玉米因其营养特性和在食品工业中的广泛应用而在食品工业中很重要。同时,玉米是世界上产量(吨)最大的粮食作物Maize is important in the food industry because of its nutritional characteristics and wide applications in the food industry. At the same time, maize is the food crop with the largest production volume (tons) in the world [1],因此,它被许多国家视为国家粮食安全的战略作物, and, thus, it is considered to be a strategic crop for national food security by many countries [2].因此,对玉米产品的持续需求推动了其产量的增加。转基因技术可用于改变玉米的遗传性状,常用的性状转化方法很多 Therefore, the continuous demand for maize products has driven an increase in its production. Transgenic technology can be used to change the genetic traits of maize, and many trait transformation methods are commonly used [3][4][5][6].目前,通常研究的性状包括害虫和除草剂抗性。
Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 2 by yun Tong Luo and Version 1 by yun Tong Luo.
转基因玉米是全球最早实现大规模商业化种植的转基因作物之一,在转基因安全政策的制定和实施以及技术监督方面取得优异成绩具有重要意义。玉米的提取技术对核酸检测质量有重要影响。玉米采用的传统核酸检测技术,涵盖变温扩增,等温扩增检测技术和基因芯片技术。Genetically modified (GM) maize is one of the earliest GM crops to have achieved large-scale commercial cultivation globally, and it is of great significance to excel in the development and implementation of safety policy regarding GM, and in its technical oversight.The extraction technology of maize has an important impact on the quality of nucleic acid detection.The traditional nucleic acid detection technology used in maize covers variable-temperature amplification, isothermal amplification detection technology and gene chip technology.
- genetically modified maize
- amplification technology
- nucleic acid detection
1. Introduction
At present, the commonly studied traits include insect pest and herbicide resistance.
In 1996年,美国首次批准, the United States first approved the commercialization of Cry1Ab转基因玉米( GM maize (“Bt176”、, “MON810”和 and “Bt11”)的商业化,转基因玉米现已推广应用27年。), with GM maize now having been promoted and applied for 27 years [7].截至本出版物,已有 As of this publication, 244个转基因玉米品种被批准种植 GM maize varieties have been approved for cultivation [8].根据国际农业生物技术应用服务社( According to a report by the International Service for the Application of Agricultural Biotechnology (ISAAA)的一份报告,截至29年,转基因作物在2019个国家种植。转基因玉米种植面积为6.09×), GM crops are grown in 29 countries as of 2019. The planting area of GM maize was 6.09 × 107嗯 hm2 in 2019年,占转基因作物种植总面积的32%。种植面积是美国最大的,为3.317×, accounting for 32% of the total planting area of GM crops. The planting area was the largest in the United States at 3.317 × 107嗯 hm2,其次是巴西和阿根廷, followed by Brazil and Argentina [9].转基因玉米在全球主要用于动物饲料 The main use of GM maize globally is used for animal feed [10][11]或作为工业原料提取酒精。只有一小部分被人类直接消费。用于食品的转基因玉米主要用于提取玉米油 or as an industrial raw material to extract alcohol. Only a small proportion is consumed directly by humans. The GM maize used for food is mainly used to extract maize oil [12],用于制作玉米糖浆、玉米粉或其他玉米成分, to make maize syrup, maize flour, or other maize ingredients [13]尤其是玉米淀粉,在食品工业中广泛用作增稠剂、胶凝剂、填料、保水剂, especially maize starch, which is widely used as a thickener, gelling agent, filler, and water-retention agent in the food industry [14].用于食品的转基因玉米也直接用作食品生产和加工的原料,例如在墨西哥生产玉米饼 GM maize used for food is also directly used as a raw material for food production and processing, such as in the production of tortillas in Mexico [15];南非的白玉米,它是大多数人的主食 white maize in South Africa, where it is a staple food for most people [16],以及玉米片、爆米花和玉米相关零食, and maize flakes, popcorn, and maize-related snacks [17].毫无疑问,转基因玉米已融入全世界人类的生活,因此,有效规范其生产和加工非常重要。
欧盟( There is no doubt that GM maize is integrated into the lives of humans worldwide and it is, therefore, important to effectively regulate its production and processing.
