Krüppel-like factors (KLFs) are transcription factors regulating various biological processes such as proliferation, differentiation, migration, invasion, and homeostasis. Importantly, they participate in disease development and progression. KLFs are expressed in multiple tissues, and their role is tissue- and context-dependent. KLF4 and KLF5 are two fascinating members of this family that regulate crucial stages of cellular identity from embryogenesis through differentiation and, finally, during tumorigenesis. They maintain homeostasis of various tissues and regulate inflammation, response to injury, regeneration, and development and progression of multiple cancers such as colorectal, breast, ovarian, pancreatic, lung, and prostate, to name a few.
- Krüppel-like factors
- intestine
- homeostasis
- colorectal cancer
1. Introduction
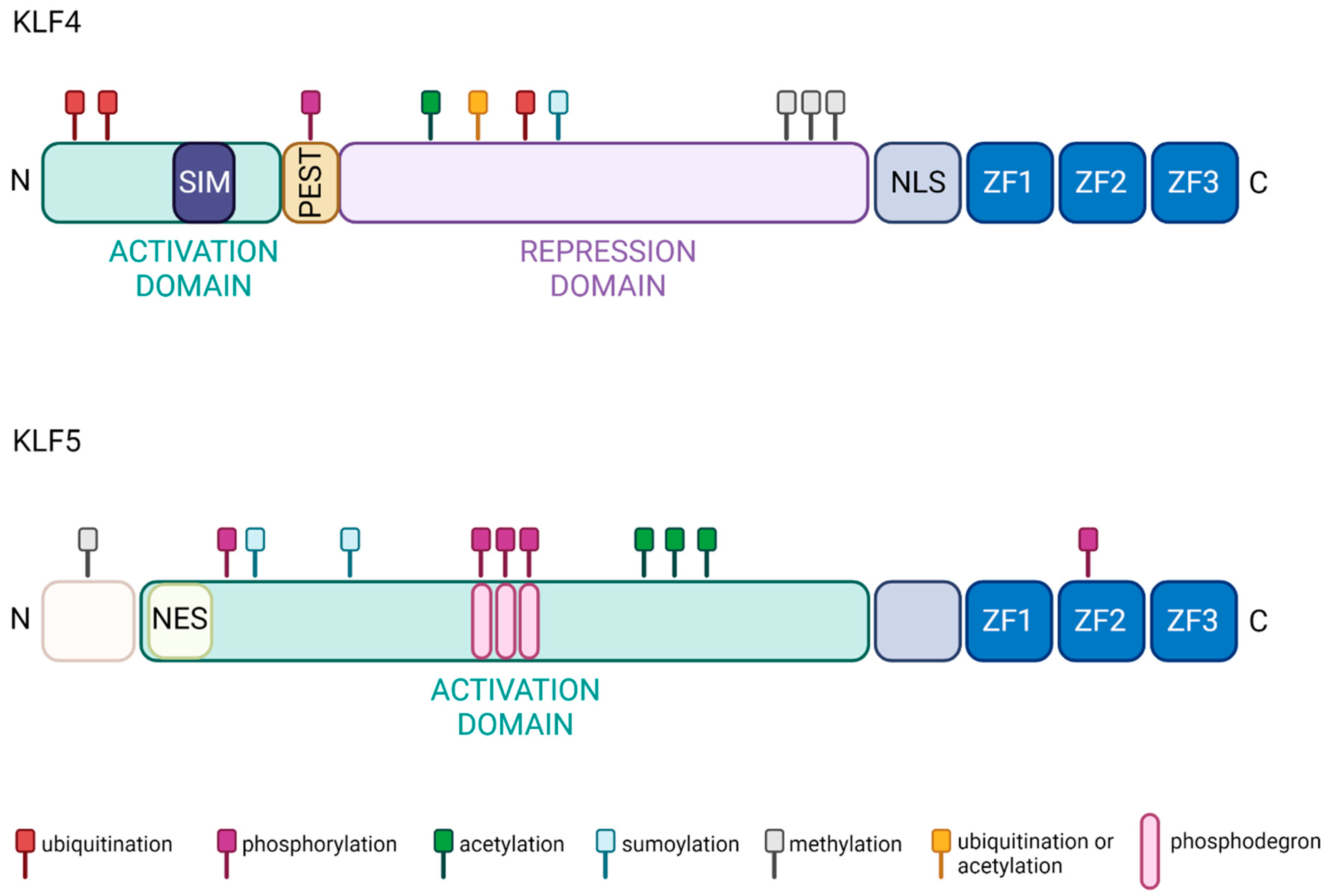
2. Krüppel-like Factor 4
2.1. Homeostasis
KLF4 is also called gut-enriched Krüppel-like factor (GKLF). KLF4 regulates transcription by modulating histone H4 acetylation at promoter sites [27]. An overview of its biochemical properties, regulation, and physiological functions was reviewed by us elsewhere [21]. KLF4 plays a significant role during gastrointestinal development and subsequent epithelial homeostasis. During murine fetal development, gastrointestinal KLF4 levels rise on embryonic day 13 and peak at day 17 [59][36]. By birth, KLF4 levels in colonic cells are typically higher than in small intestine cells. KLF4 levels persist throughout the gastrointestinal tract during life and rise with increasing age throughout adulthood [59][36]. Specifically, KLF4 is expressed in terminally differentiated epithelial cells at the mucosal villus border and reaches peak levels at terminal differentiation [60,61,62,63][37][38][39][40]. It is involved in goblet cell differentiation and maintenance and regulation of cell polarity [61][38]. Conditional ablation of Klf4 from the intestinal tract resulted in viable mice but with increased rates of epithelial proliferation and migration [64][41]. Partial depletion of KLF4 in terminally differentiated intestinal cells led to an increase in goblet cells, implying a role for KLF4 in maintaining goblet cell population and mispositioning of Paneth cells, suggesting KLF4-dependent localization [61][38]. In contrast, deletion of Klf4 from intestinal epithelial cells early in murine development led to a decrease in the number of colonic goblet cells in adult mice. This is due to differentiation failure, as KLF4 also plays a crucial role in negative regulation of the WNT pathway. Furthermore, Paneth cells were dislocated into the upper crypt due to reduced levels of Ephrin-B1, a KLF4 target [64,65][41][42]. Thus, KLF4 plays a vital role in differentiation and maintaining intestinal cell population and organization. Recent studies have shown that KLF4 regulates the proliferation status of a subpopulation of quiescent intestinal stem cells marked by the expression of B Lymphoma Mo-MLV Insertion Region 1 Homolog (Bmi1) [66][43]. Deletion of Klf4 from Bmi1-positive cells during homeostasis led to an increase in Bmi1-positive cell proliferation. In contrast, Bmi1-specific Klf4 deletion upon radiation injury reduces levels of Musashi-1 expression and inhibits crypt regeneration, demonstrating the context-dependent function of KLF4 [66,67][43][44]. Additionally, KLF4 plays an essential role in maintaining genetic stability, initiating apoptosis, and preventing epithelial–mesenchymal (EMT) transition in the progression of CRC. Murine cells that are absent in or have suppressed Klf4 expression demonstrate higher levels of genetic instability [68,69][45][46]. Next, wthe researchers discuss the contribution KLF4 has made towards the development, prognosis, and treatment of CRC, which will highlight KLF4′s role as both a tumor suppressor and oncogene.2.2. Colorectal Cancer
CRC has the third-highest annual incidence and second-highest mortality among men and women in the United States and worldwide [70][47]. Every year, an estimated 1.4 million new cases are reported worldwide. Of note, obesity, lack of physical activity, active and passive smoking, and high salt and red meat consumption have been established as risk factors for colorectal cancer [71][48]. While prevalence and mortality in those aged 50-and-older are declining due to early screening and improving therapies, the everyday nature of risk factors for colon cancer makes it a continued threat. Notably, the incidence of early-onset colorectal cancer in those aged 50 and younger has risen globally between 2.8–36.5% within the last 30 years [72,73,74,75][49][50][51][52]. Therefore, understanding how KLFs work in the context of colon cancer will be beneficial in preventing and treating colon cancer. Most CRC development follows a linear