Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 2 by Catherine Yang and Version 1 by Simon Pierce.
The state of ‘being alive’ is difficult to characterize because ‘life’ is currently defined using superficial features or long-term processes, rather than a single physical property unique to living things. Here, a mechanistic theory and definition of life is outlined, based on the observation that a disparate range of biomolecules share the principle of uniplanar conformation state changes that direct thermodynamic disequilibria into progressive motion to perform work.
- brownian motor
- death
- definition of life
- feynman–smoluchowski ratchet
1. Introduction
Life is a bewilderingly complex phenomenon involving a vast range of integrated biochemical and biophysical processes. Every cell contains millions of components performing very specific roles: biological complexity that is difficult to summarize and distil into a single defining feature. Indeed, life is typically described with a combination of properties (e.g., growth, structure, self-sustaining replication, capacity to evolve, homeostasis and metabolism) to the extent that ‘biologists now accept a laundry list of features characteristic of life rather than a unified account’ [1]. Recent thinking on the attempt to define life could even be described as defeatist [2].
More optimistically, ourthe understanding improves with advancements in science and technology, and in light of current knowledge some of the discussion so far has proven to be relatively superficial. For example, living organisms demonstrate agency or an apparent sense of purpose (end-directed activity, also termed teleonomy), which has been suggested as the defining feature of life [3,4][3][4]. Some proponents have gone so far as to suggest that even the simplest biological organisms possess a literal, cognizant sense of purpose [5]. However, agency cannot be the distinguishing feature of life because it is not unique to biological organisms. Robot vacuum cleaners, clockwork toys and heat-seeking missiles evoke a sense of agency in human observers in precisely the same way that a turtle, a beetle or a bee would, but they do not exhibit any other features of life. This illustrates a key point: current theories and definitions fail because they focus on secondary phenomena or emergent properties without successfully discerning the underlying mechanism producing these effects.
2. Philosophical Barriers to Defining Life
Before setting out to discover what life is, it is important to address certain philosophical arguments that cast doubt on whether the search for an explanation of life is a realistic proposition or even a worthwhile venture [2]. A classic argument against the prospect of a scientific theory of life arises from the fact that all organisms on Earth have a common evolutionary origin. Another contention is that definitions of life have been formulated very differently across a range of scientific disciplines, including different fields of the natural sciences and artificial life (Alife) research [2]. In fields such as astrobiology there may be various definitions for various applications, not all of which attempt to explain life. A working definition may be satisfactory for practical applications such as detecting habitable environments, whereas attempts to understand the origin of life are based on the same kind of reductive biological sciences used to scrutinize the life presently occupying the Earth, and definitions have similar theoretical goals. Definitions for Alife can only be speculative until biology has successfully explained organic life, from which to draw comparisons. This is not to say that only biology matters, rather that a realistic theory of life in organic systems would be a useful starting point for speculative considerations of life. In a sense, biology currently fails in its duty to inform other branches of science, and a lack of a clear definition of the phenomenon at the heart of biology is a major source of embarrassment. Essentially, there is good reason to attempt a theory and definition of life, and no good reason not to. A spectrum of complexity is evident from atoms, simple chemical compounds, complex macromolecules, cells, multicellular microbes through to large-scale organisms, and the point along this spectrum at which chemistry becomes biology (abiogenesis) is difficult to identify and define, lying at the empirical and philosophical heart of the problem [15][6]. However, organisms, as material objects, consist of atoms and molecules and thus exhibit measurable physicochemical properties, and at every point along the spectrum scaling from atoms to organisms the researchers now possess the methods to quantify and compare the states of matter, and have actually done so (Figure 1).
Figure 1. The properties of matter can now be investigated and visualized across a wide range of spatial scales, from macroscopic objects to atoms, promising a mechanistic explanation of living things as material objects. Individual images are credited in descending order by (1). material object (Karl Gaff; ZEISS Microscopy; Electron Microscopy Facility at The National Cancer Institute at Frederick; Guest2625; Y. Roiter, M. Ornatska, A. R. Rammohan, J. Balakrishnan, D. R. Heine, and S. Minko; Kota Iwata; NIST, Joseph Stroscio) and (2). visualization method (Zeiss Microscopy; Zeiss; Oak tree road; AAMonitor96; Rickinasia) and are public domain or published under a Creative Commons Attribution-Share Alike 2.0, 3.0 or 4.0 license at commons.wikimedia.org, accessed on 8 July 2023).
