1. Introduction
The hallmarks of cancer are characteristic alterations in the cells that accompany or lead to the development and progression of cancer. One of these hallmarks is uncontrolled or dysregulated angiogenesis. This allows the tumor mass to be provided with nutrients as a result of the development of new blood vessels (reviewed in
[1]). Recently, evidence brought forward shows how the creation of new nerve tissue, neurogenesis, is another important contributing factor to the development and progression of cancer
[2][3][4][2,3,4]. Multiple studies indicate that tumors in tissue with a high level of blood vessels present are more likely to develop intra-tumoral neural infiltration, a condition that is associated with a poor prognosis. The more extensive this intra-tumoral nerve density is, the more severe the metastatic potential of the tumor and the poorer the prognosis is for the patient
[5][6][7][5,6,7]. In the body, the distribution of blood vessels and nerves mirrors each other (
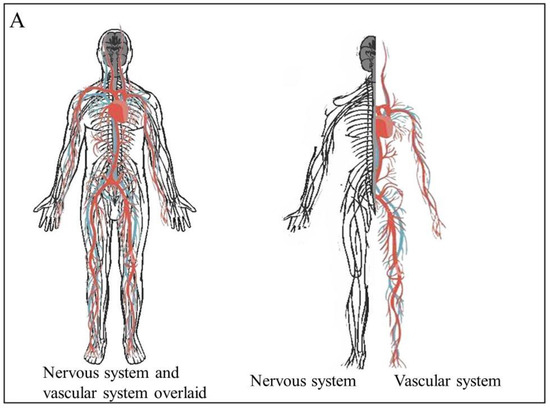
Figure 1A). This is due their embryological and growth factor similarity - they share many of the same molecules and signaling pathways that guide their growth
[8]. These shared molecules and pathways may be the reason why the molecules that induce angiogenesis during cancer also induce neurogenesis. Examples of the interaction between the nervous tissue and the surrounding blood vessels include the blood brain barrier (BBB), where astrocytes and pericytes around blood vessels interact with these blood vessels to aid in the regulation of the BBR. Interactions between the nervous tissue and the blood vessels surrounding it are known as neurovascular units (NVUs) (
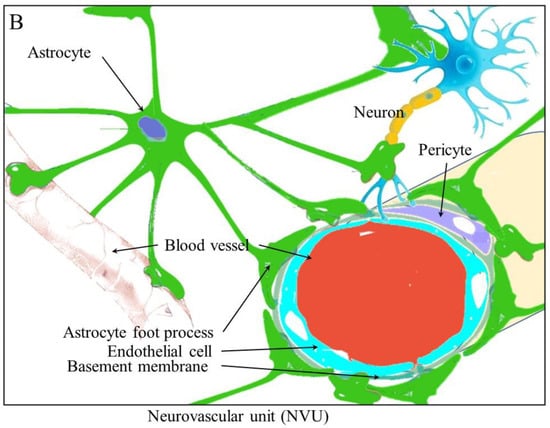
Figure 1B)
[9]. Brain tumors are known to alter the structure of the blood brain barrier (BBB) into what is known as the blood–tumor barrier (BTB). It is now known that cancers can drive neurogenesis, axonogenesis, and the repurposing of existing nerve fibers.
Figure 1. The physiological relationship between the vascular and nervous systems. (A) The vascular and neural systems mirror each other in the body as they both are required to service the entire body (B). The structure of the neurovascular unit in the brain and the blood brain barrier.
The new nervous tissue formed during neurogenesis is also dependent on new blood vessels to supply the nervous tissue with oxygen and nutrients to ensure the new tissues’ continued survival
[10].
2. Angiogenesis and Neurogenesis
The reciprocal communication between tumors and the nervous system is evidenced by the fact that cancer patients experience cancer related pain
[11][12] as a result of neuro-oncogenic pressure on the fibers as the tumor volume increases
[12][13], secretion of stimulatory factors on peripheral fibers with depolarizing effects, axon demyelination, and pathological neural plasticity induced by tumor-derived factors
[13][14].
