Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 3 by Sirius Huang and Version 2 by Sirius Huang.
Pathogens and pests constantly challenge food security and safety worldwide. The use of plant protection products to manage them raises concerns related to human health, the environment, and economic costs. Basic substances are active, non-toxic compounds that are not predominantly used as plant protection products but hold potential in crop protection. Basic substances’ attention is rising due to their safety and cost-effectiveness.
- disease management
- pest management
- sustainable crop protection
- integrated pest management
- organic farming
1. Introduction
The use of plant protection products, such as fungicides, insecticides, and herbicides, is crucial for controlling diseases and pests in agriculture, but their safety, costs, and availability are a growing concern [1][2][3][4]. The possible adverse effects on human and environmental health have led to the development of risk exposure indicators [5][6] and more stringent legislative requirements [7][8]. The EU, for example, regulates plant protection products authorization [9] and utilization to endorse a new paradigm for agricultural production with the transition to low-input farming, promoting integrated pest management and complementary alternatives to minimize the utilization of plant protection products [10]. The use of plant protection products has negative impacts, in terms of their direct costs and negative externalities, on producers and the environment, especially in developing countries [11]. While ensuring rigorous testing for safety and quality, the product registration process increases the costs of developing new products and lengthens the time to market [12][13]. Additionally, the shift towards single-site compounds, which have a more favorable profile than multi-site compounds, increases the risk of resistance development in pests and pathogens [14].
Basic substances can represent an opportunity to mitigate the problems associated with traditional plant protection products. Basic substances are defined as compounds that are not predominantly used as plant protection products but may be useful in crop protection. They have no toxicological concerns and do not cause adverse effects on humans, animals, or the environment [9]. Interestingly, ‘foodstuff’ substances (as defined by Regulation (EC) No. 178/2002) are intrinsically considered basic substances [15]. Basic substances have no residue limits, and usually no pre-harvest interval [16]. Also, since they are not currently placed on the market as plant protection products, they are not considered in the Harmonized Risk Indicator 1 calculation that is used in the EU for highlighting the trends in the risks associated with the use of pesticides [17]. European basic substances partially overlap with the American “Generally Recognized as Safe” (GRAS) substances, which are approved for use in food products as preservatives [18].
2. Activity of Approved Basic Substances against Fungal Diseases
Fungicides have been the top-selling group of plant protection products in the EU for a long time. Three countries, namely Spain, France, and Italy, make up around 62% of the total volume of pesticides (330 thousand tons) sold annually between the years 2011 and 2020 [19]. Interestingly, these countries also have the highest surface area dedicated to viticulture, which accounts for 75% of the 3.2 million hectares under vines [20]. This is due to the fact that many fungicide sprays are applied each growing season to the grapevine crop (Vitis vinifera) to manage three major fungal diseases: downy and powdery mildews, and grey mold [21]. The major fungicide markets are fruit and vegetables, cereals, grapevines, and potatoes, which account for about 60% of the global fungicide market (https://www.apsnet.org/edcenter/apsnetfeatures/Pages/Fungicides.aspx, accessed on 3 May 2023). In the following paragraphs, information on the results achieved through basic substance applications to control the important diseases of grapevines, potatoes, and fruits, in general, will be provided, taking into consideration that pre-harvest treatments also affect the post-harvest control of the pathogens (Table 1).2.1. Grapevine
2.1.1. Grapevine Downy Mildew
European grapevine exhibits a high level of susceptibility to grapevine downy mildew, caused by the oomycete Plasmopara viticola [22]. To prevent infections and the consequent production loss, several treatments with chemical fungicides are needed during the season under both organic and integrated pest management systems [22]. This results in negative consequences for the environment and risks for human health. Copper is the most widely applied plant protection product acting against grapevine downy mildew, although the Regulation (EU) 2018/1981 restricted the quantities allowed and classified this heavy metal as an active substance candidate for substitution [23]. Copper fungicides are fundamental for organic productions, where the use of synthetic curative compounds is not allowed, but also play a central role in integrated pest management to limit the outbreak of resistant strains. This situation encouraged the search for alternative tools to protect plants from P. viticola. Chitosan [24], biocontrol agents [25], aptamers [26], hydrolyzed proteins [27], laminarin [28], stilbenes [29], and other plant extracts [30][31][32] showed promising results under in vitro or in vivo experiments. Among these alternatives, basic substances could present a good opportunity. Chitosan, Equisetum arvense (horsetail), sucrose, Salix spp. cortex, lecithin, fructose, and nettle (Urtica spp.) are the basic substances that may exhibit effectiveness against grapevine downy mildew [16], especially when integrated into reduced copper strategies. The field application of chitosan hydrochloride alone showed promising results in plot trials, under different environmental conditions, and even under the presence of a high disease pressure [33][34][35]. This biopolymer is obtained from chitin deacetylation, and it can perform eliciting, antimicrobial, and film-forming activities once applied on plant tissues [33]. Results obtained with chitosan individual treatments were similar to those obtained with a conventional application of copper, showing disease reductions compared to the untreated control, which in some cases exceeded 95% on leaves and 80% on grape bunches [33][34]. Nevertheless, chitosan effectiveness against grapevine downy mildew is strictly linked to two main factors: volume of applications and active ingredient concentrations. To best perform the triple mode of action on plant tissues, a good wetting of the canopy is required and the standard spraying volume for grapevine (1000 L/ha of water) is recommended. Application of 0.8% chitosan has been found to perform better than copper hydroxide in seasons characterized by frequent rainfall and high disease pressure [34]. The 0.5% of active ingredient does not usually show significant differences compared to the 0.8% in terms of their efficacy, as well as being less expensive for the growers. Furthermore, treatments with high concentrations of chitosan for the whole season can induce undesired collateral physiological responses in vines, such as reduced growth and leaf area [34]. In addition to being dangerous for humans and ecosystems [36], copper residues on the berries affect the wine quality, reducing the concentration of several amino acids in the must [37]. Unlike copper, chitosan and other natural compounds, such as laminarin, have lower impacts on the final product quality [37]. Results obtained in the past years have suggested chitosan as a promising tool to support or eventually replace copper for grapevine downy mildew management. Copper and chitosan could even coexist to begin with, for example with alternating or combined treatments, even if validations on a commercial scale for these strategies are needed. According to the data available in the literature, no copper could be applied under instances of low disease pressure, while a valid strategy for difficult seasons could be to apply copper until flowering (in the period of higher susceptibility) and then replace it with chitosan. In this way, it could be possible to reduce the quantities of copper distributed per year on the one hand and the costs of chitosan on the other hand. Indeed, the main limitations regarding chitosan diffusion so far are represented by its cost and the lack of operational knowledge. It will be important to invest in new formulations and to investigate the miscibility of this biopolymer with other plant protection products, since farmers are used to simultaneously applying several compounds so as to target different pests within a single treatment.2.1.2. Grey Mold on Table Grape
Grey mold is a globally widespread and economically relevant disease of grapes caused by the second most important phytopathogenic fungus, Botrytis cinerea [38]. This broad host range pathogen affects several crops, both under pre- and post-harvest. B. cinerea can survive and develop in vineyards as both a necrotrophic pathogen and a saprophyte [39][40]. Grey mold can result from multiple infection pathways on ripening grape berries, including latent infections established during blooming, direct berry infection due to airborne conidia, and berry-to-berry infection caused by mycelium originating from previously infected berries (nesting path) within the cluster [41][42], which spread according to a nesting path. Although B. cinerea causes about 30% of latent infections [43], it is difficult to precisely estimate the global losses due to its broad host range and specific missing statistics. New Zealand recorded costs due to grey mold direct crop losses and grey mold control measures of up to NZD 5000/ha and NZD 1500/ha, respectively, in growing seasons favorable for disease development [39]. In Australia, Chile, and South Africa, grey mold is the main cause of wine and table grape losses, from the vineyard to the retail outlet, entailing profit reductions of AUD 52 million/year, USD 22.4 million/year, and ZAR 25 million/year, respectively [38]. Chemical fungicides are the most important control means available, although fungicide resistance is an increasing issue for this pathogen [44][45][46]. Fungicide-resistant phenotypes were detected in B. cinerea populations in table grape vineyards in California, with genotypic resistance against boscalid, cyprodinil, fenhexamid, and pyraclostrobin in 95%, 85%, 23%, and 14% of tested isolates, respectively [47]. Differences in the fungicide resistance profile of B. cinerea may be due to the species/groups included within the complex; as an example of biodiversity, in the pomegranate fruit, B. cinerea, B. pseudocinerea, and Botrytis group S were the etiological agents of grey mold [48][49]. Currently, latent infections caused by Botrytis spp. are completely prevented in storage through the use of SO2-generating pads [43], although these entail adverse effects on food, humans, and the environment (i.e., phytotoxicity, development of antimicrobial resistance, allergy, pollution, etc.) [50], and cannot be applied to organic table grapes. This encourages the set-up of new, safer, more effective, and cheaper alternative control means and strategies. Basic substances, such as salts and chitosan, and potential basic substances can be a promising alternative to chemical fungicides for grey mold management [51][52]. Chitosan treatments have been shown to significantly reduce disease incidence both in the field and after harvest. It indirectly enhances the activity of the key plant enzymes involved in disease resistance, such as superoxide dismutase, peroxidase, catalase, and ascorbate peroxidase, that damage the mycelial structures of Botrytis spp. and reduce pathogen development [53][54][55]. Grey mold on table grapes is a disease affecting clusters both in the field and during the post-harvest phases. Unfortunately, since B. cinerea affects grapes more heavily during the post-harvest phases, most of the papers that are available on this subject concern the disease development after harvest, and very few concern pre-harvest evaluations [56]. Grey mold protection starts during the grapevine growing season following a well-established scheme, in which four applications of fungicides are carried out under the following specific phenological stages: berry set, pre-bunch closure, veraison, and 1–3 weeks before harvest. This strategy, that is mandatory to avoid latent infections during the growing season [42], was also adopted for chitosan and other alternative control means. In field treatments on table grapes, 1% chitosan demonstrated the same ability to protect grapes from grey mold as the strategy based on synthetic fungicide application. In an integrated program lasting two years, chitosan-treated “Chardonnay” wine grapes exhibited a degree of disease severity at harvest that was more than halved compared to the untreated control and was as effective as the synthetic fungicide program [57]. Chitosan has also been combined with active antimicrobial substances, such as essential oils, and applied as pre-harvest treatments [58] or as post-harvest coatings to improve the preservation of qualitative parameters and reduce the product losses caused by Botrytis spp. A possible evolution in the application of chitosan is its formulation as nanoparticles, which in preliminary trials behaved better than standard formulations [55]. This basic substance has been used formulated as a chitosan/silica nanocomposite-based compound, which reduced conidial germination and germ tube elongation, affecting the development of grey mold on the grapes [55]. Various substances have been tested at pre-harvest in combination with chitosan, such as with chitosan added or complexed with salicylic acid. In particular, the CTS-g-SA complex improved fruit physiology (transpiration and respiration rates), qualitative parameters (soluble solids, titratable acidity, and total phenolic content), and defense mechanisms involving the control of disease incidence [50]. Among the other basic substances currently approved, sodium bicarbonate has been studied worldwide, leading to results that are, in many cases, not different, if not better, than the synthetic fungicides [59]. When applied before harvest, sodium bicarbonate showed a significant reduction in botrytis storage rots in both small-scale and large-scale tests. In large-scale trials simulating practical commercial conditions adopted in Southern Italy, two salt applications (at 30 and 90 days before harvest) of sodium bicarbonate significantly reduced grey mold from 23% (untreated control) to 12% [60]. Among these basic substances, sodium bicarbonate may represent one of the most useful and effective compounds, considering that it is easy to find on the market, is very cheap, has a broad spectrum of activity against a variety of pathogens, is well accepted by consumers and operators, and has an acceptable environmental profile.2.2. Potato Leaf Diseases
The potato (Solanum tuberosum) is one of the most important vegetable crops in the world. It belongs to the family Solanaceae and is an important starchy food crop. Potato plants are subjected to numerous diseases wherever the crop is grown. Among the approved basic substances, there are some that have the potential to limit the early and late blight of potatoes by spraying or dipping tubers before sowing. Early blight of potatoes caused by Alternaria solani and Alternaria alternata, and late blight caused by the oomycete Phytophthora infestans are major causes of concern in potato production. This problem is particularly important in organic farming, where synthetic fungicides are prohibited. Therefore, a necessary condition in the organic cultivation of potatoes is the timely implementation of treatments and adherence to the rules of agricultural technology regarding the appropriate variety and crop rotation. Alternaria spp. are air- and soil-borne organisms that cause disease on foliage (leaf blight), stems (collar rot), and tubers (tuber rot), resulting in severe damage during all stages of plant development [61]. This disease causes losses in crop productivity in the field and in tuber quality during storage. The average annual yield loss of potatoes due to this disease is approximately 79% of the total production, depending on the nature of the disease, weather conditions, and the type of variety grown [62]. It can destroy foliage prematurely and in a short time, reducing production, while the tuber infections associated with rots can cause significant crop losses during storage. It is considered one of the most destructive crop pathogens threatening global food security. In organic potato production, late blight can cause severe losses in the potato yield and quality. Currently, in organic farming it can only be effectively controlled using copper fungicides. However, some countries prohibit copper use in organic farming based on their national laws, as the harmfulness of copper in ecosystems is still being debated. Studies aiming at the reduction in copper usage and testing of potential basic substances against late blight for organic farming are needed. Currently, basic substances, such as extracts from the bulb of the onion crop (Allium cepa) and horsetail, have been proposed as protective treatments against early blight. In the case of P. infestans, chitosan hydrochloride is most frequently mentioned as an elicitor, along with nettle extracts, lecithins, and dried horsetail [16]. However, the use of these substances in the field has not often been demonstrated. The application of elicitors and botanical fungicides, beneficial microorganisms, and basic substances should be a combination of compounds and microorganisms with different modes of action, beginning at the early stages of the potato plant’s growth [63][64][65][66]. This strategy of minimizing the risk to the ecosystem is a global trend, especially in the EU, where the policy of greening agriculture is being promoted. Chitosan significantly inhibits the mycelial growth and in vitro spore germination of P. infestans, induces resistance to the pathogen in potato pieces and leaves [67], and forms a mechanical barrier to the pathogen penetration [68][69]. It also has a synergistic effect with plant protection products, making it a potential way to reduce the use of chemical plant protection products. In field conditions, the use of chitosan can stimulate plants to defend themselves, which in turn contributes to limiting the harmful effects of potato disease symptoms. Late blight epidemics were delayed on plots that received eight sprays of 0.1% chitosan [70] and provided 60% protection against late blight by mixing 4% chitosan with a plant elicitor [71]. Some late blight reducing potential for 0.4% chitosan was found in field tests performed in Germany on the cultivars Nicola and Ditta [72]. Field tests confirmed some of the major results coming from the lab and growth chamber assays. Most effects were only visible during the early phases of the disease, when plants were still vigorous, but might have been more pronounced under a different infection regime with an earlier onset of the disease. Both in full-field tests, chitosan (0.4%) and the copper fungicide, and in a small plot trial, chitosan (0.4%) accompanied with the horsetail and liquorice products seemed to be able to cause some degree of disease reduction, even under an extremely late infection regime [72]. Good results with a low-level copper formulation (copper sulfate pentahydrate), together with chitosan as an adhesive substance to increase rain fastness, were also obtained [73]. In recent years, practical applications of chitosan were tested against P. infestans in in vivo experiments under outdoor conditions [74]. This experiment showed that chitosan is very effective against P. infestans. An average damage of over 76% was observed in the control plants. In the treated variants with 1–4 applications of chitosan, the final damage to the plants ranged from 48% to 0.5%. Expressed as values of the final inhibitory protective effect, a single application of a 0.4% solution of chitosan provided an inhibitory effect of 37%. In cases where chitosan was applied four times, an inhibitory effect of up to 99.3% was demonstrated [74]. The newest study also confirmed that chitosan can be applied as the nano compound. The bioactivity and absorbency of elicitors are critical factors that limit the large-scale field application. A star polymer was constructed to deliver the nano-sized (particle size from 144.61 nm to 17.40 nm in an aqueous solution) chitosan to enhance the control effects against potato late blight [75]. As basic substances, lecithins have fungicidal activity due to their inhibition of the fungal hypha penetration into the plant cells. Unlike chitosan, lecithins are not fully soluble in water. Since chitosan would be the first choice in most situations when looking for a fungicide among the basic substances, the next option for controlling oomycetes may be to use lecithins in combination with chitosan in joint field treatments (https://eutrema.co.uk/basic-substances-what-are-they-and-how-can-they-used-for-pest-and-disease-control-on-farms/, accessed on 20 January 2023). The fatty acids present in the lecithins could act more positively via plant defense stimulation, rather than through a toxic effect. In fact, linolenic acid and its precursor linoleic acid, both present in soy lecithin, are the precursors of a wide variety of oxylipins and the plant hormone jasmonic acid, which actively participate in plant defenses [76]. Lecithins have not been studied in field trials. In field tests, the application of 12 kg/hL horsetail macerate showed effectiveness in protecting the tomato crop (Solanum lycopersicum) from late blight that was analogous to the copper-based treatments [77]. Nettle slurry (Urtica dioica), used as a foliar fertilizer in different doses, alone or in combination with horsetail, had no significant effects on the yield, chlorophyll content, or the presence of pests and diseases in organic potato crops [78]. Conversely, the methanolic leaf extracts of nettle slurry and broad-leaf hopbush (Dodonaea viscosa) demonstrated a strong antifungal efficacy against A. alternata. Among the many polyphenolic compounds that were detected in the HPLC of the extract, coumaric acid, caffeic acid, ferulic acid, and α-tocopherol showed potent in vitro fungicidal activity against A. alternata, either applied alone or in combination at low concentrations [79]. In Romania, 2.2% and 3.3% water solutions of the onion crop showed significant protection against A. solani in potato fields [80]. Dry extracts of onion (concentration 20.0 mg/mL) showed antifungal activity against A. alternata and P. infestans. In particular, red onion extracts showed a higher efficacy in inhibiting A. alternata than white onion extracts, which showed no efficacy. This result is surprising, considering that both extracts have a similar amount of quercetin, an antioxidant with antifungal activity. Evidently, other components of these extracts are responsible for the A. alternata inhibition [81].2.3. Pre-Harvest Treatment Affecting the Post-Harvest Diseases of Fruits
Fruit-bearing plants may be infected in the field before or during their harvest, providing inoculum for post-harvest decay following their harvest [82]. The accumulation and/or survival of the inoculum can be prevented through pre-harvest treatments [83]. Basic substances may provide environmentally friendly alternatives to pre-harvest fungicide application to prevent the post-harvest decay of fruits and vegetables, although there is limited information about their efficacy. Chitosan hydrochloride and chitosan are the most widely studied basic substances in pre-harvest application, either alone or in combination. Similarly to what has been observed for table grapes [84][85][86][87], the pre-harvest application of 0.2–1% chitosan was effective against grey mold latent infection and the decay of strawberries (Fragaria x ananassa and Fragaria chiloensis) [88][89][90][91][92]. The pre-harvest application of 1% chitosan was effective against the grey mold and brown rot of sweet cherries (Prunus avium) and date palm fruits (Phoenix dactylifera) [93][94]. The soft rot of kiwifruit (Actinidia deliciosa) caused by Botryosphaeria dothidea and Phomopsis sp. was also reduced following a chitosan-containing spray [95]. This basic substance was effective in being able to reduce the A. alternata-related decay of apricots (Prunus armeniaca) [96][97] and in the decay of peaches (Prunus persica) [98][99]. The pre-harvest treatment of jujube (Zizyphus jujuba) and tomato plantations with 0.3–1 g/L chitosan was also effective in being able to reduce the decay of these harvested fruits [100][101]. However, in the case of raspberries (Rubus idaeus), 1% or 2% chitosan was only effective in reducing the decay of these fruits during their storage [102].Table 1. List of the basic substances which effectively protected the crops described herein from specific diseases.
