Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Atul Pandey and Version 3 by Catherine Yang.
Sleep is essential for the survival of most living beings. Numerous researchers have identified a series of genes that are thought to regulate “sleep-state” or the “deprived state”. As sleep has a significant effect on physiology, lack of total sleep, or particularly rapid eye movement (REM) sleep, for a prolonged period would have a profound impact on various body tissues. REM sleep deprivation affected a total of 652 genes in the brain and 426 genes in the liver. Only 23 genes were affected commonly, 10 oppositely, and 13 similarly across brain and liver tissue. Nine-day REM sleep deprivation differentially affects genes and processes in the brain and liver of rats.
- rapid eye movement sleep deprivation
- differential gene expression in brain and liver tissue
1. Introduction
Sleep is a universal phenomenon but still lack fundamental knowledge of its overall functions and purpose. However, most comparative sleep data exist for terrestrial vertebrates, with much less known about sleep in invertebrates [1]. Though, recently the scientific community has sought to characteristic sleep in non-mammalian species like the fruit fly (Drosophila melanogaster) [2][3][4][2,3,4], the zebrafish (Danio rerio) [5][6][7][5,6,7], the nematode (Caenorhabditis elegans) [8], and bees (Apis mellifera, and Bombus terrestris) [9][10][11][12][9,10,11,12]. Prolonged sleep deprivation is fatal in many of the animals studied, except for pigeons, and several studies have sought to address how sleep promotes survival in rodents and primates [13][14][15][16][13,14,15,16]. Despite the lack of general knowledge regarding the functions of sleep, loss of sleep has been shown to drastically alter the physiology of many of the animals studied thus far [17][18][19][17,18,19]. The degree of physiological changes and the fatality that sleep loss brings about often vary depending upon the nature and duration of sleep deprivation [20][21][20,21]. Many theories have been proposed to explain the evolutionary significance and functions of sleep, which includes “null” and “synaptic plasticity” theories [22][23][22,23]. Recent advancements in sleep research has shed light on two major functions of sleep-reducing synaptic potentiation and waste clearance mediated by glymphatic system [24][25][26][24,25,26]. Thus, sleep seems to have specific, overarching functions for all species that depend on it [19]. While a single characterization cannot be ascribed to sleep, numerous studies link its loss to detrimental effects on metabolism, behavior, immunity, cellular functions, and hormonal regulations across species [27][28][29][30][27,28,29,30]. There are some mechanisms that are associated with behavioral plasticity that are dependent on sociality or physiological state regarding sleep regulation [12][31][12,31]. Additionally, in Drosophila, not all stages of sleep are necessary for basic survival, but questions relating to the critical functions of sleep, plasticity, and its overall importance are still being explored [31].
Rapid eye movement (REM) sleep is an essential part of sleep and is present only in avians and mammals, with the exception of reptiles, in which REM sleep has only been recently discovered [32]. Unearthed thus far, the functional aspects of REM sleep includes mainly memory consolidation, brain maturation, muscle re-aeration, special memory acquisition, and maintenance of general physiological mechanisms of the body [33][34][35][36][37][38][39][33,34,35,36,37,38,39]. In the brain, REM sleep is involved in the reorganization of hippocampal excitability, pruning and maintenance of new synapses during development, and learning and memory consolidation [40][41][42][40,41,42]. Some recent studies also suggest that lack of REM sleep may cause cell death of somatic cells and neurons [43][44][45][43,44,45]. Outside of the brain, deprivation of the REM sleep was found to be associated with acute phase response in the liver, increased synthesis of pro-inflammatory cytokines such as IL1β, IL-6, and IL-12, and an increase in liver enzymes, alanine transaminase and aspartic transaminase [46]. In addition, REM sleep deprivation induces the production of reactive oxygen species (ROS) and causes inflammation [47] and an increase in nitric oxide (NO) in hepatocytes, along with an increase in sensitivity to oxidative stress by the hepatocytes [48]. REM loss also affected the weight and content of nucleic acid in the liver [49]. REM loss was also found to be further associated with oxidative stress and liver circadian clock gene expression [50]. An elevated increase in metabolic rate and UCP1 gene expression is reported in response to chronic REM sleep loss in the brown adipose tissue of rats [51]. Recently, REM sleep loss has been found to be associated with blood-brain barrier function regulation and metabolic changes [52][53][52,53].