The term angiogenesis describes the generation of new blood vessels. There are two main methods whereby angiogenesis occurs. Vasculogenic involves vascular progenitor cells forming new blood vessels. These circulating endothelial progenitor cells (EPCs) are derived from the bone marrow
[14][15]. These cells possess surface markers, such as CD34, CD31, VEGFR-2, and Tie-2. Tumor cells are known to secrete VEGF and cytokines such as stromal-derived factor-1. These pro-angiogenic factors recruit the circulating progenitor cells and stimulate proliferation. These pro-angiogenic stimuli also activate the matrix metalloproteases to break down the extracellular matrix, allowing cells to migrate and penetrate cell layers
[15][16].
Sprouting angiogenesis involves the formation of new vascular structures from an existing vessel. This, firstly, involves the destabilization of endothelial cells and endothelial mesenchymal transition. Once again, activated proteases are required to degrade the ECM and basement membrane, allowing for directed migration and proliferation
[15][16]. Once the cells form the lumen and tubulogenesis of the new blood vessels, the transition is reversed, with the cells reverting to the resting state
[16][17]. This process involves the VEGF and Notch signaling pathways
[17][18]. As such, sprouting angiogenesis is a process whereby new blood vessels are formed from pre-existing blood vessels. This process occurs in response to both mechanical and chemical stimuli and is important for normal development and wound healing. It facilitates tumor survival and progression as it provides cancer cells with oxygen and nutrients to sustain their growth (reviewed in
[18][19]). During angiogenesis, new micro vessels are formed as they branch off from pre-existing vessels
[19][20].
Angiogenesis involves processes, such as proliferation of endothelial cells and the formation of tube-like vascular structures, as well as branching and anastomosis (reviewed in
[20][21]). This re
svie
arch w will mainly focus on sprouting angiogenesis. Both pro- and anti-angiogenic factors act to regulate angiogenesis, with any changes in the balance of these factors either promoting or inhibiting angiogenesis. Pro-angiogenic factors are released by endothelial cells, monocytes, and tumor cells. Angiogenesis also requires the activation of various processes, such as basement membrane degradation, endothelial cell proliferation, migration, and cell remodeling, to form a tube or blood vessel (reviewed in
[21][22]). Angiogenesis can be promoted by adrenergic signaling, which is thought to contribute to the formation of preneoplastic niches and decreased immune function
[22][23]. β-adrenergic receptor activation has been found to be essential in the progression and malignant growth of ovarian
[23][24], pancreatic
[24][25], and pulmonary
[25][26] cancers.
In adult mice, neurogenesis predominantly occurs in the subventricular zone (SVZ) and the hippocampal dentate gyrus (DG), and it is thought that these two areas are where neural stem cells are found within the adult brain
[26][27][27,28]. Different types of cells are involved in neurogenesis. The slow-dividing or true stem cells are mostly quiescent but activate and divide asymmetrically to self-renew and give rise to immediate progenitor cells
[26][27]. These intermediate progenitor cells divide rapidly to give rise to nervous tissue. In addition to these progenitor cells, other types of cells integrate into existing neuronal networks. These include neuroblasts
[28][29][29,30] and newborn neurons, which are electro-physiologically active
[30][31]. Neurogenesis can be activated or inhibited by a multitude of signals, including growth factors, cytokines, chemokines, neurotrophins, steroids and extracellular matrix components, the activation of specific transcription factors, and signal transduction pathways (reviewed by
[31][32]). Additional signals may result from environmental stimuli, such as exercise alterations in an organism’s environment, stress, or social isolation
[32][33][33,34]. The growth and migration of tumors around and along nerve fibers are known as perineural invasion
[34][35]. Additionally, it is now known that nerves actively grow into and throughout cancer tissue, resulting in increased metastasis as these nerves provide signals to the cancer as well as a pathway to migrate along
[35][36]. Autonomic neurotransmitter receptors can stimulate cancer cell growth through the activation of corresponding signaling pathways
[36][37]. In its most basic form, the crosstalk between neurogenesis, angiogenesis, and carcinogenesis is based upon common signaling pathways and chemokines. Cancer cells express neurotrophic and pro-angiogenic markers. In many cases, these molecules are both neurotrophic and pro-angiogenic. The infiltration of new nervous tissue into cancer cells gives cancer cells a route to migrate along in the same way that these cells can use lymphatic and blood vessels. In other words, the signaling pathways activated by more cells stimulate both neurogenesis and angiogenesis while assisting in metastasis
[37][38].