framework characterized by the adenoma–carcinoma–metastasis sequence [76][53]. The most well-studied mutations driving this sequence involve an initial suppression of Adenomatous polyposis coli (APC) followed by overexpression of Kirsten rat sarcoma viral oncogene homolog (KRAS) and loss of Tumor protein 53 (TP53) and Mothers against decapentaplegic homolog 4 (SMAD4) [77][54]. A review examining the role of KLF4, KLF5, and KLF6 in CRC was published by outhe researchers' group in 2008 [78][55]. The current revisewarch expands on the role of KLF4 and KLF5 in animal models of colorectal cancer, providing recent discoveries in their involvement in regulating the development, progression, and metastasis of CRC. In addition, wethe researchers described novel pathways that regulate KLF4 and KLF5 activity in CRC and summarized their role in the context of chemotherapy and radiation therapy and their potential as biomarkers of CRC. KLF4 is decreased in both adenomas from multiple intestinal neoplasia (ApcMin/+) mice and humans with familial adenomatous polyposis (FAP) when compared to either normal-appearing intestinal tissue from the same individual or healthy controls [79][56]. KLF4 has been shown to protect against the advancement of colitis into CRC via increased genetic stability in murine models [80][57]. Immuno-stains of normal colon show a gradient in KLF4 concentration that is the highest near the surface epithelium and lowest towards the crypt [81][58]. This gradient is disrupted in adenomas and carcinomas [81][58]. Loss of heterozygosity in KLF4 and hypermethylation at its 5′-untranslated region are common in CRC [82][59]. KLF4 primarily contributes to early CRC development and is associated with EMT in CRC [83,84][60][61]. The discussion here focuses on KLF4 in solid tumor CRC.3. Krüppel-like Factor 5
3.1. Homeostasis
KLF5 is also known as Intestinal Krüppel-like factor (IKLF) due to its high expression in intestinal epithelium. However, KLF5 can be detected in almost all tissues, including breast, prostate, pancreas, intestine, lung, bladder, and skeletal muscle [22,32,60,149,150,151][22][32][37][62][63][64]. KLF5 regulates many cellular processes, including cell cycle, proliferation, migration, invasion, stemness, apoptosis, and autophagy, and plays a crucial role in maintaining gut homeostasis [32,152,153,154,155,156,157,158,159,160,161,162,163][32][65][66][67][68][69][70][71][72][73][74][75][76]. Notably, KLF5 regulates villus formation and initiates cytodifferentiation in embryonic intestinal epithelium. Deletion of Klf5 from intestinal epithelium during embryogenesis leads to downregulation of multiple genes such as E74-like ETS transcription factor 3 (Elf3), Pparg, Atonal BHLH transcription factor 1 (Atoh1), Achaete-scute family bHLH transcription factor 2 (Ascl2), Hepatocyte nuclear factor 4 alpha (Hnf4a), Neurogenin 3 (Neurog3), and Caudal Type Homeobox 1 (Cdx1) [164][77]. Similarly, data obtained from Klf5 deletion in the gut suggest that KLF5 plays a role in maintaining epithelial proliferation, differentiation, and cell positioning along the crypt radial axis in adult mice [165,166][78][79]. Mice with deletion of Klf5 within active intestinal epithelial stem cells have decreased expression of intestinal stem cell signature genes, such as Lgr5, Olfactomedin 4 (Olfm4), and Ascl2, and impaired stem cell renewal. KLF5 is crucial for stem cell activity and regeneration of the intestinal epithelium after injury [167,168][80][81]. KLF5 also regulates DNA damage repair in intestinal epithelial cells upon radiation injury. In mice with heterozygous deletion of Klf5 in intestinal epithelial cells, genes involved in nucleotide excision repair, mismatch repair, and non-homologous end-joining were significantly downregulated compared to wild-type mice [169][82]. Mice with intestinal epithelium-specific deletion of Klf5 also developed a Th-17-mediated immune response and subsequent colitis, suggesting a protective role of KLF5 against intestinal inflammation [170][83]. Evidently, KLF5 is indicated in a wide range of processes to ensure intestinal epithelial homeostasis in the presence of insults. While the lack of KLF5 activity can lead to insufficient self-renewal and intestinal integrity, overactivation of KLF5 may cause uncontrolled cell proliferation and differentiation, ultimately leading to tumorigenesis. As such, understanding the role of KLF5 in achieving balance in these cellular processes is essential to ensure intestinal health. However, whether KLF5 functions to upregulate or downregulate these processes is context-dependent and highly controversial.3.2. Colorectal Cancer
3.2.1. KLF5 Is a Pro-Proliferative Factor in CRC
