Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Jan Smalle and Version 3 by Jessie Wu.
Biological modularity refers to the organization of living systems into separate functional units that interact in different combinations to promote individual well-being and species survival. Modularity provides a framework for generating and selecting variations that can lead to adaptive evolution. While the exact mechanisms underlying the evolution of modularity are still being explored, it is believed that the pressure of conflicting demands on limited resources is a primary selection force. One prominent example of conflicting demands is the trade-off between survival and reproduction.
- biological modules
- survival-reproduction trade-off
- reproduction maximization
- reproduction assurance
- cytokinin
1. The Survival-Reproduction Trade-Off in Plants
Before delving into the role of each hormone module in the trade-off between survival and reproduction, it is important to address certain processes and concepts that differentiate the survival-reproduction trade-off in plants from that of other organisms. Notably, plants possess unique adaptations that enable them to sustain growth and reproduction even in the face of heightened stress conditions. Unlike mobile organisms, plants are unable to escape unfavorable conditions, thus relying on growth and metabolic adjustments to cope with their environment. In contrast to most non-plant species, which respond to stress by entering a quiescent state, plants ensure their survival by modifying growth and, in many cases, sustaining growth at a reduced rate [1][14]. As a result, when discussing plants, it is more accurate to use the terms “reproduction assurance” under elevated stress conditions and “reproduction maximization” under optimal conditions instead of the conventional “survival-reproduction trade-off”. These more precise descriptors better capture the dynamics of plant responses to varying environmental circumstances, highlighting the strategic choices plants make concerning reproduction in different contexts. This reproduction assurance versus reproduction maximization balance will be referred to as the reproduction trade-off throughout the text.
1.1. Direct Reproduction Assurance, Indirect Reproduction Assurance, and Reproduction Maximization—Definitions
We can make a distinction between direct and indirect reproduction assurance strategies (Figure 1). Direct strategies involve metabolic and growth adjustments that enable reproduction under elevated stress conditions, albeit with reduced reproductive output. On the other hand, indirect reproduction assurance strategies encompass growth and physiological changes that delay reproduction until environmental conditions become more favorable. Depending on specific environmental cues, indirect strategies may eventually give way to direct strategies, ensuring successful reproduction under prolonged environmental stress.
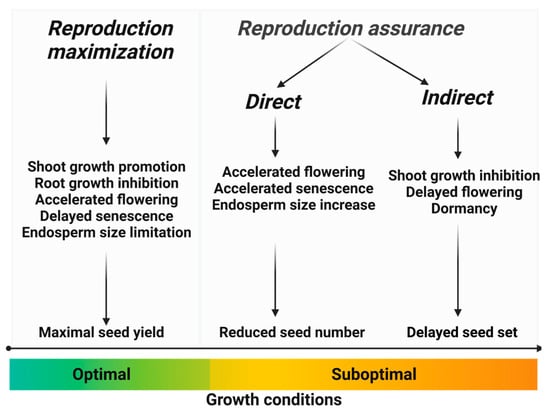
Figure 1. The impact of favorable and unfavorable growth conditions on the balance between reproduction maximization and survival (reproduction assurance), and the processes involved in maintaining this balance.
Hence, it is crucial to perceive the connection between survival and reproduction in plants as a continuum rather than a binary distinction. It represents a spectrum wherein one end emphasizes reproduction maximization, the other end focuses on indirect reproduction assurance, and different forms of direct reproduction assurance occupy the space in between. This understanding implies a diverse range of strategies and resource allocations between survival and reproduction, with varying degrees of emphasis depending on the prevailing environmental conditions.
1.2. Most Prominent Processes Involved in the Reproduction Assurance-Reproduction Maximization Trade-Off
Based on the prevailing environmental conditions, plants employ a diverse array of developmental and physiological strategies to effectively balance reproduction assurance and reproduction maximization. Some key examples are given in the following subsection.