2. Differential Gene Expression in Brain and Liver Tissue of Wistar Rats after Rapid Eye Movement Sleep Deprivation
Several of the aforementioned genes that were commonly associated with brain and liver tissue, and any combination of direction of change, were found in previous literature regarding sleep and REM. Several genes of the solute carrier (Slc) family (Table S4) were up- and down-regulated in the brain and liver, respectively, except for slc2a12, which was down-regulated in both the brain and liver. Previously, genes of the slc family were reported to be associated with glucose homeostasis, and slc17a8 is down-regulated in Tinaja cave fish in response to sleep deprivation [54][104]. Slc38a5a is up-regulated in response to sleep deprivation when glucose levels drop and circulating amino acid levels increase [55][105].
REM sleep deprivation is found to be associated with modification of expression of long-term potentiation in the visual cortex of immature rats [56][119], and the research report up-regulation of structural constituents of ribosomes, translation regulation activity, while dopamine receptor-signaling pathway, dopaminergic, cholinergic, GABAergic regulation of synaptic transmission, serotonin binding, and receptor activity were down-regulated in the brain. The dopamine receptor-signaling pathways regulating sleep, learning, and its plasticity are well known [57][58][83,84].
Processes and pathways in the liver following REM sleep deprivation are largely associated with metabolism and the immune system. Many metabolic processes and cellular metabolic processes such as gluconeogenesis, the triglyceride metabolic process, the negative regulation of fatty acid biosynthetic process, oxidation reduction, and the arachidonic acid metabolic process were up-regulated in the liver in response to REM loss. Whole body energy expenditure decreases by 15–35 percent, with the lowest expenditure during slow-wave sleep and a marginally higher expenditure during REM sleep [59][120], and sleep restriction involves reduced muscle glucose uptake, elevated blood glucose production, and pancreatic β-cell dysfunction [60][61][121,122]. An increasing body of evidence indicates that Obstructive Sleep Apnea Syndrome is associated with a variety of metabolic alterations such as dyslipidemia, insulin resistance, and glucose intolerance [62][123]. REM sleep impairs glucose metabolism, which is involved in intermittent hypoxemia [63][124]. An up-regulation of gluconeogenesis may serve as a mechanism to compensate for hypoxemia due to prolonged REM loss. The GO terms related to homeostatic processes, such as cholesterol homeostasis, nitric oxide homeostasis, fatty acid homeostasis, retina homeostasis, and cytosolic calcium ion homeostasis, are associated with genes that were up-regulated in the liver, while T cell homeostasis and other processes associated with the immune system were down-regulated. The immune functions of sleep and associated diseases have been studied [64][65][125,126], and it has been evidenced that the immune system is compromised by lack of sleep [66][127].
REM sleep loss negatively affects several genes linked to neuroactive ligand-receptor interaction pathways in the brain, primarily gamma-Aminobutyric acid, the Human Thrombin receptor, and associated receptor signaling dopamine. A recent review of sleep and protein-dependent synaptic plasticity indicated that sleep deprivation impairs many of the related biological and physiological processes [67][128]. Many of the pathways in the liver that have been up-regulated are linked to metabolism, immunity, and depression. On the other hand, only a few down-regulated pathways in the liver have been established, which include nitrogen metabolism and circadian rhythm. The findings further support the secondary hypothesis that REM sleep loss affects the processes and pathways related to synaptic potentiation and learning and memory (Figure 18A) and processes related to homeostasis and immunity in the liver (Figure 18A–C).