3. The Remodeling of Tissues and the Proliferation and Migration of Cells during Angiogenesis and Neurogenesis
When malignant cells invade surrounding tissues, they displace the normal cells or integrate them, altering their function. These cells that are co-opted by cancer cells to contribute to the survival of the tumor include fibroblasts
[38][59], endothelial cells
[39][60], immune cells
[40][41][61,62], and neuronal extensions
[42][63]. Metastatic tumors then need to form capillaries to supply oxygen and nutrients but also to act as routes for further cell movement for metastatic dissemination
[43][64]. Nerve fibers can provide similar functions by providing the cancer cells with nerve cell signals as well as serving as migrations routes
[44][65]. Both nerve cells and the endothelial cells that become blood vessels are attracted to and co-opted by the cancer cells
[45][66]. Neurotrophins, such as nerve growth factor (NGF), can be released by leukocytes, such as macrophages and mast cells, to promote an axonogenic switch resulting in tumor innervation. Many immune cells express NGF
[46][67] following induction by IL-1β
[47][68]. This can occur during inflammatory pain and neurogenesis. In a mouse model of arthritis, the activation of macrophages results in increased levels of innervation, related to and in conjunction with angiogenesis
[48][69].
Neurogenesis and angiogenesis require increased proliferation of neural and endothelial cells. This can be achieved by altering the length of the cell cycle, shortening it, and allowing for more cycles of division
[49][70]. Alternatively, the rate of different types of divisions can be altered since symmetric divisions will increase the number of stem cells, while asymmetric division will result in a differentiated cell and one stem cell being generated
[50][71].
The beta-adrenergic receptors, β2/3 receptors, have been shown to be involved in tumor development and progression. In a mice model, the lack of the β2 or β3 receptor led to a delay in tumor growth and angiogenesis
[5][51][5,72]. In prostate cancer, these receptors lead to the stimulation of endothelial cells, resulting in angiogenesis by metabolic adjustments. The sympathetic nerves release noradrenaline, which activates β2-signaling in endothelial cells, leading to the expression of the mitochondrial cytochrome c oxidase component, COA6. This results in a decrease in normal oxidative respiration and the induction of angiogenesis
[51][72]. This noradrenaline also promotes vascular endothelial growth factor (VEGF) expression, leading to angiogenesis
[52][73].
3.1. Migration and Remodeling
Both angiogenesis and neurogenesis require the initiation or alteration of cell migration as well as the remodeling of tissue, which, themselves, require the breakdown of the extracellular membrane. In order to accomplish this altered migration and tissue remodeling, there must be specific interactions between cells, such as the immature migrating neuroblasts, astrocytic processes, and blood vessels
[53][74]. Some of the molecules involved in these processes include stromal-cell-derived factor-1 (SDF-1), CXC chemokine receptor 4 (CXCR4), monocyte chemoattractant protein-1 (MCP-1), and matrix metalloproteinases (MMPs).