| Crop (Species) | Disease (Pathogen) | Basic Substance | Reference |
|---|---|---|---|
| Grapevine (Vitis vinifera) | Downy mildew (Plasmopara viticola) | Chitosan | [34][35] |
| Botrytis bunch rot (Botrytis cinerea) | Chitosan | [58] | |
| Potato (Solanum tuberosum) | Early blight (Alternaria alternata) | Nettle slurry (Urtica dioica) and broad-leaf hopbush (Dodonaea viscosa) methanolic extracts | [79] |
| Early blight (Alternaria solani) | Water solutions of Allium cepa | [80] | |
| Late blight (Phytophthora infestans) | Chitosan | [70][71][72][74] | |
| Strawberry (Fragaria x ananassa and Fragaria chiloensis) | Grey mold (Botrytis cinerea) | Chitosan | [83][88][89][90] |
| Sweet cherry (Prunus avium) | Storage decay | Chitosan | [91] |
| Botrytis rot (B. cinerea) | Sodium bicarbonate salts | [101] | |
| Date palm fruit (Phoenix dactylifera) | Storage decay | Chitosan | [92] |
| Kiwifruit (Actinidia deliciosa) | Soft rot (Botryosphaeria dothidea and Phomopsis sp.) | Chitosan | [93] |
| Apricot (Prunus armeniaca) | Decay (A. alternata) | Chitosan | [94][95] |
| Peach (Prunus persica) | Decay (A. alternata) | Chitosan | [96][97] |
| Jujube (Zizyphus jujuba) | Storage decay | Chitosan | [98] |
| Tomato (Solanum lycopersicum) | Storage decay | Chitosan | [99] |
| Pear (Pyrus communis) | Storage decay | Onion (Allium cepa) extract | [102] |
3. Activity of Approved Basic Substances against Insects
Based on the analysis of data obtained from Scopus, over 3000 articles regarding eco-friendly natural product pesticides in crop protection have been published in the Agricultural and Biological Sciences sector, with an increasing trend in the last 30 years and a peak of 271 papers published in 2021. However, a search on Scopus using the keywords ‘pest control’ and ‘basic substance’ yielded only 11 scientific articles published since 2015, with the first article published in 2015 [15]. According to the EU Regulation (EC) 1107/2009, among the twenty-four basic substances permitted for plant protection use, eight are approved as insecticides (nettle, sodium chloride, L-cysteine, sucrose, and fructose), physical barriers (talc E553B), attractants (diammonium phosphate), or repellents (onion oil). In the following paragraphs, the literature concerning the field application of these eight basic substances will be described, along with their modes of action (Figure 21).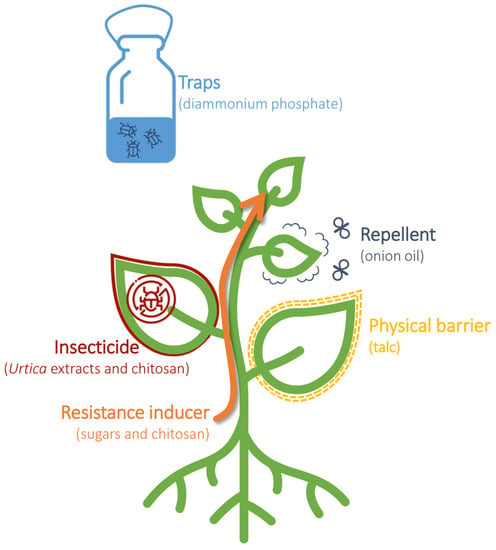
Figure 21. Utilization of basic substances in the management of plant pests. Basic substances can be used as insecticides, resistance inducers, physical barriers, repellents, and traps to control and manage plant pests.
3.1. Nettle
The extracts of nettle, commonly known as a foodstuff and medicine, are traditionally used by farmers who claim a significant reduction in the aphid and Coleoptera presence [108][109][110]. Searching the keywords “Urtica” and “pest management” on the Scopus database resulted in the retrieval of more than 500 papers that were published in the Agricultural and Biological Sciences sector, demonstrating that nettle is one of the most studied basic substances for pest management purposes. Nettle can be used as a fermented aqueous extract in spray applications against different aphid species, such as Myzus persicae, Macrosiphum rosae, Eriosoma lanigerum, and Panaphis juglandi, to protect fruit trees (Malus domestica, Prunus spp.), elder trees, beans (e.g., Phaseolus vulgaris), leafy vegetables (Lactuca sativa, Brassica oleracea), Rosa spp., and Spiraea spp. With a population density reduction of more than 30%, nettle extracts can also be used on Brassicaceae crops against the flea beetle, Phyllotreta nemorum, and the diamond back moth, Plutella xylostella, as well as on apple and pear trees against the codling moth, Cydia pomonella. In field trials, nettle slurry fermented extract showed a repellent activity towards Hyalopterus pruni and P. juglandi [111][112], but not against Aphis spiraephaga [111], suggesting that the efficacy of the nettle slurry extract against aphids is species-dependent. Under controlled conditions, Urtica urens water extract effectively limited the fertility of M. persicae, slightly reducing the increase of its population (by 20% on average), while no negative effects were registered on its natural enemy, Macrolophus pygmaeus [108]. Furthermore, the nettle extract used in combination with other biorational insecticides could improve the efficacy against aphid pests [108]. Nettle extracts can also be used to control the mites Tetranychus urticae on beans and Tetranychus telarius on grapevines. Repellent, acaricidal, and antifeedant activities of the nettle extracts against T. urticae, one of the economically most important pests in a wide range of outdoor and protected crops worldwide, have been reported [113][114]. Less information is available in the literature regarding the effect of nettle extracts on T. telarius. In this scenario, extracts from several plants proved to exert insecticidal or miticidal activity against vegetables and stored-product pests [115][116][117][118], that, in some cases, were comparable to those achieved using chemical insecticides (e.g., synthetic pyrethroid) [119], and could suggest potential basic substances as alternatives to synthetic chemical insecticides in crop protection.3.2. Sucrose and Fructose
Sucrose and fructose are involved in the phenomenon of “Sweet Immunity”, according to which the sugar metabolism and signaling influence the plant immunity networks [120][121][122]. The quantities and ratios of three soluble carbohydrates (sucrose, D-fructose, and glucose) and three sugar alcohols (sorbitol, quebrachitol, and myo-inositol) of apple tree surfaces play a role in the trees’ resistance, as they influence the host preference, egg laying, and the behavior of the neonate larvae of Cydia pomonella [123][124][125][126]. Recent studies demonstrated that sucrose, in micro-dose foliar applications, can induce partial resistance via antixenosis to C. pomonella egg laying [127]. Moreover, the spraying of glucose or fructose significantly reduced the percentage of damaged fruits by C. pomonella by 70% compared to the untreated control, with an effectiveness comparable with the spraying of the chemical insecticide deltamethrin [128][129]. In field trials, sucrose treatment was found to be as efficient as thiacloprid treatment in the reduction of damage by C. pomonella. Furthermore, synergistic effects were found when sucrose was combined with the thiacloprid insecticide [127], and between fructose and organophosphorus or insect growth regulator insecticides against the codling moth [130]. The quantities and ratios of soluble carbohydrates and on the leaf surface could also influence the egg-laying preferences of Ostrinia nubilalis on maize hybrids [131][132][133][134][135]. A study contributed to explore the efficacy of sucrose and fructose, used alone or in combination with natural pyrethrum, against O. nubilalis and Scaphoideus titanus [136]. The authors found that the application of sucrose associated with fructose provided the best efficacy in reducing the number of corn borer larvae per plant with a 23% efficacy. In the case of S. titanus, sucrose seemed to increase the action of natural pyrethrum, whilst the fructose showed the same efficacy as the natural pyrethrum. Finally, sugars could be also interesting as components of commercial biopesticides due to the phagostimulant activity for a more effective ingestion by larvae [137]. These studies demonstrate a promising alternative to conventional crop protection tools [138] and pave the way for the development of eco-friendly control strategies using the new concept of “Sweet Immunity” induction.3.3. Talc
Magnesium hydrogen metasilicate, known by the common name of talc, is approved as a basic substance to be used in outdoor applications on grapevines and fruit orchards to act as a physical barrier towards insects and mites, like Cacopsylla pyri, Cacopsylla fulguralis, Drosophila suzukii, Panonychus ulmi, and Bactrocera oleae [139][140][141]. Nowadays, the research interest in the use of inert dusts and their potential role in agriculture to manage diseases and protect crops from insect pests is increasing [142][143]. Among mineral products, natural zeolites, a broad range of crystalline hydrated aluminosilicates [144][145], could represent potential basic substances. Thanks to their physical and chemical properties and uses, the Codex Alimentarius Commission (1999) endorsed their use for pest control in food commodities and listed zeolites as granted substances in the organic food production and plant protection [146]. The insecticidal activity of zeolites towards stored-product insect pests, such as Sitophilus zeamais, Rhyzopertha dominica, Sitophilus oryzae, Tribolium castaneum, Lasioderma serricorne, Tribolium confusum, Callosobruchus maculatus, and Meligethes spp., was intensively reported [143][147][148][149][150][151][152][153][154][155][156]. In addition, the 40% reduction in the oviposition rates of B. oleae females due to zeolite applications was observed [157].3.4. Diammonium Phosphate
Plant volatile compounds are involved in the host-finding process and oviposition site selection by insects [158][159]. The efficiency of traps used in indirect (e.g., monitoring) and direct (e.g., mass trapping, attract, and kill) semiochemical-based control tools can be improved significantly through the addition of certain food attractants [160]. Ammonia-releasing substances play an important role in both sexes of fruit fly attraction to food sources [161][162][163]. Thus, ammonia bait traps are currently used for monitoring fruit fly populations [164]. The use of diammonium phosphate is permitted to bait one trap per tree in orchards, including Prunus spp., Citrus spp., and olives (Olea europaea), to enable the massive capture of adults of the above-mentioned insect species. In this context, fruit fly pheromones added to food attractants, such as diammonium phosphate, are efficient for the monitoring and mass trapping of C. capitata, B. cucurbitae, and B. dorsalis [165], and are commonly used in the monitoring of B. oleae in most olive-growing countries of the Mediterranean basin [166].3.5. Onion Oil
Onion oil obtained from A. cepa is authorized as a basic substance due to its repellent and scent masking activity against the carrot root fly, Psila rosae [167]. Dispensers of undiluted oil placed in the field are able to disorient adult flies which cannot find its host plant. Dispensers are filled with onion oil alone or with ethylene vinyl acetate granules that are able to improve the release of vapor.3.6. Chitosan
Among these basic substances, chitosan stimulates the defense system of crops against several classes of pathogens, including fungi, viruses, bacteria, and phytoplasmas [33], and its use as an elicitor of the crop’s self-defense mechanisms has also been approved. Chitosan also exhibits a strong level of insecticidal activity against various insect pests [168]. The insecticidal activity of chitosan and its derivatives was demonstrated against the lepidopterans Spodoptera littoralis [70][169], Helicoverpa armigera, and P. xylostella, and the aphids Aphis gossypii, Metopolophium dirhodum, H. pruni, Rhopalosiphum padi, Sitobium avenae, and M. persicae [170][171]. The mortality of six types of aphids generally ranged between 60% and 80%, with a peak of 99.7%. Furthermore, recent studies showed that a new chitosan derivative, named avermectin-grafted-N,O-carboxymethyl chitosan (NOCC), showed an excellent insecticidal and acaricidal activity against Aphis fabae, Nilaparvata lugens, and Tetranychus cinnabarinus [172].References
- Lucas, J.A. Foresight Project on Global Food and Farming Futures: Advances in Plant Disease and Pest Management. J. Agric. Sci. 2011, 149, 91–114.