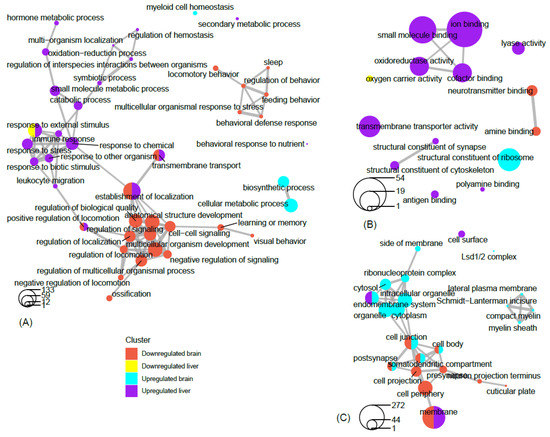 Network analysis of filtered GO terms allowed for the visualization of major themes and the connectivity of processes across brain and liver tissue in rats deprived of REM sleep (Figure 18). Several biological processes, such as positive regulation of locomotion, establishment of localization, and transmembrane transport were terms that were significantly enriched for genes that were both down-regulated in the brain and up-regulated in the liver. Interestingly, responses to external stimuli genes were found to be both positively and negatively affected in the liver, indicating the up- and down-regulation of separate sets of genes associated with this term (Figure 18A). There was no connectivity between terms in the molecular function category; however, terms associated with metabolism and transport, such as oxidoreductase activity, small molecule binding, iron binding, and cofactor binding, were each up-regulated in the liver (Figure 18B). Networking of terms in the cellular component category returned the GO terms cell junction, cell body, post synapse, and somatodendritic compartment, that were up- and down-regulated in the brain and liver, respectively (Figure 18C). To summarize a major theme, some processes that were mainly up-regulated in the liver were also down-regulated in the brain as a result of REM sleep loss. One possible explanation for this is that REM sleep loss influences processes linked to the fear response of the brain and locomotive activity related to the peripheral circadian clock, hemoglobin level, and transport of oxygen throughout the liver. The evidence suggests that the genes and processes involved are highly contrasted between the brain and the liver; however, some processes may be connected across major organs in response to REM sleep loss and should be investigated in the future.
Previous studies have shown that there are transcript level variations in many genes involved in the regulation of reactive oxygen species (ROS), including heme oxygenase, superoxide dismutase, and catalase, in patients with obstructive sleep apnea [68][129]. Similarly, REM sleep has recently been found to be associated with acute phase response and ROS stress in the liver [46][48][46,48]. REM sleep loss also affected several genes such as prostaglandin-endoperoxide synthase (Ptgs2), B-cell lymphoma 2 (Bcl-2), Proto-Oncogene, Tyrosine Kinase receptor (Kit), KRAS Proto-Oncogene (K-Ras), and Fos Proto-Oncogene (Fos), which are marked in cancer pathways. A number of recent studies have shown that sleep dysfunction/loss and cancer processes are closely related [69][70][71][72][73][74][75][130,131,132,133,134,135,136]. However, some emerging evidence also suggests that sleep loss/insomnia prior to the onset of cancer is independently associated with cancer risk [70][74][76][77][131,135,137,138]. Ptgs2, an enzyme, plays a key role in various pathological processes by catalyzing conversion of arachidonic acid to prostaglandins [78][139]. Studies have shown that overexpression of Ptgs2 is associated with angiogenesis, metastases, and immunosuppression [79][80][75,76]. Pgst2 is also found to be associated with the chemoresistance of some malignant tumors, including liver, pancreatic, lung, esophageal, and gastric cancers [81][82][83][77,78,79]. Inhibition of Ptgs2 effectively increased the sensitivity of tumors to drugs [84][140]. Similarly, Bcl-2, Kit, K-Ras, and Fos genes have been found to be associated with cancer [85][86][87][88][141,142,143,144]. These genes play an important role in the sleep-wake cycle regulation and are shown to be correlated with sleep [43][44][89][90][91][43,44,145,146,147]. At the same time, the glycerophospholipid metabolism pathway was found to be significantly up-regulated in the liver (Figure S4). These include the genes Phospholipase, PLa2g, Phosphatidylcholine 2-Acylhydrolase 12A Pla2g12a, Glycerol-3-Phosphate Dehydrogenase 2, Gpd2, CDP-Diacylglycerol Synthase 2, Cds2, and Phospholipid Phosphatase 2, Plpp2. The PLa2g associates with neurodegeneration and elevated mitochondrial lipid peroxidation and dysfunction [92][93][94][148,149,150]. The PLa2g is further found to be positively associated with sleep loss and psoriasis in humans [95][96][151,152]. Similarly, the Gpd2 gene is found to be associated with intellectual disability in humans [97][153] and positively affected due to circadian desynchrony in mice [98][154]. The chronic sleep deprivation in rats affected the protein profile of Gpd2 in hypothalamic astrocytes [99][103]. The functional aspect of the other genes affected (e.g., Pla2g12a, Cds2, and Plpp2) is lacking and needs further exploration. These findings further support the idea of REM sleep being related to restorative functions against diseases and oxidative stress.