KLF5 is a pro-proliferative transcription factor downstream of the classical Mitogen-activated protein kinase (MAPK-ERK-RAS) pathway and directly regulated by Early Growth Response 1 (EGR1) [171][84]. Activation of the KRAS oncogene plays an essential role in CRC pathophysiology, and KLF5 contributes to colorectal tumorigenesis induced by a constitutively activating KRAS mutation (G12V) (Figure 2). For example, Klf5 haploinsufficiency in ApcMin/+/KrasG12V mice resulted in significantly reduced tumor number and size compared to ApcMin/+ mice [172][85]. In addition, increased levels of KLF5 were observed in spontaneous hyperplastic intestinal polyp development and colonic tumorigenesis in Villin-Cre/LSL-KRASG12D mice, further supporting KLF5′s role as a mediator of the KRAS pathway in CRC formation [173][86]. Interestingly, while the Villin-Cre/LSL-KRASG12D mice displayed decreased survival when treated with AOM compared to controls, loss of one Klf5 allele showed reduced levels of KRAS effector proteins and, as a result, reduced mortality upon AOM treatment [173][86]. Overall, KLF5 expression appears essential in exerting the oncogenic, pro-proliferative effects of KRAS mutations in CRC.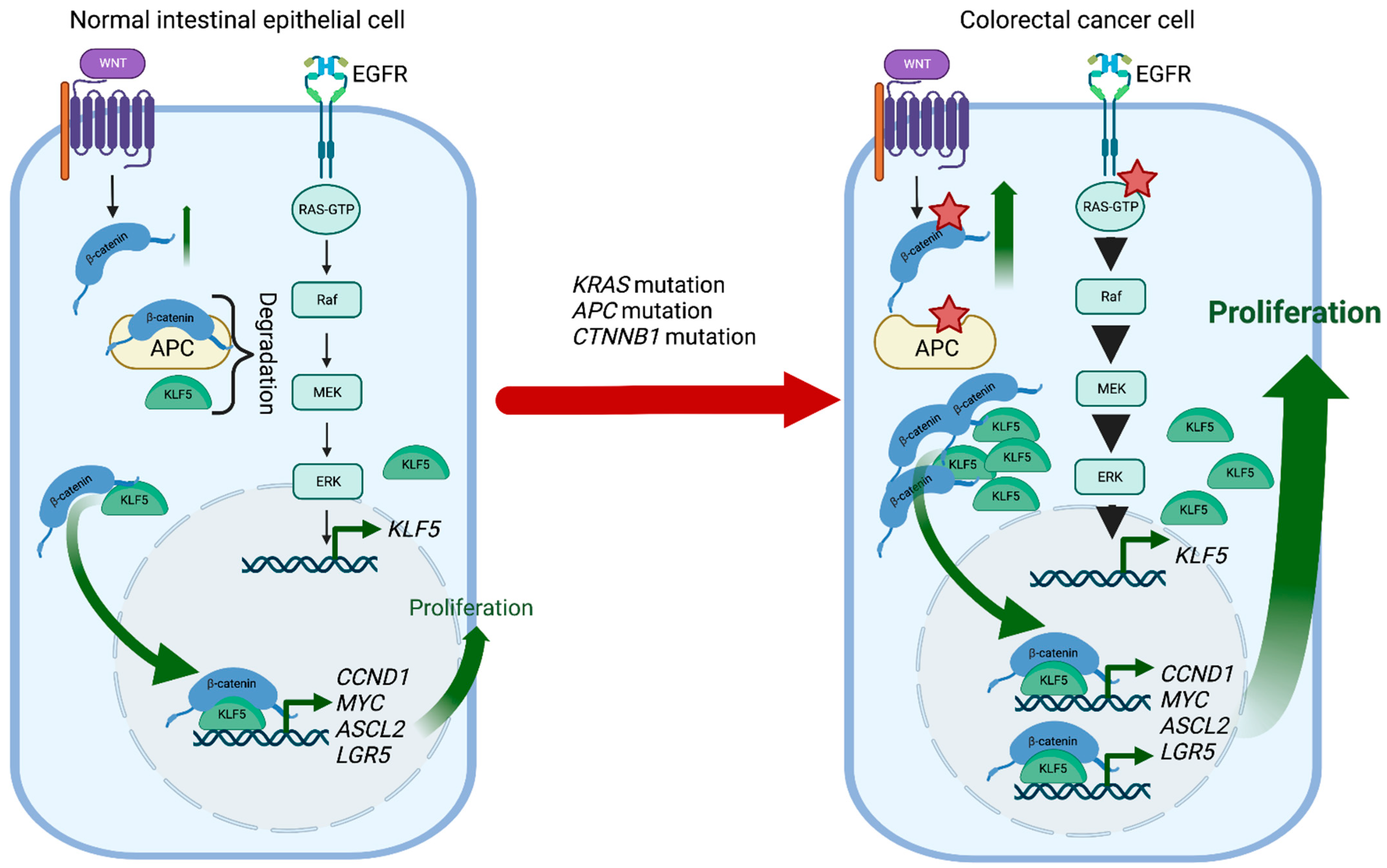
3.2.2. KLF5-WNT/β-Catenin Positive Feedback Loop Regulated CRC Development and Progression
Germline loss-of-function mutation of APC and mutations in CTNNB1 have been identified as a cause of colorectal cancer. APC plays an essential role in regulating the activity of β-catenin, which controls the WNT signaling pathway responsible for maintaining the proliferation of the intestinal crypt epithelium. KLF5 is a crucial mediator of these interactions contributing to CRC tumorigenesis. Klf5 haploinsufficiency in the context of Apc mutation was associated with lower levels and reduced nuclear localization of β-catenin, resulting in reduced expression of Ccnd1 and c-Myc, downregulation of the WNT pathway activity, and decreased polyp formation [177][90]. In addition, the formation of lethal colorectal adenomas and carcinomas induced by β-catenin mutations in Lgr5+ stem cells was entirely suppressed by Klf5 deletion [178][91]. Overall, lack of KLF5 expression prevented the tumorigenic effects of Apc mutation and β-catenin activation, suggesting the oncogenic function and necessity of KLF5 in CRC (Figure 2). Lysophosphatidic acid (LPA), a simple phospholipid with potent mitogenic effects, and its receptor LPAR modulate the tumorigenic effects of APC mutation. Compared to ApcMin/+ mice, ApcMin/+/Lpar2−/− mice exhibited decreased tumor progression and hypoxia in response to reduced expression of Klf5, Ctnnb1, Ccnd1 and c-Myc [179][92]. A recent study proposed a new mechanism by which KLF5 modulates the WNT/β-catenin pathway in the presence of LPA. Contrary to previous findings, silencing KLF5 did not alter the nuclear translocation of β-catenin by LPA. Instead, KLF5 was found to facilitate LPA-induced formation and transcriptional activity of the β-catenin/TCF complex to promote colon cancer cell proliferation [180][93]. Ketogenesis is significantly decreased in the tumor microenvironment of CRC. As such, a ketogenic diet of high lipids and low carbohydrates has been recommended for cancer patients. Increasing ketogenesis markedly decreased KLF5-dependent synthesis of C-X-C Motif Chemokine Ligand 12 (CXCL12) in cancer-associated fibroblasts, ultimately increasing the infiltration of immune effector cells in tumors and enhancing sensitivity to immune checkpoint inhibitors specific for programmed cell death 1 (PD-1) [181][94]. By the same mechanism, increasing ketogenesis inhibited CRC migration, invasion, and metastasis both in vitro and in vivo [182][95].3.2.3. KLF5 and microRNA in CRC
3.2.4. KLF5 as a Therapeutic Target in CRC
Using an ultra-high-throughput screen, outhe researchers' group identified two KLF5-selective compounds, CID 439501 and 5951923, that significantly decrease endogenous KLF5 protein levels and reduce the viability of several CRC cell lines [191][103]. A small-molecule compound called ML264 was found to be a KLF5 inhibitor, preventing the expression of KLF5 and the growth of CRC xenograft tumors [192][104]. ML264 exerted this effect by inhibiting the RAS/MAPK/PI3K and the WNT/β-catenin signaling pathway. The same KLF5 inhibitor was recently used to investigate CRC resistance to oxaliplatin, a first-line chemotherapy drug commonly used in CRC. Using ML264, the study successfully inhibited the KLF5/BCL-2/Caspase 3 signaling pathway, thereby restoring the apoptotic response and significantly restoring sensitivity to oxaliplatin in CRC patient-derived organoids [193][105]. Interestingly, SR18662, a derivative of ML264, demonstrated enhanced abilities to inhibit KLF5, the MAPK and WNT pathways, and the growth of CRC in vitro and in vivo with the ability to exert cytotoxic effects [194][106]. Dual-specificity phosphatase 10 (DUSP10), known for its role in deactivating MAP kinases, reduced intestinal