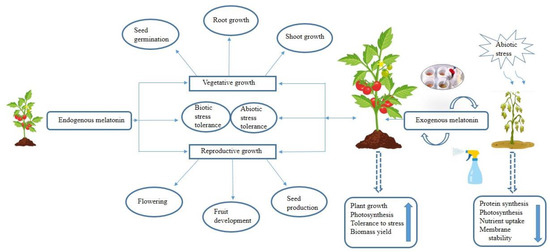
Melatonin has emerged as a crucial plant master-regulator due to its ability to manage oxidative stress from generating free radicals during photosynthesis and respiration. Additionally, melatonin functions as a potent plant biostimulant against biotic and abiotic challenges due to promoting (a) the accumulation of soluble sugars and proline levels, (b) the modulation of the activity of different antioxidant enzymes that scavenge free radicals such as superoxide dismutase (SOD), catalase (CAT), peroxidase (POD), ascorbate peroxidase (APX), glutathione peroxidase (GPX), glutathione reductase (GR), dehydroascorbate reductase (DHAR), and monodehydroascorbate reductase (MDHAR)
[9][10][56,57], and (c) the upregulation of genes associated to stress tolerance
[11][12][46,58].
Recent research results have suggested that applying exogenous melatonin can improve plant photosynthetic rates. Melatonin has been shown to inhibit chlorophyll degradation, increase leaf photosynthetic rates by enhancing RuBisCO activity, and promote the accumulation of dry matter in plants
[13][59]. According to Ren et al.
[5], exogenous melatonin application on stressed plants resulted in a remarkable enhancement in biomass, leaf area, and photosynthetic efficiency. Many studies have reported that exogenous melatonin further enhances plant tolerance in response to drought stress
[14][15][16][60,61,62]. Despite the role of melatonin in the stress mitigation response which has been reported in many plant species, there is still limited understanding regarding its regulatory mechanism on photosynthesis and its protective effect on the photosynthetic machinery. In this context, researchers have outlined the protective effects of melatonin on the photosynthetic machinery of plants exposed to a range of abiotic stresses. Melatonin’s multifaceted impact on photosynthesis offers valuable insights into its potential as a key regulator in promoting plant resilience and survival under adverse conditions.
2. Drought
Among the abiotic stresses, drought stress poses a major threat to plant growth, survival, and productivity. When faced with this challenge in environmental conditions, plants trigger diverse physiological, biochemical, and molecular responses to aid in their adaptation and survival
[17][63]. One component in this adaptation process is the plant growth regulator, melatonin. Research has shown that melatonin plays a crucial role in the regulation of photosynthetic processes and bolsters the antioxidant defense system in plants when subjected to drought stress
[18][64]. Moreover, melatonin contributes to enhanced stomatal conductance and improved photosynthetic efficiency protecting against chlorophyll degradation during drought
[19][65].
In wheat, melatonin has proven to be a valuable component in mitigating the negative impacts of drought stress. Melatonin treatments led to a reduction in ROS and membrane damage, while simultaneously enhancing antioxidant activity
[20][66]. The effects extend to the improvement in various aspects of leaf structure, including large epidermal cells and undamaged chloroplast granules resulting in enhanced photosynthetic activity
[20][66]. Notably, melatonin’s application downregulates the expression of the gene
pheophorbide A oxygenase (PAO), a key component of chlorophyll degradation under stressful circumstances. Furthermore, melatonin positively influences the expression of key photosynthetic genes (e.g.,
RBCS2), leading to increased overall photosynthetic efficiency and supporting crop growth even in drought-stressed environments
[21][67]. Similar positive outcomes were observed in oats subjected to PEG-induced drought stress when melatonin was applied
[22][68]. The treatment improved performance indices, such as PI
ABS and PI
total, and showed a remarkable impact on quantum yield and efficiency of PSII. Moreover, the application of melatonin led to the upregulation of five genes (
PsbA, PsbB, PsbC, PsbD, and
PsbO) encoding core proteins of PSII, further supporting the enhancement of photosynthesis performance and resilience
[23][69].