- Carvalho, F.P. Pesticides, Environment, and Food Safety. Food Energy Secur. 2017, 6, 48–60.
- Lykogianni, M.; Bempelou, E.; Karamaouna, F.; Aliferis, K.A. Do Pesticides Promote or Hinder Sustainability in Agriculture? The Challenge of Sustainable Use of Pesticides in Modern Agriculture. Sci. Total Environ. 2021, 795, 148625.
- Jacquet, F.; Jeuffroy, M.H.; Jouan, J.; Le Cadre, E.; Litrico, I.; Malausa, T.; Reboud, X.; Huyghe, C. Pesticide-Free Agriculture as a New Paradigm for Research. Agron. Sustain. Dev. 2022, 42, 8.
- Pierlot, F.; Marks-Perreau, J.; Soulé, E.; Keichinger, O.; Bedos, C.; Prevost, L.; Van Dijk, P.; Bockstaller, C. An Indicator to Assess Risks on Water and Air of Pesticide Spraying in Crop Fields. Sci. Total Environ. 2023, 870, 161000.
- van den Berg, F.; Jacobs, C.M.J.; Butler Ellis, M.C.; Spanoghe, P.; Doan Ngoc, K.; Fragkoulis, G. Modelling Exposure of Workers, Residents and Bystanders to Vapour of Plant Protection Products after Application to Crops. Sci. Total Environ. 2016, 573, 1010–1020.
- Tudi, M.; Li, H.; Li, H.; Wang, L.; Lyu, J.; Yang, L.; Tong, S.; Yu, Q.J.; Ruan, H.D.; Atabila, A.; et al. Exposure Routes and Health Risks Associated with Pesticide Application. Toxics 2022, 10, 335.
- Kenko, D.B.N.; Ngameni, N.T.; Awo, M.E.; Njikam, N.A.; Dzemo, W.D. Does Pesticide Use in Agriculture Present a Risk to the Terrestrial Biota? Sci. Total Environ. 2023, 861, 160715.
- European Parliament Regulation (EC). No 1107/2009 of the European Parliament and of the Council of 21 October 2009 Concerning the Placing of Plant Protection Products on the Market and Repealing Council Directives 79/117/EEC and 91/414/EEC. Off. J. Eur. Union 2009, L309, 1–50.
- Hillocks, R.J. Farming with Fewer Pesticides: EU Pesticide Review and Resulting Challenges for UK Agriculture. Crop. Prot. 2012, 31, 85–93.
- Wilson, C.; Tisdell, C. Why Farmers Continue to Use Pesticides despite Environmental, Health and Sustainability Costs. Ecol. Econ. 2001, 39, 449–462.
- Gehen, S.; Corvaro, M.; Jones, J.; Ma, M.; Yang, Q. Challenges and Opportunities in the Global Regulation of Crop Protection Products. Org. Process. Res. Dev. 2019, 23, 2225–2233.
- McDougall, P. The Cost of New Agrochemical Product Discovery, Development and Registration in 1995, 2000, 2005–2008 and 2010–2014. Available online: https://croplife.org/wp-content/uploads/2016/04/Cost-of-CP-report-FINAL.pdf (accessed on 26 July 2023).
- Hawkins, N.J.; Bass, C.; Dixon, A.; Neve, P. The Evolutionary Origins of Pesticide Resistance. Biol. Rev. 2019, 94, 135–155.
- Marchand, P.A. Basic Substances: An Opportunity for Approval of Low-Concern Substances under EU Pesticide Regulation. Pest Manag. Sci. 2015, 71, 1197–1200.
- Romanazzi, G.; Orçonneau, Y.; Moumni, M.; Davillerd, Y.; Marchand, P.A. Basic Substances, a Sustainable Tool to Complement and Eventually Replace Synthetic Pesticides in the Management of Pre and Postharvest Diseases: Reviewed Instructions for Users. Molecules 2022, 27, 3484.
- European Commission. Commission Directive (EU) 2019/782 of 15 May 2019 Amending Directive 2009/128/EC of the European Parliament and of the Council as Regards the Establishment of Harmonized Risk Indicators. Off. J. Eur. Union 2019, L127, 4–10.
- Sun, C.; Zhu, C.; Tang, Y.; Ren, D.; Cai, Y.; Zhou, G.; Wang, Y.; Xu, L.; Zhu, P. Inhibition of Botrytis cinerea and Control of Gray Mold on Table Grapes by Calcium Propionate. Food Qual. Saf. 2021, 5, 1–12.
- Eurostat. Key Figures on the European Food Chain—2021 Edition; Publications Office of the European Union: Bietlot, Belgium, 2021; ISBN 9789276415152.
- Eurostat. Agriculture, Forestry and Fishery Statistics—2020 Edition; Publications Office of the European Union: Bietlot, Belgium, 2020; ISBN 978-92-76-21521-9.
- Bois, B.; Zito, S.; Calonnec, A.; Ollat, N. Climate vs Grapevine Pests and Diseases Worldwide: The First Results of a Global Survey. J. Int. Sci. Vigne Vin 2017, 51, 133–139.
- Gessler, C.; Pertot, I.; Perazzolli, M. Plasmopara viticola: A Review of Knowledge on Downy Mildew of Grapevine and Effective Disease Management. Phytopathol. Mediterr. 2011, 50, 3–44.
- European Commission. Commission Implementing Regulation (EU) 2018/1981 of 13 December 2018 Renewing the Approval of the Active Substances Copper Compounds, as Candidates for Substitution, in Accordance with Regulation (EC) No 1107/2009 of the European Parliament and of the Co. Off. J. Eur. Union 2018, L317, 16–20.
- Maia, A.J.; Leite, C.D.; Botelho, R.V.; Faria, C.M.D.R.; Machado, D. Quitosana Como Opção de Controle Do Míldio Para Viticultura Sustentável. Semin. Cienc. Agrar. 2012, 33, 2519–2530.
- Puopolo, G.; Giovannini, O.; Pertot, I. Lysobacter capsici AZ78 Can Be Combined with Copper to Effectively Control Plasmopara Viticola on Grapevine. Microbiol. Res. 2014, 169, 633–642.
- Rosa, S.; Pesaresi, P.; Mizzotti, C.; Bulone, V.; Mezzetti, B.; Baraldi, E.; Masiero, S. Game-Changing Alternatives to Conventional Fungicides: Small RNAs and Short Peptides. Trends Biotechnol. 2022, 40, 320–337.
- Lachhab, N.; Sanzani, S.M.; Adrian, M.; Chiltz, A.; Balacey, S.; Boselli, M.; Ippolito, A.; Poinssot, B. Soybean and Casein Hydrolysates Induce Grapevine Immune Responses and Resistance against Plasmopara viticola. Front. Plant. Sci. 2014, 5, 716.
- Aziz, A.; Poinssot, B.; Daire, X.; Adrian, M.; Bézier, A.; Lambert, B.; Joubert, J.M.; Pugin, A. Laminarin Elicits Defense Responses in Grapevine and Induces Protection Against Botrytis cinerea and Plasmopara viticola. Mol. Plant-Microbe Interact. 2003, 16, 1118–1128.
- Gabaston, J.; Cantos-Villar, E.; Biais, B.; Waffo-Teguo, P.; Renouf, E.; Corio-Costet, M.F.; Richard, T.; Mérillon, J.M. Stilbenes from Vitis vinifera L. Waste: A Sustainable Tool for Controlling Plasmopara viticola. J. Agric. Food Chem. 2017, 65, 2711–2718.
- Dagostin, S.; Formolo, T.; Giovannini, O.; Pertot, I.; Schmitt, A. Salvia Officinalis Extract Can Protect Grapevine Against Plasmopara viticola. Plant Dis. 2010, 94, 575–580.
- La Torre, A.; Mandalà, C.; Pezza, L.; Caradonia, F.; Battaglia, V. Evaluation of Essential Plant Oils for the Control of Plasmopara viticola. J. Essent. Oil Res. 2014, 26, 282–291.
- Rienth, M.; Crovadore, J.; Ghaffari, S.; Lefort, F. Oregano Essential Oil Vapour Prevents Plasmopara viticola Infection in Grapevine (Vitis Vinifera) and Primes Plant Immunity Mechanisms. PLoS ONE 2019, 14, e0222854.