epithelial cell proliferation via inhibition of ERK1/2 activation and KLF5 expression [195][107]. KLF5 also modulates CRC response to radiation therapy. For example, HCT116 cells with significantly higher levels of KLF5 were shown to increase CyclinD1 and β-catenin and promote better cell viability than control cells when subjected to radiation therapy [199][108]. In addition, the depletion of KLF5 in HCT116 cells increased CRC sensitivity to DNA-damaging ultraviolet irradiation therapy by failing to induce the proto-oncogene, serine/threonine kinase 1 (PIM1) survival kinase [200][109]. It appears that overexpression of KLF5 confers resistance to radiotherapy, while reduction of KLF5 may increase susceptibility to radiotherapeutic effects in CRC.3.2.5. KLF5 as a Biomarker of CRC
Overexpression of KLF5 may be used as a predictive biomarker for poor tumor regression after preoperative chemoradiation therapy, the standard treatment for locally advanced rectal cancer [199][108]. A recent study was the first to examine the expression of levels of KLF5 in patients with colorectal cancer to determine correlation with clinical outcomes. The study revealed that high expression of KLF5 in tissues collected from CRC patients was associated with vascular invasion, increased serum carbohydrate 19-9, larger metastatic liver tumors, and poorer prognosis after surgery. While further investigation is needed, KLF5 upregulation of c-MYC and CCND1 via promoter binding may be the mechanism underlining these effects. Thus, high KLF5 expression can independently predict poor prognosis in patients with primary CRC and liver metastasis [201][110]. However, KLF5 and its use as a prognosis marker in CRC must be studied further.References
- Pearson, R.; Fleetwood, J.; Eaton, S.; Crossley, M.; Bao, S. Kruppel-like transcription factors: A functional family. Int. J. Biochem. Cell Biol. 2008, 40, 1996–2001.
- Dang, D.T.; Pevsner, J.; Yang, V.W. The biology of the mammalian Kruppel-like family of transcription factors. Int. J. BioChem. Cell Biol. 2000, 32, 1103–1121.
- Kaczynski, J.; Cook, T.; Urrutia, R. Sp1- and Kruppel-like transcription factors. Genome Biol. 2003, 4, 206.
- Abe, M.; Saeki, N.; Ikeda, Y.; Ohba, S. Kruppel-like Factors in Skeletal Physiology and Pathologies. Int. J. Mol. Sci. 2022, 23, 15174.
- Surkova, S.; Golubkova, E.; Mamon, L.; Samsonova, M. Dynamic maternal gradients and morphogenetic networks in Drosophila early embryo. Biosystems 2018, 173, 207–213.
- Ghaleb, A.M.; Nandan, M.O.; Chanchevalap, S.; Dalton, W.B.; Hisamuddin, I.M.; Yang, V.W. Kruppel-like factors 4 and 5: The yin and yang regulators of cellular proliferation. Cell Res. 2005, 15, 92–96.
- Suzuki, T.; Aizawa, K.; Matsumura, T.; Nagai, R. Vascular implications of the Kruppel-like family of transcription factors. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 1135–1141.
- Haldar, S.M.; Ibrahim, O.A.; Jain, M.K. Kruppel-like Factors (KLFs) in muscle biology. J. Mol. Cell Cardiol. 2007, 43, 1–10.
- Nandan, M.O.; Yang, V.W. The role of Kruppel-like factors in the reprogramming of somatic cells to induced pluripotent stem cells. Histol. Histopathol. 2009, 24, 1343–1355.
- Brey, C.W.; Nelder, M.P.; Hailemariam, T.; Gaugler, R.; Hashmi, S. Kruppel-like family of transcription factors: An emerging new frontier in fat biology. Int. J. Biol. Sci. 2009, 5, 622–636.
- Cao, Z.; Sun, X.; Icli, B.; Wara, A.K.; Feinberg, M.W. Role of Kruppel-like factors in leukocyte development, function, and disease. Blood 2010, 116, 4404–4414.
- Moore, D.L.; Apara, A.; Goldberg, J.L. Kruppel-like transcription factors in the nervous system: Novel players in neurite outgrowth and axon regeneration. Mol. Cell Neurosci. 2011, 47, 233–243.
- Wu, Z.; Wang, S. Role of kruppel-like transcription factors in adipogenesis. Dev. Biol. 2013, 373, 235–243.
- Mallipattu, S.K.; Estrada, C.C.; He, J.C. The critical role of Kruppel-like factors in kidney disease. Am. J. Physiol. Renal. Physiol. 2017, 312, F259–F265.
- Kim, C.K.; He, P.; Bialkowska, A.B.; Yang, V.W. SP and KLF Transcription Factors in Digestive Physiology and Diseases. Gastroenterology 2017, 152, 1845–1875.
- Oishi, Y.; Manabe, I. Kruppel-Like Factors in Metabolic Homeostasis and Cardiometabolic Disease. Front. Cardiovasc. Med. 2018, 5, 69.
- McConnell, B.B.; Yang, V.W. Mammalian Kruppel-like factors in health and diseases. Physiol. Rev. 2010, 90, 1337–1381.
- Prosdocimo, D.A.; Sabeh, M.K.; Jain, M.K. Kruppel-like factors in muscle health and disease. Trends Cardiovasc. Med. 2015, 25, 278–287.
- Orzechowska-Licari, E.J.; LaComb, J.F.; Mojumdar, A.; Bialkowska, A.B. SP and KLF Transcription Factors in Cancer Metabolism. Int. J. Mol. Sci. 2022, 23, 9956.
- Palioura, D.; Lazou, A.; Drosatos, K. Kruppel-like factor (KLF)5: An emerging foe of cardiovascular health. J. Mol. Cell Cardiol. 2022, 163, 56–66.