Studies focusing on maize seedlings have revealed the impact of melatonin in mitigating the detrimental effects of drought on plant growth. Upon melatonin treatment, several favorable changes were observed. Firstly, stomatal conductance was improved, leading to the better regulation of water loss and helping plants against drought stress
[15][23][61,69]. Secondly, photosynthetic efficiency was enhanced, contributing to the overall growth of the maize seedling. Thirdly, the treatment led to a reduction in electrolyte leakage and the accumulation of H
2O
2. Finally, the beneficial effects of melatonin treatment extend to the accumulation of osmoprotectants, stabilization of chloroplast structure, and supported cell expansion
[15][16][23][61,62,69].
The endogenous concentration of melatonin showed a significant correlation with increased photosynthetic capability and stress-related phytohormones when exposed to water scarcity conditions
[24][70]. Exogenous melatonin treatments further amplified this effect by increasing the levels of indole acetic acid (IAA) and zeatin, while reducing the synthesis of the stress-related molecules H
2O
2 and aminocyclopropane-1-carboxylic acid (ACC)
[24][70]. Furthermore, supplementation with melatonin has been found to counteract drought stress effects by modulating the abscisic acid (ABA) levels. It has been reported that melatonin treatment upregulates genes involved in ABA catabolism, and simultaneously suppresses genes related to ABA biosynthesis, ultimately alleviating stomatal closure under drought conditions
[10][24][57,70]. The dual action resulted in improved photosynthetic capacity and water usage efficiency in wheat
[24][70].
The exogenous application of melatonin in various other crops has shown promising results in enhancing drought resistance. According to Campos et al.
[17][63], the application of lower concentrations in coffee expands the root system and protects the photosynthetic apparatus, improving gas exchange, carboxylation efficiency, and chlorophyll contents. Similarly, the application in
Triticale hexaploide increased stomatal conductance, net photosynthetic rate, transpiration rate, and relative chlorophyll concentration while enhancing the activities of SOD and POD, and reducing ROS and malonaldehyde (MDA) content
[25][71]. In kiwi plants, melatonin promoted electron transport in PSII, prevented stomatal closure, and increased light energy absorption through the induction of the transcription of 11 genes encoding for CO
2 fixation enzymes, mitigating the suppression of biomass accumulation and photosynthesis impairment caused by drought
[18][64]. Melatonin treatment through foliar and root irrigation in soybean resulted in a slight increase in chlorophyll levels and a reduction in ABA content compared with control plants, indicating improved drought resistance
[26][72]. Furthermore, in
Chrysanthemum seedlings, exogenous application lessened the impact of drought stress on the photosynthetic mechanism, promoted seedling growth, reduced water loss, preserved chlorophyll levels, increased soluble protein and sugar levels, and enhanced antioxidant enzyme activity, leading to improved drought tolerance
[27][73]. Finally, foliar treatment on
Moringa oleifera enhanced photosynthetic pigment constituents, and the accumulation of IAA and phenolics, resulting in increased photosynthesis
[28][74].
Ranunculus asiaticus and
Phoebe showed increased accumulation of photosynthetic pigments
[29][30][75,76] following melatonin treatment
[29][30][75,76].
3. Salinity
Salinity stress poses a significant challenge to crop productivity, affecting critical physiological and biochemical processes such as photosynthesis. The adverse effects include stomatal closure and disruptions in photosynthetic parameters. Various studies have demonstrated that melatonin has shown a promising role in alleviating these effects in plants by increasing photosynthetic pigment content and strengthening antioxidant defenses.