- Romanazzi, G.; Feliziani, E.; Sivakumar, D. Chitosan, a Biopolymer with Triple Action on Postharvest Decay of Fruit and Vegetables: Eliciting, Antimicrobial and Film-Forming Properties. Front. Microbiol. 2018, 9, 2745.
- Romanazzi, G.; Mancini, V.; Feliziani, E.; Servili, A.; Endeshaw, S.; Neri, D. Impact of Alternative Fungicides on Grape Downy Mildew Control and Vine Growth and Development. Plant Dis. 2016, 100, 739–748.
- Romanazzi, G.; Mancini, V.; Foglia, R.; Marcolini, D.; Kavari, M.; Piancatelli, S. Use of Chitosan and Other Natural Compounds Alone or in Different Strategies with Copper Hydroxide for Control of Grapevine Downy Mildew. Plant Dis. 2021, 105, 3261–3268.
- Rehman, M.; Liu, L.; Wang, Q.; Saleem, M.H.; Bashir, S.; Ullah, S.; Peng, D. Copper Environmental Toxicology, Recent Advances, and Future Outlook: A Review. Environ. Sci. Pollut. Res. 2019, 26, 18003–18016.
- Garde-Cerdán, T.; Mancini, V.; Carrasco-Quiroz, M.; Servili, A.; Gutiérrez-Gamboa, G.; Foglia, R.; Pérez-Álvarez, E.P.; Romanazzi, G. Chitosan and Laminarin as Alternatives to Copper for Plasmopara viticola Control: Effect on Grape Amino Acid. J. Agric. Food Chem. 2017, 65, 7379–7386.
- Dean, R.; Van Kan, J.A.L.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The Top 10 Fungal Pathogens in Molecular Plant Pathology. Mol. Plant Pathol. 2012, 13, 414–430.
- Mundy, D.C.; Elmer, P.; Wood, P.; Agnew, R. A Review of Cultural Practices for Botrytis Bunch Rot Management in New Zealand Vineyards. Plants 2022, 11, 3004.
- Plesken, C.; Pattar, P.; Reiss, B.; Noor, Z.N.; Zhang, L.; Klug, K.; Huettel, B.; Hahn, M. Genetic Diversity of Botrytis cinerea Revealed by Multilocus Sequencing, and Identification of B. Cinerea Populations Showing Genetic Isolation and Distinct Host Adaptation. Front. Plant Sci. 2021, 12, 663027.
- Fedele, G.; Brischetto, C.; Rossi, V. Biocontrol of Botrytis cinerea on Grape Berries as Influenced by Temperature and Humidity. Front. Plant Sci. 2020, 11, 1232.
- Bigot, G.; Mosetti, D.; Cargnus, E.; Freccero, A.; Moosavi, F.K.; Lujan, C.; Stecchina, M.; Tacoli, F.; Zandigiacomo, P.; Sivilotti, P.; et al. Influence of Vineyard Inter-Row Management and Clone on “Sauvignon Blanc” Performance in Friuli Venezia Giulia (North-Eastern Italy). Vitis-J. Grapevine Res. 2022, 61, 53–62.
- Habib, W.; Khalil, J.; Mincuzzi, A.; Saab, C.; Gerges, E.; Tsouvalakis, H.; Ippolito, A.; Sanzani, S. Fungal Pathogens Associated with Harvested Table Grapes in Lebanon, and Characterization of the Mycotoxigenic Genera. Phytopathol. Mediterr. 2021, 60, 427–439.
- Hahn, M. The Rising Threat of Fungicide Resistance in Plant Pathogenic Fungi: Botrytis as a Case Study. J. Chem. Biol. 2014, 7, 133–141.
- Fan, F.; Zhu, Y.; Wu, M.; Yin, W.-X.; Li, G.-Q.; Hanh, M.; Hamada, M.S.; Luo, C.-X. Mitochondrial Inner Membrane ABC Transporter Bcmdl1 Is Involved in Conidial Germination, Virulence, and Resistance to Anilinopyrimidine Fungicides in Botrytis cinerea. Microbiol. Spectr. 2023, 11, e0010823.
- Kretschmer, M.; Leroch, M.; Mosbach, A.; Walker, A.-S.; Fillinger, S. Fungicide-Driven Evolution and Molecular Basis of Multidrug Resistance in Field Populations of the Grey Mould Fungus Botrytis cinerea. PLoS Pathog. 2009, 5, 1000696.
- Saito, S.; Michailides, T.J.; Xiao, C.L. Fungicide-Resistant Phenotypes in Botrytis cinerea Populations and Their Impact on Control of Gray Mold on Stored Table Grapes in California. Eur. J. Plant Pathol. 2019, 154, 203–213.
- Testempasis, S.; Puckett, R.D.; Michailides, T.J.; Karaoglanidis, G.S. Genetic Structure and Fungicide Resistance Profile of Botrytis spp. Populations Causing Postharvest Gray Mold of Pomegranate Fruit in Greece and California. Postharvest Biol. Technol. 2020, 170, 111319.
- Mincuzzi, A.; Sanzani, S.M.; Palou, L.; Ragni, M.; Ippolito, A. Postharvest Rot of Pomegranate Fruit in Southern Italy: Characterization of the Main Pathogens. J. Fungi 2022, 8, 475.
- Shen, Y.; Yang, H. Effect of Preharvest Chitosan-g-Salicylic Acid Treatment on Postharvest Table Grape Quality, Shelf Life, and Resistance to Botrytis cinerea-Induced Spoilage. Sci. Hortic. 2017, 224, 367–373.
- Shahrajabian, M.H.; Chaski, C.; Polyzos, N.; Tzortzakis, N.; Petropoulos, S.A. Sustainable Agriculture Systems in Vegetable Production Using Chitin and Chitosan as Plant Biostimulants. Biomolecules 2021, 11, 819.
- EFSA National Summary Reports on Pesticide Residue Analysis Performed in 2018. EFSA Support. Publ. 2020, 17, 1814E.
- Zhang, Z.; Zhao, P.; Zhang, P.; Su, L.; Jia, H.; Wei, X.; Fang, J.; Jia, H. Integrative Transcriptomics and Metabolomics Data Exploring the Effect of Chitosan on Postharvest Grape Resistance to Botrytis cinerea. Postharvest Biol. Technol. 2020, 167, 111248.
- Youssef, K.; de Oliveira, A.G.; Tischer, C.A.; Hussain, I.; Roberto, S.R. Synergistic Effect of a Novel Chitosan/Silica Nanocomposites-Based Formulation against Gray Mold of Table Grapes and Its Possible Mode of Action. Int. J. Biol. Macromol. 2019, 141, 247–258.
- Youssef, K.; Roberto, S.R. Chitosan/Silica Nanocomposite-Based Formulation Alleviated Gray Mold through Stimulation of the Antioxidant System in Table Grapes. Int. J. Biol. Macromol. 2021, 168, 242–250.
- De Simone, N.; Pace, B.; Grieco, F.; Chimienti, M.; Tyibilika, V.; Santoro, V.; Capozzi, V.; Colelli, G.; Spano, G.; Russo, P.; et al. Botrytis cinerea and Table Grapes: A Review of the Main Physical, Chemical, and Bio-Based Control Treatments in Post-Harvest. Foods 2020, 9, 1138.
- Reglinski, T.; Elmer, P.A.G.; Taylor, J.T.; Wood, P.N.; Hoyte, S.M. Inhibition of Botrytis cinerea Growth and Suppression of Botrytis Bunch Rot in Grapes Using Chitosan. Plant Pathol. 2010, 59, 882–890.
- Rajestary, R.; Xylia, P.; Chrysargyris, A.; Romanazzi, G.; Tzortzakis, N. Preharvest Application of Commercial Products Based on Chitosan, Phosphoric Acid Plus Micronutrients, and Orange Essential Oil on Postharvest Quality and Gray Mold Infections of Strawberry. Int. J. Mol. Sci. 2022, 23, 15472.
- Palou, L.; Usall, J.; Munoz, A.; Smilanick, J.; Vinas, I. Hot Water, Sodium Carbonate, and Sodium Bicarbonate for the Control of Postharvest Green and Blue Molds of clementine Mandarins. Postharvest Biol. Technol. 2002, 24, 93–96.
- Nigro, F.; Schena, L.; Ligorio, A.; Pentimone, I.; Ippolito, A.; Salerno, M.G. Control of Table Grape Storage Rots by Pre-Harvest Applications of Salts. Postharvest Biol. Technol. 2006, 42, 142–149.
- Ding, S.; Meinholz, K.; Cleveland, K.; Jordan, S.A.; Gevens, A.J. Diversity and Virulence of Alternaria spp. Causing Potato Early Blight and Brown Spot in Wisconsin. Phytopathology 2019, 109, 436–445.
- Kumar Yadav, V.; Kumar, V.; Mani, A. Evaluation of Fungicides, Biocontrol Agents and Plant Extracts against Early Blight of Potato Caused by Alternaria solani. Int. J. Chem. Stud. 2018, 6, 1227–1230.
- He, D.C.; He, M.H.; Amalin, D.M.; Liu, W.; Alvindia, D.G.; Zhan, J. Biological Control of Plant Diseases: An Evolutionary and Eco-Economic Consideration. Pathogens 2021, 10, 1311.
- Palmieri, D.; Ianiri, G.; Del Grosso, C.; Barone, G.; De Curtis, F.; Castoria, R.; Lima, G. Advances and Perspectives in the Use of Biocontrol Agents against Fungal Plant Diseases. Horticulturae 2022, 8, 577.
- Kowalska, J.; Tyburski, J.; Matysiak, K.; Tylkowski, B.; Malusá, E. Field Exploitation of Multiple Functions of Beneficial Microorganisms for Plant Nutrition and Protection: Real Possibility or Just a Hope? Front. Microbiol. 2020, 11, 1904.
- Dong, S.M.; Zhou, S.Q. Potato Late Blight Caused by Phytophthora infestans: From Molecular Interactions to Integrated Management Strategies. J. Integr. Agric 2022, 21, 3456–3466.