3. The Single Property Defining Living Systems
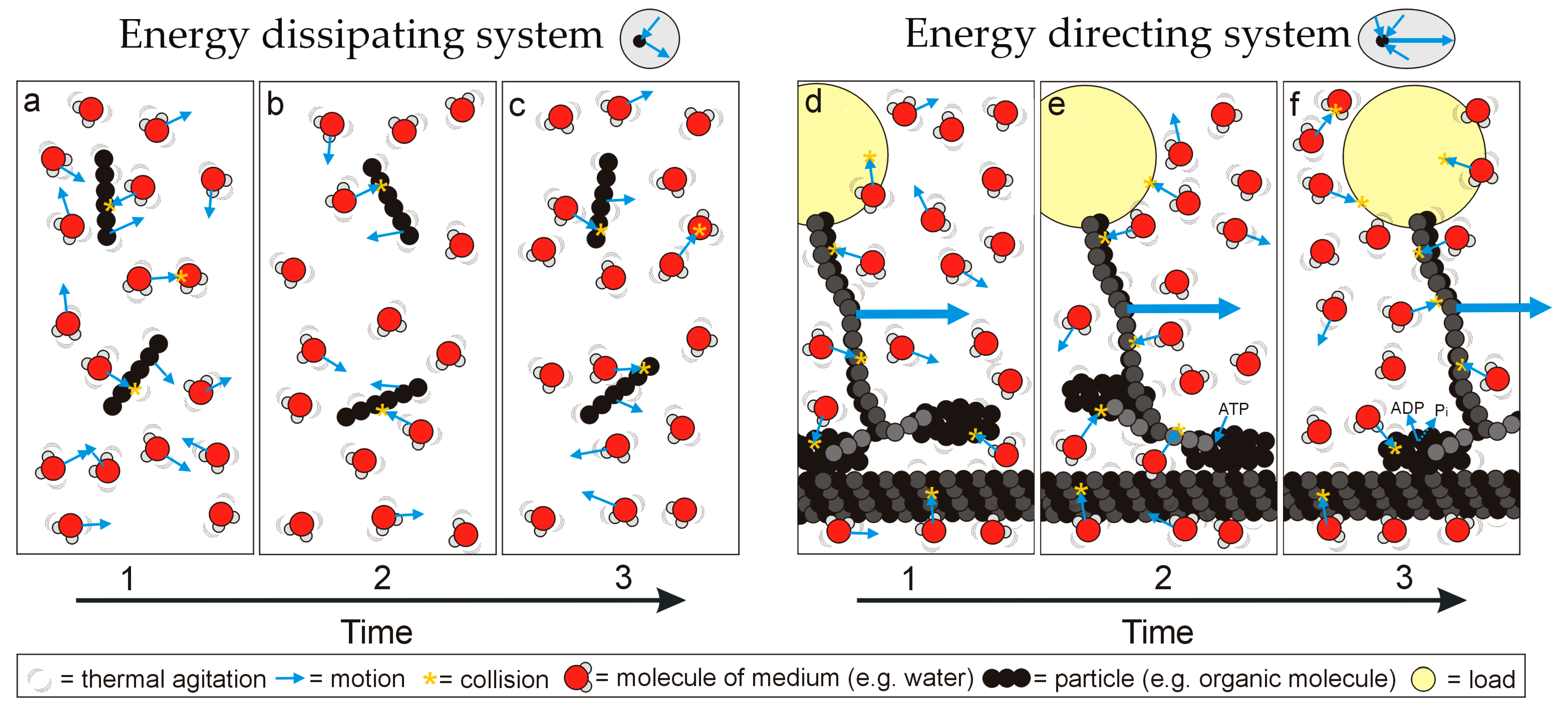
The structurally diverse biological macromolecules discussed above exhibit a shared principle of operation: that of conformation state changes directing thermodynamic disequilibria into unidirectional motion and thus work (local entropy reduction). Alternatively, molecules without preferred configuration state changes move randomly, dissipate energy inputs and are not involved in performing work. This simple functional difference suggests the existence of two fundamental functional classes of matter (‘energy directing’ or ‘energy dissipating’; Figure 2), forming the basis of the difference between living and non-living systems. Life can be defined thus:
Life is a self-regulating process whereby matter undergoes cyclic, uniplanar conformation state changes that convert thermodynamic disequilibria into directed motion, performing work that locally reduces entropy.Figure 2. Simplified representation of how different types of matter respond to the chaotic thermal environment: matter either exhibits (a–c) a banal conformation that moves randomly under thermal agitation and bombardment, dissipating energy, or (d–f) uniplanar conformation changes that convert thermal agitation or excitation into directed motion, as part of an energy directing system that can perform work over time (the representation is inspired by a motor protein ‘walking’ along a microtubule to pull a load). Note that the motive force is Brownian motion (i.e., thermal agitation of the particles themselves but also bombardment by molecules of the surrounding medium; heat engine function must be considered in the context of the medium). For the energy directing system, ATP has an additional role in completing the conformation, affecting the thermodynamic disequilibrium across the particle and preventing backwards motion.Self-regulation via integrated networks [15][6] and autonomy [63][7] are key concepts highlighted in this definition. It is not the single protein (the single heat engine) that should be considered alive, but the integrated, self-regulating and self-replicating network of heat engines. For example, looms use cyclic conformation changes (mechanical action) to convert energy and matter (electricity and wool) into an ordered state (cloth) following a pattern encoded as a set of instructions (programmed information). However, looms are not self-regulating systems and require external input (from a biological organism) for their creation, maintenance, operation and programming.Indeed, self-regulation also encompasses the process of self-replication. Mules, dogs, humans, plants, bacteria, archaea all rely on networks of heat engines performing work and replicating within them. Organisms are ‘alive’ from one moment to the next due to the operation of heat engines.The process encompassed by the definition