3.2. Stromal-Cell-Derived FactorF-1 and CXC Chemokine Receptor R4
The alpha chemokine SDF-1 protein is normally excreted from the ependymal cells; it is also known as CXCL12 and, under normal conditions, induces neural stem cell (NSC) quiescence
[54][75]. However, under abnormal conditions such as in ischemic shock, SDF-1 is released from reactive astrocytes and vascular cells and, in these cases, leads to increased activation of NSCs
[55][46]. The receptor for SDF-1, CXCR4, is also normally expressed in bone marrow, where both CXCR4 and SDF-1 are involved in hematopoietic stem cell mobilization and trafficking of NSCs
[56][76]. In a similar way, it is thought to mobilize and direct neuroblasts. The migration of NSCs in a rat stroke model was inhibited following the blocking of SDF-1 function using an antibody against CXCR4
[57][77]. SDF-1 may be one of the factors released by endothelial cells that leads to the directing of migrating neuroblasts to the vasculature
[55][46].
3.3. Monocyte Chemoattractant ProteinP-1
The (CC) family member chemokine, monocyte chemoattractant protein-1 (MCP-1), is known to interact with the CC-chemokine receptor-2 (CCR2). CCR2 is widely expressed on NSCs, and the binding of MCP-1 leads to increased NSC migration in vitro
[58][78]. It is suspected that this is accomplished through the receptor and ligand activating the PI3 kinase pathway
[59][79].
3.4. Matrix MetalloproteinaPses
Matrix metalloproteinases (MMPs) are known to have a role in cancer, promoting metastasis through the breakdown of the extracellular matrix (ECM). This then allows cells to migrate through tissue layers. This family of proteases also seems to play a role in the migration of SVZ neuroblasts. MMPs-3 and -9 are expressed in neuroblasts and their inhibition results in reduced neuroblast migration
[60][80]. MMPs-2 and -9 act on the vasculature to assist in the migration of neuroblasts by activating the PI3K/Akt and ERK1/2 signaling pathways
[57][77].
4. Therapeutic Targeting of the Interplay between Angiogenesis and Neurogenesis
Targeting angiogenesis for therapy in cancer is a well-established strategy that involves targeting the communication between tumor cells and the nearby blood vessels. It is known that interfering with neurogenic signaling can affect cancer development and progression. In mouse and tissue culture models of both prostate and lung cancer, it has been shown that chemical (6-hydroxydopamine, 6-OHDA) and surgical (hypogastric nerve cut) sympathectomy can prevent the development and progression of these cancers
[5][52][5,73]. Since it appears that angiogenesis is triggered by axonogenesis and neurogenesis through adrenergic signaling, targeting the adrenergic receptor or ligands is a viable treatment option. For instance, perineural invasion (PNI) is a promising therapeutic target. PNI is induced by the activation of the β2-adrenergic receptor, leading to PKA/STAT3 activation, which, in turn, activates NGF, MMP2, and MMP9 expression. Ligands which bind to and activate this receptor in PNI include sympathetic fiber-derived noradrenaline. This can be achieved using drugs, such as propranolol and penbutolol, which are β Adrenergic blockers, or atropine and hyoscine, which are muscarinic antagonists. Studies have shown that these drugs prevent prostate cancer cell migration
[61][138]. Dopamine receptors are also potential drug targets. In mouse models of lung cancer, Dopamine receptor D2 (D2R) agonist inhibits angiogenesis
[62][91]. Dopamine (DA) inhibitors can prevent cancer cell proliferation. These inhibitors lead to the downregulation of ERK1/2 and PI3K/AKT pathways
[63][139]. Both NGF and BDNF rely on Trk receptors to initiate the signaling pathways. The antagonism or inhibition of these Trks could potentially prevent neurotrophin signaling in cancer progression and initiation. This can be used to treat neuropathic pain related to cancer as well as inhibiting neurogenesis, angiogenesis, and their interplay. The Trk receptor inhibitors Larotrectinib and entrectinib have both been approved for use in the treatment of tumors
[64][140]. It has long been known that innervation of gastric tissue promotes the development of gastric cancers. The severing of the vagus nerve at particular branches has been found to inhibit the development of gastric cancers in a mouse model. In addition to this, similar results were achieved when the mice were treated with BOTOX, which acted to block neural signaling
[65][135].