- Ghaleb, A.M.; Yang, V.W. Kruppel-like factor 4 (KLF4): What we currently know. Gene 2017, 611, 27–37.
- Luo, Y.; Chen, C. The roles and regulation of the KLF5 transcription factor in cancers. Cancer Sci. 2021, 112, 2097–2117.
- Vangapandu, H.; Ai, W. Kruppel like factor 4 (KLF4): A transcription factor with diverse context-dependent functions. Gene Ther. Mol. Biol. 2009, 13, 194–204.
- Swamynathan, S.K. Kruppel-like factors: Three fingers in control. Hum. Genom. 2010, 4, 263–270.
- Hu, D.; Gur, M.; Zhou, Z.; Gamper, A.; Hung, M.C.; Fujita, N.; Lan, L.; Bahar, I.; Wan, Y. Interplay between arginine methylation and ubiquitylation regulates KLF4-mediated genome stability and carcinogenesis. Nat. Commun. 2015, 6, 8419.
- Du, J.X.; McConnell, B.B.; Yang, V.W. A small ubiquitin-related modifier-interacting motif functions as the transcriptional activation domain of Kruppel-like factor 4. J. Biol. Chem. 2010, 285, 28298–28308.
- Evans, P.M.; Zhang, W.; Chen, X.; Yang, J.; Bhakat, K.K.; Liu, C. Kruppel-like factor 4 is acetylated by p300 and regulates gene transcription via modulation of histone acetylation. J. Biol. Chem. 2007, 282, 33994–34002.
- Kim, M.O.; Kim, S.H.; Cho, Y.Y.; Nadas, J.; Jeong, C.H.; Yao, K.; Kim, D.J.; Yu, D.H.; Keum, Y.S.; Lee, K.Y.; et al. ERK1 and ERK2 regulate embryonic stem cell self-renewal through phosphorylation of Klf4. Nat. Struct. Mol. Biol. 2012, 19, 283–290.
- Ray, S.K. The Transcription Regulator Kruppel-Like Factor 4 and Its Dual Roles of Oncogene in Glioblastoma and Tumor Suppressor in Neuroblastoma. For. Immunopathol. Dis. Therap. 2016, 7, 127–139.
- Lim, K.H.; Kim, S.R.; Ramakrishna, S.; Baek, K.H. Critical lysine residues of Klf4 required for protein stabilization and degradation. Biochem. Biophys. Res. Commun. 2014, 443, 1206–1210.
- Rane, M.J.; Zhao, Y.; Cai, L. Krupsilonppel-like factors (KLFs) in renal physiology and disease. EBioMedicine 2019, 40, 743–750.
- Dong, J.T.; Chen, C. Essential role of KLF5 transcription factor in cell proliferation and differentiation and its implications for human diseases. Cell Mol. Life Sci. 2009, 66, 2691–2706.
- Li, J.; Liu, L.; Zhou, W.Q.; Cai, L.; Xu, Z.G.; Rane, M.J. Roles of Kruppel-like factor 5 in kidney disease. J. Cell Mol. Med. 2021, 25, 2342–2355.
- Luan, Y.; Wang, P. FBW7-mediated ubiquitination and degradation of KLF5. World J. Biol. Chem. 2014, 5, 216–223.
- Bialkowska, A.B.; Liu, Y.; Nandan, M.O.; Yang, V.W. A colon cancer-derived mutant of Kruppel-like factor 5 (KLF5) is resistant to degradation by glycogen synthase kinase 3beta (GSK3beta) and the E3 ubiquitin ligase F-box and WD repeat domain-containing 7alpha (FBW7alpha). J. Biol. Chem. 2014, 289, 5997–6005.
- Ton-That, H.; Kaestner, K.H.; Shields, J.M.; Mahatanankoon, C.S.; Yang, V.W. Expression of the gut-enriched Kruppel-like factor gene during development and intestinal tumorigenesis. FEBS Lett. 1997, 419, 239–243.
- McConnell, B.B.; Ghaleb, A.M.; Nandan, M.O.; Yang, V.W. The diverse functions of Kruppel-like factors 4 and 5 in epithelial biology and pathobiology. Bioessays 2007, 29, 549–557.
- Yu, T.; Chen, X.; Zhang, W.; Li, J.; Xu, R.; Wang, T.C.; Ai, W.; Liu, C. Kruppel-like factor 4 regulates intestinal epithelial cell morphology and polarity. PLoS ONE 2012, 7, e32492.
- Xu, J.; Lu, B.; Xu, F.; Gu, H.; Fang, Y.; Huang, Q.; Lai, M. Dynamic down-regulation of Kruppel-like factor 4 in colorectal adenoma-carcinoma sequence. J. Cancer Res. Clin. Oncol. 2008, 134, 891–898.
- Flandez, M.; Guilmeau, S.; Blache, P.; Augenlicht, L.H. KLF4 regulation in intestinal epithelial cell maturation. Exp. Cell Res. 2008, 314, 3712–3723.
- Ghaleb, A.M.; McConnell, B.B.; Kaestner, K.H.; Yang, V.W. Altered intestinal epithelial homeostasis in mice with intestine-specific deletion of the Kruppel-like factor 4 gene. Dev. Biol. 2011, 349, 310–320.
- Evans, P.M.; Chen, X.; Zhang, W.; Liu, C. KLF4 interacts with beta-catenin/TCF4 and blocks p300/CBP recruitment by beta-catenin. Mol. Cell Biol. 2010, 30, 372–381.
- Kuruvilla, J.G.; Kim, C.K.; Ghaleb, A.M.; Bialkowska, A.B.; Kuo, C.J.; Yang, V.W. Kruppel-like Factor 4 Modulates Development of BMI1(+) Intestinal Stem Cell-Derived Lineage Following gamma-Radiation-Induced Gut Injury in Mice. Stem Cell Rep. 2016, 6, 815–824.
- Orzechowska, E.J.; Katano, T.; Bialkowska, A.B.; Yang, V.W. Interplay among p21(Waf1/Cip1), MUSASHI-1 and Kruppel-like factor 4 in activation of Bmi1-Cre(ER) reserve intestinal stem cells after gamma radiation-induced injury. Sci. Rep. 2020, 10, 18300.
- Hagos, E.G.; Ghaleb, A.M.; Dalton, W.B.; Bialkowska, A.B.; Yang, V.W. Mouse embryonic fibroblasts null for the Kruppel-like factor 4 gene are genetically unstable. Oncogene 2009, 28, 1197–1205.
- El-Karim, E.A.; Hagos, E.G.; Ghaleb, A.M.; Yu, B.; Yang, V.W. Kruppel-like factor 4 regulates genetic stability in mouse embryonic fibroblasts. Mol. Cancer 2013, 12, 89.
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics, 2022. CA Cancer J. Clin. 2022, 72, 7–33.