1.2.1. Co-Regulation of Photosynthesis and Reproduction
Considering that photosynthesis is the main supplier of energy and resources for the development of reproductive structures in plants, it is expected that higher rates of photosynthesis would positively impact reproductive success. The connection between photosynthesis, reproduction, and environmental conditions dates back to the early stages of the evolutionary history of plants. For instance, in unicellular photosynthetic algae, elevated photosynthetic rates are positively associated with rates of binary fission (i.e., cell division), while they are negatively correlated with nutrient availability [2][15]. Studies on mutants of Chlamydomonas reinhardtii that are unable to suppress photosynthesis in response to nutrient deprivation reveal an increased proliferation rate under optimal nutrient conditions but a reduced survival rate under phosphate-deprived conditions [3][16].
1.2.2. Regulation of the Shoot-to-Root Growth Ratio
Since the shoot is the main photosynthetic organ and the main organ responsible for both sexual and asexual reproduction, it is expected that an increased shoot-to-root growth ratio translates into increased reproduction potential, as this represents a maximized carbon allocation for reproductive development and sustenance [4][5][17,18]. Therefore, control of the plant’s shoot-to-root growth ratio is an important reproduction trade-off. The regulation of the shoot-to-root growth ratio is an evolutionary response to the greater water and nutrient stresses faced by land plants compared to their algal ancestors [5][18]. In response to these stresses, plants reallocate resources by inhibiting shoot growth, thereby delaying or minimizing reproduction, in favor of promoting root growth. This reallocation of resources ensures improved hydration and nutritional well-being and also reproductive success in the post-stress period. Inhibition of shoot growth is also an unintended consequence of resource allocation towards enhancing tolerance to abiotic and biotic stresses, and this is generally referred to as the growth-defense trade-off [6][7][8][19,20,21].
1.2.3. Timing of Senescence
Senescence is a process primarily driven by nutrient status and it ensures the reallocation of nutrients based on developmental and environmentally imposed needs [9][22]. At the core of senescence are the controlled degradation of the photosynthetic apparatus, the main nutrient-containing part of leaf cells, and the loading of its nutrient constituents into the phloem for transport toward beneficiary organs [10][23]. Therefore, senescence acts as a mechanism that inhibits photosynthesis and shoot growth, prioritizing resource allocation toward reproductive development. This process plays a critical role in direct reproduction assurance by enabling the reproductive development of plants facing elevated stress levels that pose a threat to shoot and reproductive well-being [10][23].
1.2.4. Regulation of Flowering Time
Flowering time is a critical trait that influences the survival and reproductive success of plant species. Flowering, the transition from vegetative to reproductive growth, limits the size of the shoot and limits the resources that can be allocated to maximal seed yield. However, flowering is obviously also needed for reproduction, and actions that delay flowering will negatively impact the length of the reproduction phase. Thus, flowering time can be viewed as an important reproduction trade-off. From the perspective of maximizing reproduction, early flowering will be beneficial only if it is accompanied by extending the length of the reproduction phase. There are two distinct forms of accelerated flowering that warrant examination in this context (Figure 2). The first type entails accelerated flowering coupled with delayed senescence, which extends the reproductive phase and signifies a focus on reproduction maximization. Conversely, the second type encompasses accelerated flowering without senescence inhibition or in combination with senescence acceleration, representing direct strategies for reproduction assurance in response to environmental stresses.
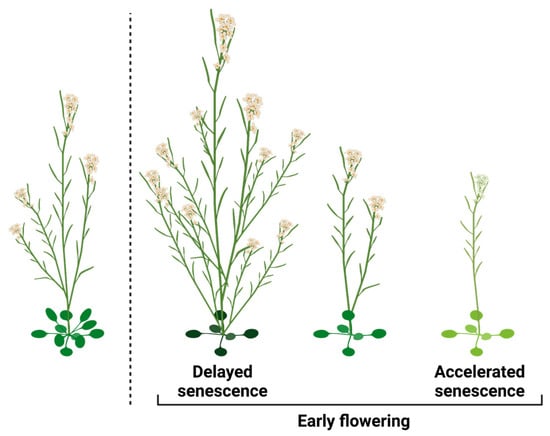
Figure 2. The effects of flowering time on reproduction. Early flowering can enhance reproduction, but its benefits are contingent on delayed senescence. On the other hand, certain forms of early flowering diminish overall reproduction, particularly when accompanied by accelerated senescence.