Melatonin has shown remarkable effectiveness in mitigating the negative impact of salinity stress on cucumber plants. It effectively reduces the decline in the net photosynthetic rate, the maximum quantum efficiency of PSII, and the overall chlorophyll content during salinity stress. Additionally, melatonin treatment lowers electrolyte leakage and MDA concentration and increases chlorophyll production in leaves under salinity stress
[31][77]. In tomato plants subjected to NaCl stress, exogenous melatonin treatment exhibits a reduction in chlorophyll degradation, regulation of the distribution of photosynthetic electrons to minimize the generation of ROS, and enhancement of the activity of enzymes involved in the ascorbate–glutathione cycle
[32][78]. This treatment improved tomato seedling growth under NaCl by increasing antioxidant enzyme activity, proline content, and glycine betaine levels, while decreasing glycolate oxidase activity, chlorophyll degradation, and ROS levels
[33][79]. Furthermore, Zhou et al.
[34][80] have reported that melatonin pretreatment in tomato saline conditions led to an increase in Fv/Fm, the photochemical quenching coefficient, and the percentage of open PSII centers. Exogenous melatonin supplementation has also been shown to enhance sugar accumulation, chlorophyll production, PS-II protection, and upregulation of genes associated with antioxidant protection under salt-stressed conditions in
Beta vulgaris,
Phaseolus vulgaris, and rice
[35][36][37][81,82,83]. In cotton seedlings exposed to salt stress, melatonin effectively prevents the production of ROS, resulting in increased biomass and chlorophyll content, and controls the photosynthetic properties of the plants. It also promoted control over the photosynthesis properties through stomatal opening and the preservation of the mitochondria and grana lamella structure of cotton chloroplasts during salt stress
[38][84].
In wheat seedlings, exogenous melatonin pre-treatment partially mitigated salt-induced suppression of plant growth, as evidenced by improvements in shoot dry weight, IAA concentration, leaf photosynthesis rate, maximum photochemistry efficiency of PSII, and chlorophyll content
[39][85]. Similarly, the application of melatonin in salt-stressed seeds and seedlings regulated the levels of soluble protein, sugar and Ca
2+, ion compartmentation in roots and leaves, and changes in amino acid contents resulting in maintained high water status and a low level of H
2O
2, ultimately leading to a high photosynthetic rate
[40][86]. In oats, melatonin has been found to increase PSII efficiency under salt stress by reducing stress-related PSII damage and encouraging enzymatic antioxidants to scavenge ROS. Melatonin-treated oats showed higher chlorophyll content and proline accumulation, along with reduced levels of MDA and inhibition of electrolyte leakage
[41][13]. Application of exogenous melatonin also significantly improved sugar beet tolerance to salt stress, as evidenced by the higher levels of net photosynthetic rate, chlorophyll fluorescence, and chlorophyll content compared to untreated salt-stressed plants
[35][81].
4. Temperature
Temperature stress poses a significant threat to plant growth and development. Photosynthesis acts as an important sensor of temperature stress by often being inhibited before other cellular processes are affected. To counteract the adverse impact of this stress, the application of melatonin emerges as a very promising strategy
[42][43][87,88]. Melatonin has demonstrated significant effects on the production of total phenols and flavonoids, contributing to the enhancement of heat stress tolerance in grapes
[44][89]. The modulation of the antioxidant defense system, accumulation of proline, decline in MDA content, and increased concentration of chlorophyll and carotenoids were considered to be the protective effects of melatonin on the photosynthetic machinery of wheat to alleviate heat stress
[45][90]. Furthermore, its application has been found to reduce oxidative damage in wheat by decreasing the levels of thiobarbituric acid reactive substances (TBARS) and H
2O
2 and enhancing photosynthetic efficiency through the activation of antioxidants
[46][91]. Moreover, in rice leaves subjected to high temperatures, melatonin treatment resulted in enhanced antioxidant capacity, leading to an improvement in photosynthetic performance. The treated rice leaves maintained a fresh green appearance, and photosynthesis-related parameters with higher values, indicating better photosynthetic efficiency
[47][92].