- Huang, X.; You, Z.; Luo, Y.; Yang, C.; Ren, J.; Liu, Y.; Wei, G.; Dong, P.; Ren, M. Antifungal Activity of Chitosan against Phytophthora infestans, the Pathogen of Potato Late Blight. Int. J. Biol. Macromol. 2021, 166, 1365–1376.
- Amborabé, B.E.; Bonmort, J.; Fleurat-Lessard, P.; Roblin, G. Early Events Induced by Chitosan on Plant Cells. J. Exp. Bot. 2008, 59, 2317–2324.
- El Hadrami, A.; Adam, L.R.; El Hadrami, I.; Daayf, F. Chitosan in Plant Protection. Mar. Drugs 2010, 8, 968–987.
- Rabea, E.I.; Badawy, M.E.T.; Stevens, C.V.; Smagghe, G.; Steurbaut, W. Chitosan as Antimicrobial Agent: Applications and Mode of Action. Biomacromolecules 2003, 4, 1457–1465.
- Acar, O.; Aki, C.; Erdugan, H. Fungal and Bacterial Diseases Control with Elexa Plant Booster. Fresenius Environ. Bull. 2008, 17, 797–802.
- Nechwatal, J.; Zellner, M. Potential Suitability of Various Leaf Treatment Products as Copper Substitutes for the Control of Late Blight (Phytophthora infestans) in Organic Potato Farming. Potato Res. 2015, 58, 261–276.
- Hadwiger, L.A.; McBride, P.O. Low-Level Copper Plus Chitosan Applications Provide Protection Against Late Blight of Potato. Plant Health Prog. 2006, 6, 7.
- Žabka, M.; Pavela, R. The Dominance of Chitosan Hydrochloride over Modern Natural Agents or Basic Substances in Efficacy against Phytophthora infestans, and Its Safety for the Non-Target Model Species Eisenia fetida. Horticulturae 2021, 7, 366.
- Wang, X.; Zheng, K.; Cheng, W.; Li, J.; Liang, X.; Shen, J.; Dou, D.; Yin, M.; Yan, S. Field Application of Star Polymer-Delivered Chitosan to Amplify Plant Defense against Potato Late Blight. Chem. Eng. J. 2021, 417, 129327.
- Wasternack, C.; Feussner, I. The Oxylipin Pathways: Biochemistry and Function. Annu. Rev. Plant Biol. 2018, 69, 363–386.
- Trebbi, G.; Negri, L.; Bosi, S.; Dinelli, G.; Cozzo, R.; Marotti, I. Evaluation of Equisetum Arvense (Horsetail Macerate) as a Copper Substitute for Pathogen Management in Field-Grown Organic Tomato and Durum Wheat Cultivations. Agriculture 2021, 11, 5.
- Garmendia, A.; Raigoń, M.D.; Marques, O.; Ferriol, M.; Royo, J.; Merle, H. Effects of Nettle Slurry (Urtica dioica L.) Used as Foliar Fertilizer on Potato (Solanum tuberosum L.) Yield and Plant Growth. PeerJ 2018, 6, 4729.
- Behiry, S.I.; Philip, B.; Salem, M.Z.M.; Amer, M.A.; El-Samra, I.A.; Abdelkhalek, A.; Heflish, A. Urtica dioica and Dodonaea Viscosa Leaf Extracts as Eco-Friendly Bioagents against Alternaria Alternata Isolate TAA-05 from Tomato Plant. Sci. Rep. 2022, 12, 16468.
- Catuna (Petrar), T.; Odagiu, A.; Balint, C.; Darjan, S.; Bordea, D.; Mihaiescu, R. Testing the Anti-Alternariosis Effect of Aqueous Extract of Allium cepa L. in Potato. ProEnvironment 2021, 14, 87–90.
- Wianowska, D.; Olszowy-Tomczyk, M.; Garbaczewska, S. A Central Composite Design in Increasing the Quercetin Content in the Aqueous Onion Waste Isolates with Antifungal and Antioxidant Properties. Eur. Food Res. Technol. 2022, 248, 497–505.
- Narayanasamy, P. Ecology of Postharvest Microbial Pathogens. In Postharvest Pathogens and Disease Management; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2005; pp. 79–116.
- Aharoni, Y.; Barkai-Golan, R. Pre-Harvest Fungicide Sprays and Polyvinyl Wraps to Control Botrytis Rot and Prolong the Post-Harvest Storage Life of Strawberries. J. Hortic. Sci. 1987, 62, 177–181.
- Romanazzi, G.; Gabler, F.M.; Smilanick, J.L. Preharvest Chitosan and Postharvest UV Irradiation Treatments Suppress Gray Mold of Table Grapes. Plant Dis. 2006, 90, 445–450.
- Meng, X.; Li, B.; Liu, J.; Tian, S. Physiological Responses and Quality Attributes of Table Grape Fruit to Chitosan Preharvest Spray and Postharvest Coating during Storage. Food Chem. 2008, 106, 501–508.
- Ehtesham Nia, A.; Taghipour, S.; Siahmansour, S. Pre-Harvest Application of Chitosan and Postharvest Aloe Vera Gel Coating Enhances Quality of Table Grape (Vitis vinifera L. Cv. ‘Yaghouti’) during Postharvest Period. Food Chem. 2021, 347, 129012.
- Bhaskara Reddy, M.V.; Belkacemi, K.; Corcuff, R.; Castaigne, F.; Arul, J. Effect of Pre-Harvest Chitosan Sprays on Post-Harvest Infection by Botrytis cinerea Quality of Strawberry Fruit. Postharvest Biol. Technol. 2000, 20, 39–51.
- Mazaro, S.M.; Deschamps, C.; May de Mio, L.L.; Biasi, L.A.; de Gouvea, A.; Sautter, C.K. Comportamento Pós-Colheita de Frutos de Morangueiro Após a Aplicação Pré-Colheita de Quitosana e Acibenzolar-S-Metil. Rev. Bras. Frutic. 2008, 30, 185–190.
- He, Y.; Bose, S.K.; Wang, W.; Jia, X.; Lu, H.; Yin, H. Pre-Harvest Treatment of Chitosan Oligosaccharides Improved Strawberry Fruit Quality. Int. J. Mol. Sci. 2018, 19, 2194.
- Mazur, S.; Waksmundzka, A. Effect of Some Compounds on the Decay of Strawberry Fruits Caused by Botrytis cinerea Pers. Meded. Rijksuniv. Gent Fak. Landbouwkd. Toegep. Biol. Wet. 2001, 66, 227–231.
- Romanazzi, G.; Nigro, F.; Ippolito, A. Short Hypobaric Treatments Potentiate the Effect of Chitosan in Reducing Storage Decay of Sweet Cherries. Postharvest Biol. Technol. 2003, 29, 73–80.
- Ahmed, Z.F.R.; Kaur, N.; Maqsood, S.; Schmeda-Hirschmann, G. Preharvest Applications of Chitosan, Salicylic Acid, and Calcium Chloride Have a Synergistic Effect on Quality and Storability of Date Palm Fruit (Phoenix dactylifera L.). HortScience 2022, 57, 422–430.
- Zhang, C.; Long, Y.H.; Wang, Q.P.; Li, J.H.; An, H.M.; Wu, X.M.; Li, M. The Effect of Preharvest 28.6% Chitosan Composite Film Sprays for Controlling the Soft Rot on Kiwifruit and Its Defence Responses. Hortic. Sci. 2019, 46, 180–194.
- Cui, K.; Shu, C.; Zhao, H.; Fan, X.; Cao, J.; Jiang, W. Preharvest Chitosan Oligochitosan and Salicylic Acid Treatments Enhance Phenol Metabolism and Maintain the Postharvest Quality of Apricots (Prunus armeniaca L.). Sci. Hortic. 2020, 267, 109334.
- Elmenofy, H.M.; Okba, S.K.; Salama, A.M.; Alam-Eldein, S.M. Yield, Fruit Quality, and Storability of ‘Canino’ Apricot in Response to Aminoethoxyvinylglycine, Salicylic Acid, and Chitosan. Plants 2021, 10, 1838.
- El-Badawy, H.E.M. Effect of Chitosan and Calcium Chloride Spraying on Fruits Quality of Florida Prince Peach under Cold Storage. Res. J. Agric. Biol. Sci. 2012, 8, 272–281.
- Gayed, A.A.N.A.; Shaarawi, S.A.M.A.; Elkhishen, M.A.; Elsherbini, N.R.M. Pre-Harvest Application of Calcium Chloride and Chitosan on Fruit Quality and Storability of ‘Early Swelling’ Peach during Cold Storage. Ciênc. Agrotecnol. 2017, 41, 220–231.
- Yan, J.; Cao, J.; Jiang, W.; Zhao, Y. Effects of Preharvest Oligochitosan Sprays on Postharvest Fungal Diseases, Storage Quality, and Defense Responses in Jujube (Zizyphus Jujuba Mill. Cv. Dongzao) Fruit. Sci. Hortic. 2012, 142, 196–204.
- Migliori, C.A.; Salvati, L.; Di Cesare, L.F.; Lo Scalzo, R.; Parisi, M. Effects of Preharvest Applications of Natural Antimicrobial Products on Tomato Fruit Decay and Quality during Long-Term Storage. Sci. Hortic. 2017, 222, 193–202.
- Tezotto-Uliana, J.V.; Fargoni, G.P.; Geerdink, G.M.; Kluge, R.A. Chitosan Applications Pre- or Postharvest Prolong Raspberry Shelf-Life Quality. Postharvest Biol. Technol. 2014, 91, 72–77.
- Ippolito, A.; Schena, L.; Pentimone, I.; Nigro, F. Control of Postharvest Rots of Sweet Cherries by Pre- and Postharvest Applications of Aureobasidium pullulans in Combination with Calcium Chloride or Sodium Bicarbonate. Postharvest Biol. Technol. 2005, 36, 245–252.
- Mahmoud, G.; Ahmed, S.; Abbas, M.; Soliman, A.S. Effect of Garlic and Onion Extracts as a Preharvest Applications on the Post-Harvest Quality and Oxidative Enzyme Activity of Pearfruit during Cold Storage. J. Biol. Chem. Environ. Sci. 2018, 13, 329–356.