The regulation of the timing of entry into the reproductive phase in response to stress is an ancient evolutionary trait. For example, in algae, both sexual and asexual reproduction can be triggered by wounding or other forms of stress [11][24].
1.2.5. Dormancy
This aspect of plant developmental programming includes shoot growth arrest and the suppression of seed germination [12][13][14][25,26,27]. Dormancy can be triggered by elevated stress levels, but it also plays a crucial role in the anticipation of impending lethal stress, such as winter, through circadian clock control and photoperiodic cues. Consequently, dormancy can be categorized as a component of indirect reproduction assurance, as it ensures plant survival, thereby preserving the potential for future reproduction.
1.2.6. Determination of Seed Size
Seed size exhibits significant variation among angiosperms and these variations in seed size exemplify another aspect of the reproduction trade-off. Smaller seeds prioritize maximal reproduction potential relative to resource allocation [15][28]. On the other hand, larger seeds serve as a form of indirect reproduction assurance by providing seedlings with ample nutrients during early development [15][28]. This becomes particularly crucial when germination takes place in nutrient-deficient conditions, leading to higher survival rates and an enhanced potential for reproduction [15][28]. The size of the endosperm assumes a crucial role in this context, as it determines the nutrient status of the seed and ultimately affects the viability of developing and germinating seedlings under conditions of nutrient deficiency [16][17][29,30].
2. Origins
Although the exact origins of plant hormones remain unknown, one possible explanation for their emergence is linked to the synthesis of metabolites associated with beneficial physiological and developmental processes. If a particular metabolite consistently arises when a beneficial event like cell division occurs, it creates a stable environment for the evolutionary development of a mechanism capable of binding or detecting this metabolite. Subsequently, this mechanism can then evolve to stimulate further cell division, establishing yet another stable environment. Over time, this could lead to the evolution of a biosynthesis pathway that regulates the production of this metabolite, resulting in a cell division initiation or promotion mechanism that is responsive to the cell’s developmental and metabolic state. The structural similarities between CKs and nucleotides, the building blocks of nucleic acids, raise the possibility that CKs may have evolved from or interfaced with nucleotide-based signaling pathways and that CK is the original module for promoting reproduction [18][109]. Supporting this notion, the promotion of photosynthesis and reproduction by CK is observed even in microalgae, indicating its ancient nature [19][110].
Applying the same logical reasoning, it is worth noting that the primary natural auxin, indoleacetic acid (IAA), bears a structural resemblance to amino acids [20][111]. Interestingly, amino acids universally serve as signals for detecting the nutritional status of cells [21][22][23][112,113,114]. This observation leads to the tempting speculation that the auxin module, an essential well-being module, initially evolved to sense intracellular amino acid levels and respond to low levels by promoting nutrient uptake. Enhancing nutrient uptake in response to nutrient-deficient conditions is one of the primary functions of the current auxin module in higher plants [5][18]. Additionally, IAA, being an ancient molecule, has been demonstrated to promote cellular homeostasis in various primitive photosynthetic organisms [19][24][110,115]. Hence, it is plausible that, similar to CKs, auxins represent the original hormone module for promoting well-being, and the reciprocal interaction between CKs and auxins controlled the original balance between reproduction maximization and reproduction assurance.