In tomato plants under heat stress conditions, the exogenous application of melatonin induces the expression of genes encoding the HSPs
[48][93]. Notably, tomato plants exhibited a deceleration in heat-induced leaf senescence, as evidenced by reduced leaf yellowing, an elevated Fv/Fm ratio, and decreased ROS generation
[49][94]. This effect was attributed to melatonin functioning to downregulate the expression of key genes involved in ROS production (e.g.,
RBOHS), genes associated with chlorophyll degradation and senescence. Furthermore, exogenous melatonin treatment in tomatoes leads to an increase in the endogenous levels of melatonin and gibberellins while simultaneously decreasing the levels of ABA
[49][94]. In a study focusing on tomato photosynthesis under heat stress, researchers investigated the mechanism through which melatonin treatment improved photosynthesis when facing ROS production. The findings revealed that the melatonin treatment improved the net photosynthetic rate and chlorophyll fluorescence. Additionally, melatonin played a protective role in PSII by promoting a balanced electron transfer on the donor, reactive center, and acceptor sides, thus mitigating oxidative stress and damage to the photosynthetic machinery
[50][95].
Cold or chilling stress also affects the photosynthetic machinery inducing photoinhibition at both PSII and PSI. However, melatonin treatment has shown promising results in mitigating these adverse effects in melons, rice, tomatoes, watermelons, and peppers.
In melons, melatonin-treated plants exhibited numerous positive responses to cold stress. The treatment led to an increased concentration of antioxidant enzymes, higher chlorophyll content, enhanced photosynthetic rate, improved stomatal conductance, and a higher maximal quantum yield of PSII. Moreover, the fluorescence parameters, especially the Fv/Fm ratio, significantly improved, indicating a reduction in photoinhibition. These findings also suggested that melatonin maintained chlorophyll stability in plants under cold stress, improving light-capturing efficiency and photosynthetic performance through the activation of RuBisCO
[51][96]. Similarly, in rice seedlings subjected to cold stress, pretreatment with melatonin resulted in the improvement in photosynthesis parameters: higher net photosynthetic rate, increased stomatal conductance, elevated intercellular CO
2, and enhanced water use efficiency
[52][97]. When melatonin was applied to tomato seedlings, it yielded significant improvements in growth traits: pigment content, gas exchange elements, and chlorophyll fluorescence metrics
[53][98]. Moreover, exogenous melatonin application also influences the expression of genes involved in phytohormone production in watermelons cultivated under cold-stress conditions. Key genes for jasmonic acid biosynthesis (Cl
AOC1), and IAA biosynthesis (Cl
AMI1) were upregulated in leaves due to melatonin treatment
[54][99]. Finally, in
Capsicum annum plants exposed to cold stress, melatonin treatment improved the photochemical activity of both PSII and PSI as well as the performance of essential photosynthetic enzymes, such as RuBisCO and fructose 1,6-biphosphatase. Furthermore, the exogenous supplementation of melatonin markedly improved photosynthetic pigment molecules, including Chlorophyll
a, Chlorophyll
b, and carotenoids. These improvement were attributed to the upregulation of pigments’ biosynthesis related-genes expression (Ca
CB12, Ca
CAB4, Ca
CAB7, Ca
CAB8, Ca
CAB21, and Ca
CAB37)
[43][88].
In summary, melatonin treatment has been shown to exert positive effects on the photosynthetic performance of various crops under both heat and cold stress conditions.
5. High Light Intensity
Despite the crucial importance of light in the photosynthesis process, the excess energy dissipated damages the photosystem complex, leading to photosynthetic efficiency decline. Exogenous melatonin is effective in alleviating cell membrane damage and inhibiting cell death under high-light conditions. In Arabidopsis thaliana, melatonin spray application enhanced tolerance to high light stress increasing chlorophyll and carotenoid contents, as well as the net photosynthetic rate
[55][100]. This protective effect of melatonin is primarily attributed to reducing the accumulation of ROS, preserving the integrity of membranes and photosynthetic pigments, and minimizing cell damage. To prevent harmful ROS levels, plants activate antioxidant enzymes, detoxification, and repair mechanisms
[56][101].
6. Metal Toxicity
The productivity of crops worldwide faces a severe threat from heavy metal contamination including cadmium (Cd), copper (Cu), zinc (Zn), nickel (Ni), chromium (Cr), and others. Extensive research has focused on understanding the role of melatonin in regulating plant growth under various heavy metal stresses.