- Lewandowska, A.; Rudzki, G.; Lewandowski, T.; Stryjkowska-Gora, A.; Rudzki, S. Title: Risk Factors for the Diagnosis of Colorectal Cancer. Cancer Control. 2022, 29, 10732748211056692.
- Ahmedin Jemal, R.S.; Tyler, B.K. Colorectal Cancer Facts and FiguRes. 2023–2025. CA Cancer J. Clin. 2023. Available online: https://www.cancer.org/research/cancer-facts-statistics/colorectal-cancer-facts-figures.html (accessed on 9 March 2023).
- Ganapathi, S.; Kumar, D.; Katsoulas, N.; Melville, D.; Hodgson, S.; Finlayson, C.; Hagger, R. Colorectal cancer in the young: Trends, characteristics and outcome. Int. J. Colorectal Dis. 2011, 26, 927–934.
- Loomans-Kropp, H.A.; Umar, A. Increasing Incidence of Colorectal Cancer in Young Adults. J. Cancer Epidemiol. 2019, 2019, 9841295.
- Vuik, F.E.; Nieuwenburg, S.A.; Bardou, M.; Lansdorp-Vogelaar, I.; Dinis-Ribeiro, M.; Bento, M.J.; Zadnik, V.; Pellise, M.; Esteban, L.; Kaminski, M.F.; et al. Increasing incidence of colorectal cancer in young adults in Europe over the last 25 years. Gut 2019, 68, 1820–1826.
- Li, J.; Ma, X.; Chakravarti, D.; Shalapour, S.; DePinho, R.A. Genetic and biological hallmarks of colorectal cancer. Genes Dev. 2021, 35, 787–820.
- Smit, W.L.; Spaan, C.N.; Johannes de Boer, R.; Ramesh, P.; Martins Garcia, T.; Meijer, B.J.; Vermeulen, J.L.M.; Lezzerini, M.; MacInnes, A.W.; Koster, J.; et al. Driver mutations of the adenoma-carcinoma sequence govern the intestinal epithelial global translational capacity. Proc. Natl. Acad. Sci. USA 2020, 117, 25560–25570.
- Ghaleb, A.M.; Yang, V.W. The Pathobiology of Kruppel-like Factors in Colorectal Cancer. Curr. Colorectal Cancer Rep. 2008, 4, 59–64.
- Dang, D.T.; Bachman, K.E.; Mahatan, C.S.; Dang, L.H.; Giardiello, F.M.; Yang, V.W. Decreased expression of the gut-enriched Kruppel-like factor gene in intestinal adenomas of multiple intestinal neoplasia mice and in colonic adenomas of familial adenomatous polyposis patients. FEBS Lett. 2000, 476, 203–207.
- Yang, V.W.; Liu, Y.; Kim, J.; Shroyer, K.R.; Bialkowska, A.B. Increased Genetic Instability and Accelerated Progression of Colitis-Associated Colorectal Cancer through Intestinal Epithelium-specific Deletion of Klf4. Mol. Cancer Res. 2019, 17, 165–176.
- Shie, J.L.; Chen, Z.Y.; O’Brien, M.J.; Pestell, R.G.; Lee, M.E.; Tseng, C.C. Role of gut-enriched Kruppel-like factor in colonic cell growth and differentiation. Am. J. Physiol. Gastrointest. Liver Physiol. 2000, 279, G806–G814.
- Zhao, W.; Hisamuddin, I.M.; Nandan, M.O.; Babbin, B.A.; Lamb, N.E.; Yang, V.W. Identification of Kruppel-like factor 4 as a potential tumor suppressor gene in colorectal cancer. Oncogene 2004, 23, 395–402.
- Agbo, K.C.; Huang, J.Z.; Ghaleb, A.M.; Williams, J.L.; Shroyer, K.R.; Bialkowska, A.B.; Yang, V.W. Loss of the Kruppel-like factor 4 tumor suppressor is associated with epithelial-mesenchymal transition in colorectal cancer. J. Cancer Metastasis Treat. 2019, 5, 77.
- Choi, B.J.; Cho, Y.G.; Song, J.W.; Kim, C.J.; Kim, S.Y.; Nam, S.W.; Yoo, N.J.; Lee, J.Y.; Park, W.S. Altered expression of the KLF4 in colorectal cancers. Pathol. Res. Pract. 2006, 202, 585–589.
- Li, J.C.; Chen, Q.H.; Jian, R.; Zhou, J.R.; Xu, Y.; Lu, F.; Li, J.Q.; Zhang, H. The Partial Role of KLF4 and KLF5 in Gastrointestinal Tumors. Gastroenterol. Res. Pract. 2021, 2021, 2425356.
- Diakiw, S.M.; D’Andrea, R.J.; Brown, A.L. The double life of KLF5: Opposing roles in regulation of gene-expression, cellular function, and transformation. IUBMB Life 2013, 65, 999–1011.
- Simmen, R.C.; Pabona, J.M.; Velarde, M.C.; Simmons, C.; Rahal, O.; Simmen, F.A. The emerging role of Kruppel-like factors in endocrine-responsive cancers of female reproductive tissues. J. Endocrinol. 2010, 204, 223–231.
- Jia, L.; Zhou, Z.; Liang, H.; Wu, J.; Shi, P.; Li, F.; Wang, Z.; Wang, C.; Chen, W.; Zhang, H.; et al. KLF5 promotes breast cancer proliferation, migration and invasion in part by upregulating the transcription of TNFAIP2. Oncogene 2016, 35, 2040–2051.
- Ma, D.; Chang, L.Y.; Zhao, S.; Zhao, J.J.; Xiong, Y.J.; Cao, F.Y.; Yuan, L.; Zhang, Q.; Wang, X.Y.; Geng, M.L.; et al. KLF5 promotes cervical cancer proliferation, migration and invasion in a manner partly dependent on TNFRSF11a expression. Sci. Rep. 2017, 7, 15683.
- Yang, Y.; Tetreault, M.P.; Yermolina, Y.A.; Goldstein, B.G.; Katz, J.P. Kruppel-like factor 5 controls keratinocyte migration via the integrin-linked kinase. J. Biol. Chem. 2008, 283, 18812–18820.
- Du, C.; Gao, Y.; Xu, S.; Jia, J.; Huang, Z.; Fan, J.; Wang, X.; He, D.; Guo, P. KLF5 promotes cell migration by up-regulating FYN in bladder cancer cells. FEBS Lett. 2016, 590, 408–418.
- Kim, C.K.; Saxena, M.; Maharjan, K.; Song, J.J.; Shroyer, K.R.; Bialkowska, A.B.; Shivdasani, R.A.; Yang, V.W. Kruppel-like Factor 5 Regulates Stemness, Lineage Specification, and Regeneration of Intestinal Epithelial Stem Cells. Cell Mol. Gastroenterol. Hepatol. 2020, 9, 587–609.