- Ahn, S.E.; Lee, A.Y.; Wang, M.H.; Hwang, Y.S. Increase of Strawberry Fruit Shelf-Life through Preharvest Spray of Calcium-Chitosan and Post-Harvest Treatment with High Pressure CO2. Hortic. Sci. Technol. 2014, 32, 636–644.
- Saavedra, G.M.; Figueroa, N.E.; Poblete, L.A.; Cherian, S.; Figueroa, C.R. Effects of Preharvest Applications of Methyl Jasmonate and Chitosan on Postharvest Decay, Quality and Chemical Attributes of Fragaria Chiloensis Fruit. Food Chem. 2016, 190, 448–453.
- EFSA (European Food Safety Authority) Outcome of the Consultation with Member States and EFSA on the Basic Substance Application for Calcium Hydroxide and the Conclusions Drawn by EFSA on the Specific Points Raised; EN-488; 2013; 41p. Available online: www.efsa.europa.eu/publications (accessed on 4 August 2023).
- Gaspari, M.; Lykouressis, D.; Perdikis, D.; Polissiou, M. Nettle Extract Effects on the Aphid Myzus Persicae and Its Natural Enemy, the Predator Macrolophus Pygmaeus (Hem., Miridae). J. Appl. Entomol. 2007, 131, 652–657.
- Thacker, J. An Introduction to Arthropod Pest Control; Cambridge University Press: Cambridge, UK, 2002; ISBN 9780521567879.
- González-Macedo, M.; Cabirol, N.; Rojas-Oropeza, M. Assessment of the Ancestral Use of Garlic (Allium Sativum) and Nettle (Urtica dioica) as Botanical Insecticides in the Protection of Mesquite (Prosopis laevigata) Seeds against Bruchins. J. Plant Prot. Res. 2021, 61, 170–175.
- Bozsik, A. Studies on Aphicidal Efficiency of Different Stinging Nettle Extracts. Anz. Schädl. Pflanzenschutz Umweltschutz 1996, 69, 21–22.
- Bozsik, A. Effect of Fermented Nettle Extract on Callaphis Juglandis. Godolloi Agrartud. Egyet. 1992, 28, 71–73.
- Dąbrowski, Z.T.; Seredyńska, U. Characterisation of the Two-Spotted Spider Mite (Tetranychus Urticae KOCH, Acari: Tetranychidae) Response to Aqueous Extracts from Selected Plant Species. J. Plant Prot. Res. 2007, 47, 113–124.
- Kapsoot, E.; Mwangi, M.; Kamau, A. Repellence and Toxicity Effect of Crude Plant Extracts on the Two-Spotted Spider Mite Tetranychus Urticae on Roses. Acta Hortic. 2015, 1077, 155–164.
- Pavela, R. History, Presence and Perspective of Using Plant Extracts as Commercial Botanical Insecticides and Farm Products for Protection against Insects—A Review. Plant Prot. Sci. 2016, 52, 229–241.
- Jairoce, C.F.; Teixeira, C.M.; Nunes, C.F.P.; Nunes, A.M.; Pereira, C.M.P.; Garcia, F.R.M. Insecticide Activity of Clove Essential Oil on Bean Weevil and Maize Weevil. Rev. Bras. Eng. Agric. Ambient. 2016, 20, 72–77.
- Isman, M.B. Commercial Development of Plant Essential Oils and Their Constituents as Active Ingredients in Bioinsecticides. Phytochem. Rev. 2020, 19, 235–241.
- Karkanis, A.C.; Athanassiou, C.G. Natural Insecticides from Native Plants of the Mediterranean Basin and Their Activity for the Control of Major Insect Pests in Vegetable Crops: Shifting from the Past to the Future. J. Pest Sci. 2021, 94, 187–202.
- Mkenda, P.; Mwanauta, R.; Stevenson, P.C.; Ndakidemi, P.; Mtei, K.; Belmain, S.R. Extracts from Field Margin Weeds Provide Economically Viable and Environmentally Benign Pest Control Compared to Synthetic Pesticides. PLoS ONE 2015, 10, e0143530.
- Bolouri Moghaddam, M.; Van den Ende, W. Sugars and Plant Innate Immunity. J. Exp. Bot. 2012, 63, 3989–3998.
- Tarkowski, Ł.P.; Van de Poel, B.; Höfte, M.; Van den Ende, W. Sweet Immunity: Inulin Boosts Resistance of Lettuce (Lactuca sativa) against Grey Mold (Botrytis cinerea) in an Ethylene-Dependent Manner. Int. J. Mol. Sci. 2019, 20, 1052.
- Trouvelot, S.; Héloir, M.C.; Poinssot, B.; Gauthier, A.; Paris, F.; Guillier, C.; Combier, M.; Trdá, L.; Daire, X.; Adrian, M. Carbohydrates in Plant Immunity and Plant Protection: Roles and Potential Application as Foliar Sprays. Front. Plant Sci. 2014, 5, 592.
- Derridj, S.; Cabanat, I.; Cochet, E.; Couzi, P.; Lombarkia, N.; Wu, B. Incidence Des Métabolites Présents à La Surface Des Organes Du Pommier Sur Le Comportement de Cydia Pomonella (Lepidoptera, Tortricidae). In Proceedings of the ANPP—5ème Conférence Internationale sur les Ravageurs en Agriculture, Montpellier, France, 7–9 December 1999; pp. 279–286.
- Lombarkia, N.; Derridj, S. Incidence of Apple Fruit and Leaf Surface Metabolites on Cydia pomonella Oviposition. Entomol. Exp. Appl. 2002, 104, 79–87.
- Derridj, S.; Moulin, F.; Ferré, E.; Galy, H.; Bergougnoux, A.; Arnaud, I.; Auger, J. Sucrose as an Apple Tree Resistance Inducer against Cydia Pomonella L. In Proceedings of the 7th International Conference on Integrated Fruit Production, Avignon, France, 27–30 October 2008; Cross, J., Brown, M., Fitzgerald, J., Fountain, M., Yohalem, D., Eds.; IOBC Working Groups “Integrated Fruit Protection in Fruit Crops″: Avignon, France, 2008; p. 66.
- Lombarkia, N.; Derridj, S. Resistance of Apple Trees to Cydia Pomonella Egg-Laying Due to Leaf Surface Metabolites. Entomol Exp. Appl. 2008, 128, 57–65.
- Arnault, I.; Lombarkia, N.; Joy-Ondet, S.; Romet, L.; Brahim, I.; Meradi, R.; Nasri, A.; Auger, J.; Derridj, S. Foliar Application of Microdoses of Sucrose to Reduce Codling Moth Cydia pomonella L. (Lepidoptera: Tortricidae) Damage to Apple Trees. Pest Manag. Sci. 2016, 72, 1901–1909.
- Tiffrent, A.E.K.; Lombarkia, N. Assessement of Control Strategy by Spraying Low Doses of Sugars on Apple Orchard against Cydia pomonella (Linnaeus, 1758.). Acta Agric. Slov. 2021, 117, 1–6.
- Tiffrent, A.-K.; Lombarkia, N. Effect of Foliar Application of Glucose and Fructose to Reduce Codling Moth (Cydia pomonella ) Damages on Apple Orchard. Acta Agric. Slov. 2022, 118, 1–6.
- Tiffrent, A.; Lombarkia, N. Effect of the Exogenous Foliar Sprays of Micro-Doses of Fructose and Glucose, on Egg-Laying of Cydia pomonella L. and its Oviposition Site Selection in Apple Orchard. J. Bioresour. Manag. 2022, 9, 85–91.
- Derridj, S.; Fiala, V. Soluble Sugars of Maize Leaves (Zea mays L.) and Oviposition of the European Corn Borer (Ostrinia Nubilalis Hbn.). C. R. Seances L’acad. D’agric. Fr. 1983, 69, 465–472.
- Fiala, V.; Derridj, S.; Jolivet, E. Influence de La Teneur En Glucides Solubles Des Feuilles de Zea mays L. Sur Le Choix Du Site de Ponte de La Pyrale, Ostrinia Nubilalis Hbn. (Lepid. Pyralidae). Agronomie 1985, 5, 927–932.
- Derridj, S.; Gregoire, V.; Boutin, J.P.; Fiala, V. Plant Growth Stages in the Interspecific Oviposition Preference of the European Corn Borer and Relations with Chemicals Present on the Leaf Surfaces. Entomol. Exp. Appl. 1989, 53, 267–276.
- Derridj, S.; Fiala, V.; Barry, P.; Robert, P.; Roessingh, P.; Städler, E. Role of Nutrients Found in the Phylloplane, in the Insect Host-Plant Selection for Oviposition. In Proceedings of the 8th International Symposium on Insect-Plant Relationships; Springer: Dordrecht, The Netherlands, 1992; Volume 49, pp. 139–140.
- Suverkropp, B.P.; Dutton, A.; Bigler, F.; Van Lenteren, J.C. Oviposition Behaviour and Egg Distribution of the European Corn Borer, Ostrinia Nubilalis, on Maize, and Its Effect on Host Finding by Trichogramma Egg Parasitoids. Bull. Insectol. 2008, 61, 303–312.
- Arnault, I.; Zimmermann, M.; Furet, A.; Chovelon, M.; Thibord, J.; Derridj, S. Fructose and Sucrose as Priming Molecules against Pathogens and Pests? In Proceedings of the Ecological Perspectives of Induced Resistance in Plants and Multitrophic Interactions in Soil, Riva del Garda (TN), Italy, 18–20 October 2017; Perazzolli, M., Puopolo, G., Pertot, I., Pieterse, C., Mauch-Mani, B., Schmitt, A., Flors, V., Eds.; IOBC-WPRS Bulletin: Riva del Garda, Italy, 2018; Volume 135, pp. 110–112.
- Derridj, S.; Lombarkia, N.; Garrec, J.P.; Galy, H.; Ferré, E. Sugars on Leaf Surfaces Used as Signals by the Insect and the Plant: Implications in Orchard Protection against Cydia pomonella L. (Lepidoptera, Tortricidae). In Moths: Types, Ecological Significance and Control; Nova Science Publishers Inc.: Hauppage, NY, USA, 2012; pp. 1–38. ISBN 9781614706267.