In tomato plants exposed to Cd stress, melatonin boosted H
+-ATPase activity, and increased glutathione and phytochelatin concentrations, facilitating Cd sequestration within plant cells. This led to an improvement in the net photosynthetic rate and the maximum quantum efficiency of PSII photochemistry, resulting in ameliorative effects under Cd stress
[57][102]. Similarly, the growth of strawberry seedlings cultivated in Cd-contaminated soil was severely hindered, leading to significantly lower growth rates, biomass output, chlorophyll levels, and antioxidant enzyme activities. However, the administration of melatonin demonstrated a remarkable increase in growth biomass output by improving the activity of antioxidant enzymes, such as APX, CAT, POD, and SOD, while reducing the accumulation of MDA
[58][103].
The exogenous foliar application of melatonin alleviated copper toxicity in tomato plants via the upregulation of several defense genes’ expression (
CAT,
APX,
GR, and
MDHAR). This enhancement in gene expression indicated that melatonin plays a role in boosting antioxidant capacity and detoxifying Cu
2+ by scavenging ROS, thereby alleviating copper toxicity
[40][86]. Furthermore, in wheat plants subjected to nano-ZnO stress, melatonin spray application enhanced photosynthetic carbon assimilation; this was attributed to increasing RuBisCO and ATPase activity, along with higher chlorophyll concentration
[59][104]. Melatonin administration has also shown promise in reducing Ni phytotoxicity in tomato seedlings. It enhanced gas exchange, increased nutrients, and photosynthetic pigment concentrations, as well as elevated the levels of minerals and secondary metabolites levels improving the net photosynthetic rate, transpiration rate, intercellular CO
2 concentration, and stomatal conductance
[60][105].
Melatonin has been found to be an effective component in reducing the harmful effects of Cr on canola and maize plant growth. By regulating photosynthesis, and improving the structural stability and efficiency of PSII and electron transport flow, it protects against photoinhibition due to oxidative damage
[61][106]. Furthermore, the supplementation in maize plants resulted in a higher photosynthetic rate, increases in chlorophyll content, and improvement in antioxidant enzyme synthesis, effectively mitigating Cr toxicity. The research also explored the role of melatonin in regulating the heavy metal binding ability of the cell wall, further contributing to enhanced stress tolerance
[62][107].
7. UV Radiation
The rapid depletion of the ozone layer has resulted in the intensification of ultraviolet (UV) radiation, posing a serious threat to agricultural production. Natural sunlight reaching the Earth’s surface contains ultraviolet-B (UV-B) light, which can impact plant survival and adaptation. The plant’s response to UV-B light is influenced by several factors, such as the wavelength, intensity, and duration, as well as the presence of photosynthetically active radiation detected by phytochrome and cryptochrome photoreceptors.
When melatonin was supplied, it effectively mitigated the inhibitory effects of UV-B radiation on various aspects of plant parameters. Notably, the detrimental impact on photosynthetic and chlorophyll fluorescence, stomatal apertures, chlorophyll levels, and leaf membrane damage were dramatically reduced. Additionally, melatonin exposure in the presence of UV-B stress led to an increase in the level of endogenous melatonin. The treatment is associated with the enhanced expression of the genes-encoding for the antioxidant enzymes APX and CAT which results in a reduction in the accumulation of H
2O
2 in
Arabidopsis leaves exposed to UV-B radiation (
[63][108]. Similar to this, melatonin-treated Arabidopsis subjected to UV-B stress demonstrated increased GR, APX, and SOD activity as well as lower lipid peroxidation levels and higher Fv/Fm values
[64][109]. The melatonin application reduced the negative effects of UV-B stress on the biomass, photosynthetic pigment levels, and membrane lipids of the rosemary in vitro shoots increasing the activity of antioxidant enzymes and accumulation of total phenols
[65][110].