- Takeda, T.; Yokoyama, Y.; Takahashi, H.; Okuzaki, D.; Asai, K.; Itakura, H.; Miyoshi, N.; Kobayashi, S.; Uemura, M.; Fujita, T.; et al. A stem cell marker KLF5 regulates CCAT1 via three-dimensional genome structure in colorectal cancer cells. Br. J. Cancer 2022, 126, 109–119.
- Wei, X.; Ye, J.; Shang, Y.; Chen, H.; Liu, S.; Liu, L.; Wang, R. Ascl2 activation by YAP1/KLF5 ensuRes. the self-renewability of colon cancer progenitor cells. Oncotarget 2017, 8, 109301–109318.
- Kinisu, M.; Choi, Y.J.; Cattoglio, C.; Liu, K.; Roux de Bezieux, H.; Valbuena, R.; Pum, N.; Dudoit, S.; Huang, H.; Xuan, Z.; et al. Klf5 establishes bi-potential cell fate by dual regulation of ICM and TE specification genes. Cell Rep. 2021, 37, 109982.
- Azami, T.; Matsumoto, K.; Jeon, H.; Waku, T.; Muratani, M.; Niwa, H.; Takahashi, S.; Ema, M. Klf5 suppresses ERK signaling in mouse pluripotent stem cells. PLoS ONE 2018, 13, e0207321.
- Dai, X.; Chen, Y.; Chen, N.; Dou, J.; Zhuang, H.; Wang, J.; Zhao, X.; Zhang, X.; Zhao, H. KLF5-mediated aquaporin 3 activated autophagy to facilitate cisplatin resistance of gastric cancer. Immunopharmacol. Immunotoxicol. 2022, 45, 1–13.
- Wang, Z.; Yang, L.; Wu, P.; Li, X.; Tang, Y.; Ou, X.; Zhang, Y.; Xiao, X.; Wang, J.; Tang, H. The circROBO1/KLF5/FUS feedback loop regulates the liver metastasis of breast cancer by inhibiting the selective autophagy of afadin. Mol. Cancer 2022, 21, 29.
- Cheng, Z.; Liu, G.; Huang, C.; Zhao, X. KLF5 activates lncRNA DANCR and inhibits cancer cell autophagy accelerating gastric cancer progression. NPJ Genom. Med. 2021, 6, 75.
- Bell, S.M.; Zhang, L.; Xu, Y.; Besnard, V.; Wert, S.E.; Shroyer, N.; Whitsett, J.A. Kruppel-like factor 5 controls villus formation and initiation of cytodifferentiation in the embryonic intestinal epithelium. Dev. Biol. 2013, 375, 128–139.
- McConnell, B.B.; Kim, S.S.; Yu, K.; Ghaleb, A.M.; Takeda, N.; Manabe, I.; Nusrat, A.; Nagai, R.; Yang, V.W. Kruppel-like factor 5 is important for maintenance of crypt architecture and barrier function in mouse intestine. Gastroenterology 2011, 141, 1302–1313.e6.
- Bell, K.N.; Shroyer, N.F. Krupple-like factor 5 is required for proper maintenance of adult intestinal crypt cellular proliferation. Dig. Dis. Sci. 2015, 60, 86–100.
- Nandan, M.O.; Ghaleb, A.M.; Bialkowska, A.B.; Yang, V.W. Kruppel-like factor 5 is essential for proliferation and survival of mouse intestinal epithelial stem cells. Stem Cell Res. 2015, 14, 10–19.
- Kuruvilla, J.G.; Ghaleb, A.M.; Bialkowska, A.B.; Nandan, M.O.; Yang, V.W. Role of Kruppel-like factor 5 in the maintenance of the stem cell niche in the intestinal crypt. Stem. Cell Transl. Investig. 2015, 2, e839.
- Li, M.; Gu, Y.; Ma, Y.C.; Shang, Z.F.; Wang, C.; Liu, F.J.; Cao, J.P.; Wan, H.J.; Zhang, X.G. Kruppel-Like Factor 5 Promotes Epithelial Proliferation and DNA Damage Repair in the Intestine of Irradiated Mice. Int. J. Biol. Sci. 2015, 11, 1458–1468.
- Shieh, J.; Chu, T.H.; Liu, Y.; Kim, J.; Ruiz de Sabando, A.; Kobayashi, S.; Zee, S.Y.; Sheridan, B.S.; Bialkowska, A.B.; Yang, V.W. KLF5 protects the intestinal epithelium against Th17 immune response in a murine colitis model. JCI Insight 2022, 7, e153488.
- Nandan, M.O.; Yoon, H.S.; Zhao, W.; Ouko, L.A.; Chanchevalap, S.; Yang, V.W. Kruppel-like factor 5 mediates the transforming activity of oncogenic H-Ras. Oncogene 2004, 23, 3404–3413.
- Nandan, M.O.; Ghaleb, A.M.; McConnell, B.B.; Patel, N.V.; Robine, S.; Yang, V.W. Kruppel-like factor 5 is a crucial mediator of intestinal tumorigenesis in mice harboring combined ApcMin and KRASV12 mutations. Mol. Cancer 2010, 9, 63.
- Nandan, M.O.; Bialkowska, A.B.; Yang, V.W. KLF5 mediates the hyper-proliferative phenotype of the intestinal epithelium in mice with intestine-specific endogenous K-Ras(G12D) expression. Am. J. Cancer Res. 2018, 8, 723–731.
- OuYang, C.; Xie, Y.; Fu, Q.; Xu, G. SYNPO2 suppresses hypoxia-induced proliferation and migration of colorectal cancer cells by regulating YAP-KLF5 axis. Tissue Cell 2021, 73, 101598.
- Siraj, A.K.; Pratheeshkumar, P.; Divya, S.P.; Parvathareddy, S.K.; Bu, R.; Masoodi, T.; Kong, Y.; Thangavel, S.; Al-Sanea, N.; Ashari, L.H.; et al. TGFbeta-induced SMAD4-dependent Apoptosis Proceeded by EMT in CRC. Mol. Cancer Ther. 2019, 18, 1312–1322.
- Xu, Z.; Gao, H.; Zhang, Y.; Feng, W.; Miao, Y.; Xu, Z.; Li, W.; Chen, F.; Lv, Z.; Huo, J.; et al. CCL7 and TGF-beta secreted by MSCs play opposite roles in regulating CRC metastasis in a KLF5/CXCL5-dependent manner. Mol. Ther. 2022, 30, 2327–2341.
- McConnell, B.B.; Bialkowska, A.B.; Nandan, M.O.; Ghaleb, A.M.; Gordon, F.J.; Yang, V.W. Haploinsufficiency of Kruppel-like factor 5 rescues the tumor-initiating effect of the Apc(Min) mutation in the intestine. Cancer Res. 2009, 69, 4125–4133.