Many KEGG pathways were associated with genes that were either significantly up- or down-regulated in the brain or liver as a result of REM sleep loss. The KEGG pathway map demonstrates that many of the genes for ribosomal proteins that are involved in protein synthesis processes were up-regulated in the brain by REM sleep loss. Indeed, research has shown that long-term sleep loss has been found to control several genes in the brain that are linked to the DNA binding/regulation of transcription, immunoglobulin synthesis, and stress response [100][101][55,90]. Contrary to the notion that Homer-1a is a key brain molecule in response to sleep loss in mice, no effect on gene expression of the Homer gene, which suggests that its regulation is modulated during other stages of sleep or is an organism-specific phenomenon [102][81]. The results underscore the complexity of sleep loss and its associated consequences, and sleep phase-, species-, and/or tissue-specific considerations rather than overarching, vague generalizations are required to deeply understand the phenomenon.
Additionally, REM sleep loss negatively affected several genes linked to neuroactive ligand-receptor interaction pathways in the brain, primarily related to gamma-Aminobutyric acid, Human Thrombin receptor, and associated receptor signaling dopamine. A recent review of sleep and protein-dependent synaptic plasticity indicated that sleep deprivation impairs many of the related biological and physiological processes [67][128]. Many of the pathways in the liver that have been up-regulated are linked to metabolism, immunity, and depression. On the other hand, only a few down-regulated pathways in the liver have been established, which include nitrogen metabolism and circadian rhythm.
Findings across studies are inconsistent regarding REM sleep deprivation and locomotor behavior and pain tolerance in rodents. Several studies have shown that REM sleep loss induces locomotor activity [103][104][105][92,155,156], while others have shown decreased locomotor activity [106][157]. The lack of a consistent explanation could be related to procedural changes in the methods of a given study, such as the degree of REM sleep loss. A widely accepted view in the scientific community is that sleep deprivation decreases pain tolerance and increases the transmission of pain in multiple chronic pain conditions [107][108][109][110][111][112][160,161,162,163,164,165]. Nonetheless, selective REM sleep deprivation is correlated with enhanced placebo analgesia effects [113][170]. Similarly, consistency exists between REM sleep loss and its association with the perception of pain [114][171]. Perhaps sleep in general and short-term REM sleep deprivation lower the pain threshold, while long-term sleep deprivation increases the pain threshold. REM sleep deprivation and pain is significantly correlated with environmental conditions (e.g., dry or wet conditions), with pain sensitivity enhanced in dry test conditions but no different in wet conditions. [115][172]. This suggests that further work is needed to understand deeply the relationship between the experience of pain and lack of sleep. Furthermore, a recent microarray analysis shows that Hspa5 gene expression increases not only in the brain but also in the liver as sleep deprivation increases [102][81]. Genes such as Wee1, Slc2a12, Hrk, and Fam110b were commonly down-regulated in both the brain and liver. Currently, however, there are disagreements about the relationship between expression of genes associated with locomotor behavior and pain tolerance, an area that is open for future research.