determines the immediate state of being alive, agrees with the concept of disequilibria driving Feynman–Smoluchowski Brownian ratchets [48[8][9],54], is a mechanism that aggregates matter to produce negative entropy [19][10], underpins the ‘self-sustaining kinetically stable dynamic reaction network derived from the replication reaction’ [15][6], its components are subject to the further long-term processes of mutation and natural selection [7[11][12],8], and it agrees with the ‘plasmogenic’ view of life as the physicochemical reactions occurring in the protoplasm [18][13]: the definition is thus consistent with a range of fundamental biological and physical concepts. Lack of coordinated, directed motion in matter reflects a state of non-life, and where directed motion was previously evident in a molecular network, this lack essentially determines death. ‘Animate matter’ really is an appropriate lay description for the essential process underpinning life, albeit one that does not quite capture the range of scales (nanoscopic to macroscopic) and the intricacy of the processes involved.
References
- Mariscal, C.; Doolittle, W.F. Life and life only: A radical alternative to life definitionism. Synthese 2020, 197, 2975–2989.
- Machery, E. Why I stopped worrying about definitions of life … and why you should as well. Synthese 2012, 185, 145–164.
- Monod, J. Le Hasard et la Nécessité: Essai sur la Philosophie Naturelle de la Biologie Modern; du Seuil, C., Ed.; Points Essais: Paris, France, 1970; p. 248. ISBN 9782020028127.
- Talbott, S.L. Evolution and the purposes of life. New Atlantis 2017, 51, 63–91.
- Shapiro, J.A. Bacteria are small but not stupid: Cognition, natural genetic engineering and socio-bacteriology. Stud. Hist. Philos. Biol. Biomed. Sci. 2007, 38, 807–819.
- Pross, A. What is Life? How Chemistry Becomes Biology; Oxford University Press: Oxford, UK, 2016; p. 224. ISBN 978-0198784791.
- Ruiz-Mirazo, K.; Moreno, A. Autonomy in evolution: From minimal to complex life. Synthese 2012, 185, 21–52.
- Moore, A. Brownian ratchets of life: Stochasticity combined with disequilibrium produces order. BioEssays 2019, 41, 1900076.
- Branscomb, E.; Biancalani, T.; Goldenfeld, N.; Russell, M. Escapement mechanisms and the conversion of disequilibria; the engines of creation. Phys. Rep. 2017, 677, 1–60.
- Schrödinger, E. What is Life? Cambridge University Press: Cambridge, UK, 1944; p. 194.
- Darwin, C.; Wallace, A. On the tendency of species to form varieties; and on the perpetuation of varieties and species by natural means of selection. Zool. J. Linn. Soc. 1858, 3, 45–62.
- Darwin, C. The Origin of Species by Means of Natural Selection or the Preservation of Favoured Races in the Struggle for Life; John Murray: London, UK, 1859; p. 502.
- Herrera, A.L. La Plasmogenia. Nueva Ciencia del Origen de la Vida; Cuadernos de Cultura; Luis Morote: Valencia, Spain, 1932; p. 45.
More