- Nakaya, T.; Ogawa, S.; Manabe, I.; Tanaka, M.; Sanada, M.; Sato, T.; Taketo, M.M.; Nakao, K.; Clevers, H.; Fukayama, M.; et al. KLF5 regulates the integrity and oncogenicity of intestinal stem cells. Cancer Res. 2014, 74, 2882–2891.
- Lin, S.; Lee, S.J.; Shim, H.; Chun, J.; Yun, C.C. The absence of LPA receptor 2 reduces the tumorigenesis by ApcMin mutation in the intestine. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 299, G1128–G1138.
- Guo, L.; He, P.; No, Y.R.; Yun, C.C. Kruppel-like factor 5 incorporates into the beta-catenin/TCF complex in response to LPA in colon cancer cells. Cell Signal 2015, 27, 961–968.
- Montrose, D.C.; Galluzzi, L. Restored Ketosis Drives Anticancer Immunity in Colorectal Cancer. Cancer Res. 2022, 82, 1464–1466.
- Wei, R.; Zhou, Y.; Li, C.; Rychahou, P.; Zhang, S.; Titlow, W.B.; Bauman, G.; Wu, Y.; Liu, J.; Wang, C.; et al. Ketogenesis Attenuates KLF5-Dependent Production of CXCL12 to Overcome the Immunosuppressive Tumor Microenvironment in Colorectal Cancer. Cancer Res. 2022, 82, 1575–1588.
- Pagliuca, A.; Valvo, C.; Fabrizi, E.; di Martino, S.; Biffoni, M.; Runci, D.; Forte, S.; De Maria, R.; Ricci-Vitiani, L. Analysis of the combined action of miR-143 and miR-145 on oncogenic pathways in colorectal cancer cells reveals a coordinate program of gene repression. Oncogene 2013, 32, 4806–4813.
- Shan, T.D.; Tian, Z.B.; Li, Q.; Jiang, Y.P.; Liu, F.G.; Sun, X.G.; Han, Y.; Sun, L.J.; Chen, L. Long intergenic noncoding RNA 00908 promotes proliferation and inhibits apoptosis of colorectal cancer cells by regulating KLF5 expression. J. Cell Physiol. 2021, 236, 889–899.
- Morimoto, Y.; Mizushima, T.; Wu, X.; Okuzaki, D.; Yokoyama, Y.; Inoue, A.; Hata, T.; Hirose, H.; Qian, Y.; Wang, J.; et al. miR-4711-5p regulates cancer stemness and cell cycle progression via KLF5, MDM2 and TFDP1 in colon cancer cells. Br. J. Cancer 2020, 122, 1037–1049.
- Kim, J.K.; Qu, X.; Chen, C.T.; Smith, J.J.; Sanchez-Vega, F.; Garcia-Aguilar, J. Identifying Diagnostic MicroRNAs and Investigating Their Biological Implications in Rectal Cancer. JAMA Netw. Open 2021, 4, e2136913.
- Wang, M.; Gao, W.; Bai, Y.F.; Lu, D.H.; Teng, L.H. Expression and bioinformatics analysis of long-chain non-coding RNA PVT1 in tumors. Zhonghua Bing Li Xue Za Zhi 2017, 46, 485–490.
- Liao, Q.; Chen, L.; Zhang, N.; Xi, Y.; Hu, S.; Ng, D.M.; Ahmed, F.Y.H.; Zhao, G.; Fan, X.; Xie, Y.; et al. Network analysis of KLF5 targets showing the potential oncogenic role of SNHG12 in colorectal cancer. Cancer Cell Int. 2020, 20, 439.
- Nakaya, T.; Aizawa, K.; Taguchi, Y.; Tsuji, K.; Sekine, S.; Murakami, K.; Kasai, M.; Nakano, H.; Kondoh, Y.; Dan, S.; et al. Development of Low-Molecular-Weight Compounds Targeting the Cancer-Associated KLF5 Transcription Factor. ACS Med. Chem. Lett. 2022, 13, 687–694.
- Bialkowska, A.B.; Crisp, M.; Bannister, T.; He, Y.; Chowdhury, S.; Schurer, S.; Chase, P.; Spicer, T.; Madoux, F.; Tian, C.; et al. Identification of small-molecule inhibitors of the colorectal cancer oncogene Kruppel-like factor 5 expression by ultrahigh-throughput screening. Mol. Cancer Ther. 2011, 10, 2043–2051.
- Ruiz de Sabando, A.; Wang, C.; He, Y.; Garcia-Barros, M.; Kim, J.; Shroyer, K.R.; Bannister, T.D.; Yang, V.W.; Bialkowska, A.B. ML264, A Novel Small-Molecule Compound That Potently Inhibits Growth of Colorectal Cancer. Mol. Cancer Ther. 2016, 15, 72–83.
- Shen, X.; Zhang, Y.; Xu, Z.; Gao, H.; Feng, W.; Li, W.; Miao, Y.; Xu, Z.; Zong, Y.; Zhao, J.; et al. KLF5 inhibition overcomes oxaliplatin resistance in patient-derived colorectal cancer organoids by restoring apoptotic response. Cell Death Dis. 2022, 13, 303.
- Kim, J.; Wang, C.; de Sabando, A.R.; Cole, H.L.; Huang, T.J.; Yang, J.; Bannister, T.D.; Yang, V.W.; Bialkowska, A.B. The Novel Small-Molecule SR18662 Efficiently Inhibits the Growth of Colorectal Cancer In Vitro and In Vivo. Mol. Cancer Ther. 2019, 18, 1973–1984.
- Png, C.W.; Weerasooriya, M.; Guo, J.; James, S.J.; Poh, H.M.; Osato, M.; Flavell, R.A.; Dong, C.; Yang, H.; Zhang, Y. DUSP10 regulates intestinal epithelial cell growth and colorectal tumorigenesis. Oncogene 2016, 35, 206–217.
- Kim, J.Y.; Park, S.G.; Kim, K.S.; Choi, Y.; Kim, N.K. CORRIGENDUM: Correction of 4th author’s name: The Kruppel-like factor (KLF5) as a predictive biomarker in preoperative chemoradiation therapy for rectal cancer. Ann. Surg. Treat. Res. 2019, 97, 157.
- Zhao, Y.; Hamza, M.S.; Leong, H.S.; Lim, C.B.; Pan, Y.F.; Cheung, E.; Soo, K.C.; Iyer, N.G. Kruppel-like factor 5 modulates p53-independent apoptosis through Pim1 survival kinase in cancer cells. Oncogene 2008, 27, 1–8.
- Takagi, Y.; Sakai, N.; Yoshitomi, H.; Furukawa, K.; Takayashiki, T.; Kuboki, S.; Takano, S.; Suzuki, D.; Kagawa, S.; Mishima, T.; et al. High expression of Kruppel-like factor 5 is associated with poor prognosis in patients with colorectal cancer. Cancer Sci. 2020, 111, 2078–2092.