Network analysis of filtered GO terms allowed for the visualization of major themes and the connectivity of processes across brain and liver tissue in rats deprived of REM sleep (Figure 18). Several biological processes, such as positive regulation of locomotion, establishment of localization, and transmembrane transport were terms that were significantly enriched for genes that were both down-regulated in the brain and up-regulated in the liver. Interestingly, responses to external stimuli genes were found to be both positively and negatively affected in the liver, indicating the up- and down-regulation of separate sets of genes associated with this term (Figure 18A). There was no connectivity between terms in the molecular function category; however, terms associated with metabolism and transport, such as oxidoreductase activity, small molecule binding, iron binding, and cofactor binding, were each up-regulated in the liver (Figure 18B). Networking of terms in the cellular component category returned the GO terms cell junction, cell body, post synapse, and somatodendritic compartment, that were up- and down-regulated in the brain and liver, respectively (Figure 18C). To summarize a major theme, some processes that were mainly up-regulated in the liver were also down-regulated in the brain as a result of REM sleep loss. One possible explanation for this is that REM sleep loss influences processes linked to the fear response of the brain and locomotive activity related to the peripheral circadian clock, hemoglobin level, and transport of oxygen throughout the liver. The evidence suggests that the genes and processes involved are highly contrasted between the brain and the liver; however, some processes may be connected across major organs in response to REM sleep loss and should be investigated in the future.
Previous studies have shown that there are transcript level variations in many genes involved in the regulation of reactive oxygen species (ROS), including heme oxygenase, superoxide dismutase, and catalase, in patients with obstructive sleep apnea [68][129]. Similarly, REM sleep has recently been found to be associated with acute phase response and ROS stress in the liver [46][48][46,48]. REM sleep loss also affected several genes such as prostaglandin-endoperoxide synthase (Ptgs2), B-cell lymphoma 2 (Bcl-2), Proto-Oncogene, Tyrosine Kinase receptor (Kit), KRAS Proto-Oncogene (K-Ras), and Fos Proto-Oncogene (Fos), which are marked in cancer pathways. A number of recent studies have shown that sleep dysfunction/loss and cancer processes are closely related [69][70][71][72][73][74][75][130,131,132,133,134,135,136]. However, some emerging evidence also suggests that sleep loss/insomnia prior to the onset of cancer is independently associated with cancer risk [70][74][76][77][131,135,137,138]. Ptgs2, an enzyme, plays a key role in various pathological processes by catalyzing conversion of arachidonic acid to prostaglandins [78][139]. Studies have shown that overexpression of Ptgs2 is associated with angiogenesis, metastases, and immunosuppression [79][80][75,76]. Pgst2 is also found to be associated with the chemoresistance of some malignant tumors, including liver, pancreatic, lung, esophageal, and gastric cancers [81][82][83][77,78,79]. Inhibition of Ptgs2 effectively increased the sensitivity of tumors to drugs [84][140]. Similarly, Bcl-2, Kit, K-Ras, and Fos genes have been found to be associated with cancer [85][86][87][88][141,142,143,144]. These genes play an important role in the sleep-wake cycle regulation and are shown to be correlated with sleep [43][44][89][90][91][43,44,145,146,147]. At the same time, the glycerophospholipid metabolism pathway was found to be significantly up-regulated in the liver (Figure S4). These include the genes Phospholipase, PLa2g, Phosphatidylcholine 2-Acylhydrolase 12A Pla2g12a, Glycerol-3-Phosphate Dehydrogenase 2, Gpd2, CDP-Diacylglycerol Synthase 2, Cds2, and Phospholipid Phosphatase 2, Plpp2. The PLa2g associates with neurodegeneration and elevated mitochondrial lipid peroxidation and dysfunction [92][93][94][148,149,150]. The PLa2g is further found to be positively associated with sleep loss and psoriasis in humans [95][96][151,152]. Similarly, the Gpd2 gene is found to be associated with intellectual disability in humans [97][153] and positively affected due to circadian desynchrony in mice [98][154]. The chronic sleep deprivation in rats affected the protein profile of Gpd2 in hypothalamic astrocytes [99][103]. The functional aspect of the other genes affected (e.g., Pla2g12a, Cds2, and Plpp2) is lacking and needs further exploration. These findings further support the idea of REM sleep being related to restorative functions against diseases and oxidative stress.