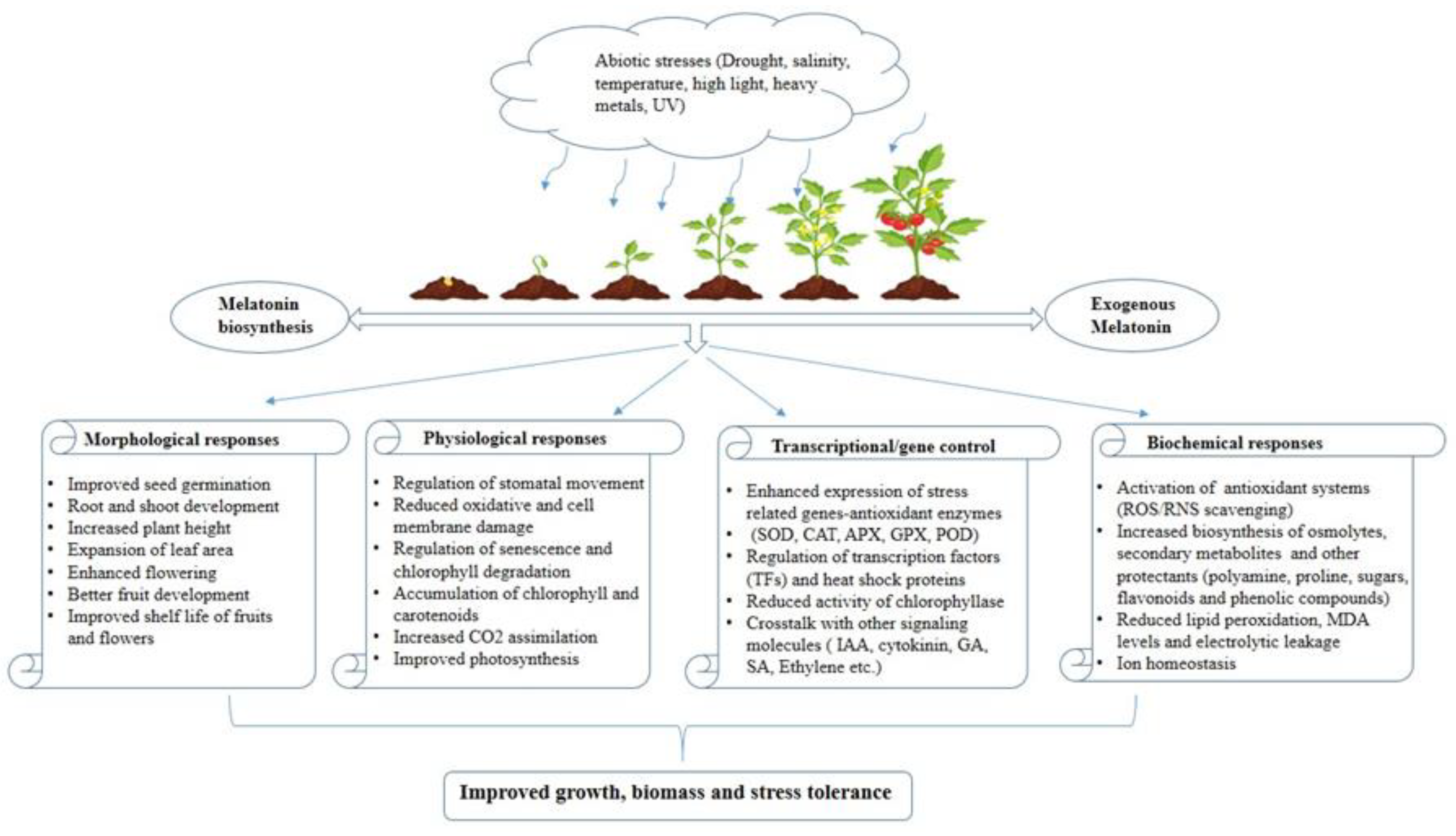
Figure 2 exhibits how plants generally react when exogenous melatonin is applied to them in a stressful environment.
Figure 2.
Response of plants treated with melatonin under abiotic stress conditions.