- Mijailovic, N.; Nesler, A.; Perazzolli, M.; Aït Barka, E.; Aziz, A. Rare Sugars: Recent Advances and Their Potential Role in Sustainable Crop Protection. Molecules 2021, 26, 1720.
- Station d’experimentation La Pugere. The TALC Efficiency Evaluation in a Preventive Control Strategy of the Pear Psylla Year; Station d’experimentation La Pugere: Mallemort, France, 2011; pp. 1–68.
- Marchand, P. Basic Substances under EC 1107/2009 Phytochemical Regulation: Experience with Non-Biocide and Food Products as Biorationals. J. Plant Prot. Res. 2016, 56, 312–318.
- Warlop, F. Évaluation de l’efficacité de Produits Naturels Vis-à-Vis de La Mouche de L’olivier. Available online: http://www.grab.fr/wp-content/uploads/2014/07/CR_mouche_olive_20131.pdf (accessed on 26 July 2023).
- De Smedt, C.; Someus, E.; Spanoghe, P. Potential and Actual Uses of Zeolites in Crop Protection. Pest Manag. Sci. 2015, 71, 1355–1367.
- Eroglu, N. A Review: Insecticidal Potential of Zeolite (Clinoptilolite), Toxicity Ratings and General Properties of Turkish Zeolites. In Proceedings of the 11th International Working Conference on Stored Product Protection, Chiang Mai, Thailand, 24–28 November 2014; pp. 755–767.
- Barrer, R.M. Zeolites and Clay Minerals as Sorbents and Molecular Sieves; Academic Press: London, UK, 1978; ISBN 0120793504.
- Christidis, G.E.; Moraetis, D.; Keheyan, E.; Akhalbedashvili, L.; Kekelidze, N.; Gevorkyan, R.; Yeritsyan, H.; Sargsyan, H. Chemical and Thermal Modification of Natural HEU-Type Zeolitic Materials from Armenia, Georgia and Greece. Appl. Clay Sci. 2003, 24, 79–91.
- Joint FAO/WHO Food Standards Programme. Codex Alimentarius Commission; Food and Agriculture Organization of the United Nations: Rome, Italy, 2019.
- Haryadi, Y.; Syarief, R.; Hubeis, M.; Herawati, I. Effect of Zeolite on the Development of Sitophilus Zeamais Motsch. In Proceedings of the 6th International Working Conference on Stored-Product Protection, Canberra, Australia, 17–23 April 1994; Highley, E., Wright, E.J., Banks, H.J., Champ, B.R., Eds.; CAB International: Wallingford, UK, 2010; pp. 633–634.
- Kljajic, P. Protection of Stored Plant Products from Harmful Organisms; Institut za Pesticide i Zaštitu Životne Sredine: Beograd, Serbia, 2008; ISBN 9788686869029.
- Kljajić, P.; Andrić, G.; Adamović, M.; Bodroža-Solarov, M.; Marković, M.; Perić, I. Laboratory Assessment of Insecticidal Effectiveness of Natural Zeolite and Diatomaceous Earth Formulations against Three Stored-Product Beetle Pests. J. Stored Prod. Res. 2010, 46, 1–6.
- Kljajić, P.; Andrić, G.; Adamović, M.; Golić, M.P. Laboratory Evaluation of Insecticidal Effectiveness of a Natural Zeolite Formulation against Sitophilus oryzae (L.), Rhyzopertha dominica (F.) and Tribolium castaneum (Herbst) in Treated Wheat. In Proceedings of the 10th International Working Conference on Stored Product Protection, 27 June–2 July 2010, Estoril, Portugal; Julius-Kuhn-Archiv: Quedlinburg, Germany; pp. 863–868.
- Kljajić, P.J.; Andrić, G.G.; Adamović, M.; Golić, M.P. Possibilities of Application of Natural Zeolites in Stored Wheat Grain Protection against Pest Insects. J. Process. Energy Agric. 2011, 15, 12–15.
- Andrić, G.G.; Marković, M.M.; Adamović, M.; Daković, A.; Golić, M.P.; Kljajić, P.J. Insecticidal Potential of Natural Zeolite and Diatomaceous Earth Formulations against Rice Weevil (Coleoptera: Curculionidae) and Red Flour Beetle (Coleoptera: Tenebrionidae). J. Econ. Entomol. 2012, 105, 670–678.
- Daniel, C.; Dierauer, H.; Clerc, M. The Potential of Silicate Rock Dust to Control Pollen Beetles (Meligethes spp.). IOBC wprs Bull. 2013, 96, 47–55.
- Rumbos, C.I.; Sakka, M.; Berillis, P.; Athanassiou, C.G. Insecticidal Potential of Zeolite Formulations against Three Stored-Grain Insects, Particle Size Effect, Adherence to Kernels and Influence on Test Weight of Grains. J. Stored Prod. Res. 2016, 68, 93–101.
- Lü, J.; Sehgal, B.; Subramanyam, B. Insecticidal Potential of a Synthetic Zeolite against the Cowpea Weevil, Callosobruchus Maculatus (Fabricius) (Coleoptera: Bruchidae). J. Stored Prod. Res. 2017, 72, 28–34.
- Eroglu, N.; Sakka, M.K.; Emekci, M.; Athanassiou, C.G. Effects of Zeolite Formulations on the Mortality and Progeny Production of Sitophilus Oryzae and Oryzaephilus Surinamensis at Different Temperature and Relative Humidity Levels. J. Stored Prod. Res. 2019, 81, 40–45.
- Checchia, I.; Perin, C.; Mori, N.; Mazzon, L. Oviposition Deterrent Activity of Fungicides and Low-Risk Substances for the Integrated Management of the Olive Fruit Fly Bactrocera Oleae (Diptera, Tephritidae). Insects 2022, 13, 363.
- Dickens, J.C. Olfaction in the Boll Weevil, Anthonomus Grandis Boh. (Coleoptera: Curculionidae): Electroantennogram Studies. J. Chem. Ecol. 1984, 10, 1759–1785.
- Visser, J. Host Odor Perception in Phytophagous Insects. Annu. Rev. Entomol. 1986, 31, 121–144.
- Katsoyannos, B.I.; Papadopoulos, N.T.; Stavridis, D. Evaluation of Trap Types and Food Attractants for Rhagoletis Cerasi (Diptera: Tephritidae). J. Econ. Entomol. 2000, 93, 1005–1010.
- Mazor, M.; Gothfil, S.; Galun, R. The Role of Ammonia in the Attraction of Females of the Mediterranean Fruit Fly to Protein Hydrolysate Baits. Entomol. Exp. Appl. 1987, 43, 25–29.
- Epsky, N.; Heath, R. Exploiting the Interactions of Chemical and Visual Cues in Behavioral Control Measures for Pest Tephritid Fruit Flies. Fla. Entomol. 2010, 81, 273–282.
- Hull, C.D.; Cribb, B.W. Olfaction in the Queensland Fruit Fly, Bactrocera Tryoni. I: Identification of Olfactory Receptor Neuron Types Responding to Environmental Odors. J. Chem. Ecol. 2001, 27, 871–887.
- Sarles, L.; Verhaeghe, A.; Francis, F.; Verheggen, F.J. Semiochemicals of Rhagoletis Fruit Flies: Potential for Integrated Pest Management. Crop Prot. 2015, 78, 114–118.
- Shelly, T.; Nishimoto, J.; Kurashima, R. Trap Capture of Three Economically Important Fruit Fly Species (Diptera: Tephritidae): Evaluation of a Solid Formulation Containing Multiple Male Lures in a Hawaiian Coffee Field. J. Econ. Entomol. 2012, 105, 1186–1193.
- Montiel Bueno, A.; Jones, O. Alternative Methods for Controlling the Olive Fly, Bactrocera Oleae, Involving Semiochemicals. Bull. OILB/SROP 2002, 25, 147–155.
- EFSA. Outcome of the Consultation with Member States and EFSA on the Basic Substance Application for Onion Oil for Use in Plant Protection as Repellent. EFSA Support. Publ. 2017, 14, 1315E.
- Sharif, R.; Mujtaba, M.; Rahman, M.U.; Shalmani, A.; Ahmad, H.; Anwar, T.; Tianchan, D.; Wang, X. The Multifunctional Role of Chitosan in Horticultural Crops; a Review. Molecules 2018, 23, 872.
- Rabea, E.I.; Badawy, M.E.I.; Rogge, T.M.; Stevens, C.V.; Höfte, M.; Steurbaut, W.; Smagghe, G. Insecticidal and Fungicidal Activity of New Synthesized Chitosan Derivatives. Pest Manag. Sci. 2005, 61, 951–960.
- Casals, P.; Cardenas, G.; Galvez, G.; Villar, A.; Cabrera, G. Agricultural Applications of Chitosan and Derivatives. In Proceedings of the Proceedings 10th IUPAC International Congress on the Chemistry of Crop Protection, Basel, Switzerland, 4–9 August 2002; p. 228.
- Zhang, M.; Tan, T.; Yuan, H.; Rui, C. Insecticidal and Fungicidal Activities of Chitosan and Oligo-Chitosan. J. Bioact. Compat. Polym. 2003, 18, 391–400.
- Li, Y.; Qin, Y.; Liu, S.; Xing, R.; Yu, H.; Li, K.; Li, P. Preparation, Characterization, and Insecticidal Activity of Avermectin-Grafted-Carboxymethyl Chitosan. Biomed. Res. Int. 2016, 2016, 9805675.
- Abbey, J.; Percival, D.; Abbey, L.; Asiedu, S.; Prithiviraj, B.; Schilder, A. Biofungicides as Alternative to Synthetic Fungicide Control of Grey Mould (Botrytis cinerea)–Prospects and Challenges. Biocontrol Sci. Technol. 2019, 29, 241–262.
- Fravel, D.R.; Deahl, K.L.; Stommel, J.R. Compatibility of the Biocontrol Fungus Fusarium oxysporum Strain CS-20 with Selected Fungicides. Biol. Control 2005, 34, 165–169.
More