Many KEGG pathways were associated with genes that were either significantly up- or down-regulated in the brain or liver as a result of REM sleep loss. The KEGG pathway map demonstrates that many of the genes for ribosomal proteins that are involved in protein synthesis processes were up-regulated in the brain by REM sleep loss. Indeed, research has shown that long-term sleep loss has been found to control several genes in the brain that are linked to the DNA binding/regulation of transcription, immunoglobulin synthesis, and stress response [100][101][55,90]. Contrary to the notion that Homer-1a is a key brain molecule in response to sleep loss in mice, no effect on gene expression of the Homer gene, which suggests that its regulation is modulated during other stages of sleep or is an organism-specific phenomenon [102][81]. The results underscore the complexity of sleep loss and its associated consequences, and sleep phase-, species-, and/or tissue-specific considerations rather than overarching, vague generalizations are required to deeply understand the phenomenon.
Additionally, REM sleep loss negatively affected several genes linked to neuroactive ligand-receptor interaction pathways in the brain, primarily related to gamma-Aminobutyric acid, Human Thrombin receptor, and associated receptor signaling dopamine. A recent review of sleep and protein-dependent synaptic plasticity indicated that sleep deprivation impairs many of the related biological and physiological processes [67][128]. Many of the pathways in the liver that have been up-regulated are linked to metabolism, immunity, and depression. On the other hand, only a few down-regulated pathways in the liver have been established, which include nitrogen metabolism and circadian rhythm.
Findings across studies are inconsistent regarding REM sleep deprivation and locomotor behavior and pain tolerance in rodents. Several studies have shown that REM sleep loss induces locomotor activity [103][104][105][92,155,156], while others have shown decreased locomotor activity [106][157]. The lack of a consistent explanation could be related to procedural changes in the methods of a given study, such as the degree of REM sleep loss. A widely accepted view in the scientific community is that sleep deprivation decreases pain tolerance and increases the transmission of pain in multiple chronic pain conditions [107][108][109][110][111][112][160,161,162,163,164,165]. Nonetheless, selective REM sleep deprivation is correlated with enhanced placebo analgesia effects [113][170]. Similarly, consistency exists between REM sleep loss and its association with the perception of pain [114][171]. Perhaps sleep in general and short-term REM sleep deprivation lower the pain threshold, while long-term sleep deprivation increases the pain threshold. REM sleep deprivation and pain is significantly correlated with environmental conditions (e.g., dry or wet conditions), with pain sensitivity enhanced in dry test conditions but no different in wet conditions. [115][172]. This suggests that further work is needed to understand deeply the relationship between the experience of pain and lack of sleep. Furthermore, a recent microarray analysis shows that Hspa5 gene expression increases not only in the brain but also in the liver as sleep deprivation increases [102][81]. Genes such as Wee1, Slc2a12, Hrk, and Fam110b were commonly down-regulated in both the brain and liver. Currently, however, there are disagreements about the relationship between expression of genes associated with locomotor behavior and pain tolerance, an area that is open for future research.
Figure 18. Network view of GO term association. Network plots of the top filtered GO terms, depicting the degree of connectivity within and between terms of enriched genes that are up-regulated in the brain, up-regulated in the liver, down-regulated in the brain, and down-regulated in the liver. The circles-legend at the bottom of each left-hand corner indicates the number of genes that are enriched for a given term. Connecting lines indicate a significant degree of semantic similarity between terms. Biological process (A), cellular component (B), and molecular function (C). GO terms were filtered (level = 3) to reduce redundancy and capture major categorical themes prior to visualization of connectivity in network plots, which were designed in R using the cluster Profiler package. Plots of filtered GO terms contained the top 20 significant categories, respectively, per subject cluster.