Taking this perspective into account, it is essential to recognize that the absence of certain canonical hormone response pathways in ancestral algae does not necessarily negate the functional existence of the corresponding hormone modules in these plant ancestors [19][110]. While ouresearchers' current understanding of hormone response pathways is primarily based on well-studied plant species, it is plausible that ancestral algae possessed alternative mechanisms or pathways that facilitated hormone signaling and modulation of physiological processes related to growth, development, and reproduction. Indeed, it is plausible that hormone signaling in higher plants represents innovations in hormone modules that have emerged in response to environmental changes, including the transition from aquatic to terrestrial habitats during the emergence of land plants. The auxin response pathway is an illustrative example, as it has not been fully identified in algal ancestors [25][116]. However, it is worth noting that auxin does play a role in the growth regulation and physiology of algae. This suggests that while the specific mechanisms and components of the auxin response pathway may have evolved in higher plants, the fundamental influence of auxin on growth and development may have been conserved across plant lineages [25][116].
Interestingly, double-negative signaling, which involves the inhibition of inhibitory pathways, appears to be specific to hormonal modules of land plants. This unique characteristic may reflect the increased environmental stress that land plants experience compared to algae. In land plants, where resource allocation for well-being and reproduction assurance became more crucial, the pressure to optimize resource utilization became accordingly more pronounced. The demand for efficient resource allocation in the face of heightened stress likely drove the evolution of sophisticated mechanisms to ensure precise resource distribution without wastage. Double-negative signaling ensures that resources are allocated strategically and minimally to maintain plant well-being and reproduction, even under challenging environmental conditions.
Indeed, the emergence of new signaling innovations and new hormone modules in land plants can be attributed to their need to adapt to a more challenging environment characterized by drought, nutrient deficiency stresses, temperature fluctuations, increased UV-B exposure, and heightened oxidative stress, which is a combination of both increased number of ROS-generating stressors and the fact that, unlike algae, land plants cannot readily release reactive oxygen species (ROS) or ROS-generating compounds into their surroundings [10][26][27][28][23,117,118,119]. The transition from aquatic to terrestrial habitats brought about significant changes not only in abiotic stress factors but also in the biotic stress environment for land plants. Unlike algae, which primarily face challenges from waterborne pathogens and herbivores, land plants had to contend with a new array of biotic stresses, including diverse microbial pathogens, herbivorous insects, and grazing animals. The transition to land plants brought about another set of environmental challenges, which were indirectly caused by important plant developmental advancements, including the evolution of a vascular system that allowed for vertical plant growth. This advancement, for example, presented new growth obstacles related to shading, which in turn led to the development of a shade-avoidance response. To meet these new requirements, novel hormone modules likely emerged as existing modules were unable to adequately address conflicting demands. This highlights the crucial role of modularity in driving adaptation to changing environmental conditions [29][13].
3. Evolution
The hormone modules found in today’s higher plants are characterized by a high level of complexity, involving the intricate regulation of thousands of genes, as well as numerous post-transcriptional and post-translational controls. Understanding how these modules have maintained their functional focus on reproduction maximization or reproduction assurance, despite their increasing complexity and extensive interactions, may seem challenging. Nevertheless, suppose researchwers view each hormone module as a tool with a central guiding principle. In that case, it becomes clear that the separation of functional focus needs to be maintained to accomplish the delicate balancing act of survival vs. reproduction through antagonistic or synergistic interactions. Under evolutionary pressures, mutations that compromise this focus would prove disadvantageous, whereas those that enhance it would be favored. This principle extends to mutations that improve modular interactions, leading to more refined and precise balancing acts. Hence, the advantageous approach of organizing life around a series of yin/yang balances between opposing motivations likely resulted in the preservation of the functional focus of distinct modules, even as their complexity increased. A case in point is the CK module that promotes photosynthesis and cell division in unicellular algae, representing an ancient version of photosynthetic life [19][110]. The main focal point of this CK module appears to have been preserved all the way to the highly complex multicellular angiosperms, wherein CK still promotes photosynthesis and reproduction.