (Figure 7). Their differentiation lies in the substituent groups present in the B-ring. In Vitis vinifera grape varieties, the aglycon (anthocyanidin) is bonded through a glycosidic bond in position 3 of the ring C to a sugar moiety, commonly glucose. Malvidin-3-O-glucoside has been proposed as the dominant anthocyanin in red grape varieties [16][44][59][62][17,44,59,62]. Moreover, acylation can occur on the glucose moiety with aromatic (such as caffeic, ferulic, sinapic, and p-coumaric acids) or aliphatic acids (such as acetic, malic, malonic, oxalic, and succinic acids) probably due to the presence of the enzyme anthocyanin acyl-transferase [9].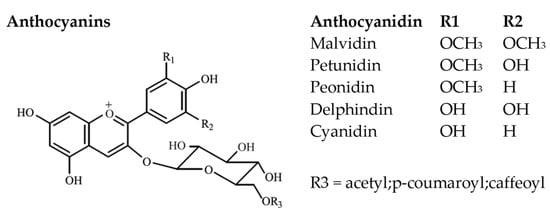
Figure 7.
Main anthocyanin chemical structures.
-
Self-association and co-pigmentation (short-term mechanisms),
-
Polymeric anthocyanins with proanthocyanidins and 3-flavanols (long-term mechanisms) and
-
New formed compounds, such as pyranoanthocyanins and their polymerized compounds.