The European Union (EU) has adopted a traceability management system and a mandatory labeling system for GM products in the market. The EU)对市场上的转基因产品采用了可追溯性管理系统和强制性标签系统。 Regulation 1830年2003月生效的欧盟法规2004/0/EC规定,含量超过9.<>%(质量分数,%m/m)的产品必须标有“遗传改良”或“从转基因作物加工”的字样。此外,应使用DNA拷贝数的比例作为表达方法。以质量百分比为准标准物质的测定结果,拷贝数百分比(DNA拷贝数比,%/ 2003/EC, which came into effect in April 2004, stipulates that products containing more than 0.9% (mass fraction, % m/m) must be labeled with the words “genetic improvement” or “processed from a GM crop”. Moreover, the ratio of the DNA copy number should be used as the expression method. The measurement result of the standard substance, based on the mass percentage, shall prevail, and the measurement result of the copy number percentage (DNA copy number ratio, % cpT/cpE)的测量结果换算成质量百分比) shall be converted into a mass percentage [18].在中国 In China [19]北美洲, North America [20][21],, Australia, New 澳大利亚,Zealand 新西兰[22]印度, India [23]等国家有法律法规明确转基因产品的标签管理体系。这确保了企业经营者和消费者能够获得准确的信息,以便他们能够有效地行使他们的选择自由,并能够控制和验证标签声明。后一项要求使得有必要通过可靠的检测方法检测转基因生物(, and other countries, there are laws and regulations to clarify the labeling management system for GM products. This ensures that business operators and consumers have access to accurate information, so that they can effectively exercise their freedom of choice and be able to control and verify label claims. The latter requirement makes it a necessity to detect the presence of GM organisms (GMO)的存在。转基因作物的检测按目标大致可分为核酸、蛋白质、代谢物的检测。在生物细胞中,与蛋白质相比,DNA相对稳定,即使在作物加工后也不容易被破坏,因此微量或可检测量的DNA片段可能残留在产品中。因此,DNA检测通常是识别作物转基因成分的首选方法。在常见的多核苷酸检测方法中,诸如启动子(例如花椰菜花叶病毒(s) through reliable detection methods. The detection of GM crops can be roughly divided into the detection of nucleic acids, proteins, and metabolites, according to the target. In biological cells, DNA is relatively stable compared to proteins, and is not easily destroyed, even after the crops are processed, such that trace or detectable amounts of DNA fragments may remain in the product. Therefore, DNA detection is often the preferred method for identifying GM components of crops. In common polynucleotide detection methods, targets such as promoters (e.g., Cauliflower mosaic virus (CaMV)) 35S启动子等靶标 promoter [24])、终止子(例如诺帕林合酶终止子(), terminators (e.g., the nopaline synthase terminator (T-nos) [25][26]),或标记基因(), or marker genes (CP4-EPSPS和 and pat [27][28])) are usually invoked as surrogate markers for transformation. However, methods for testing the above-mentioned target genes cannot distinguish between different strains of GM crops with the same exogenous gene transferred, and the specificity is low. Therefore, a pair of primers spanning the junction of the inserted transgene and the flanking 通常作为转换的代理标记调用。然而,上述靶基因的检测方法无法区分转移相同外源基因的不同转基因作物品系,特异性较低。因此,跨越插入的转基因和侧翼DNA连接的一对引物通常用于识别转基因特异性事件DNA is often used to identify transgene-specific events [29][30],即转基因作物的品系,通过检测外源基因与植物基因组的连接区域。目前,该方法广泛用于鉴别转基因玉米品系,如, that is, the strain of the GM crop, by detecting the connecting region of the foreign gene and the plant genome. At present, this method is widely used to identify GM maize lines, such as MON810、, NK603 [31],, Bt11,, TC1507,, GA21 [32],等等。
一般来说,核酸检测需要经过一个核酸提取纯化的过程,靶标检测才能获得结果。提取的核酸的纯度和质量决定了后续扩增的有效性,靶标的设定是检测转基因玉米样品特异性的基础。获取结果的方法也是基于直觉和便利性,根据不同检测场景的需求。例如,简单快速的现场测试方法对于规范转基因作物的进出口至关重要。在这种情况下,收集代表性样本的方式以及分析的时间和可靠性对于顺利实施法规和市场监督至关重要。
, and so on.
Generally, nucleic acid detection needs to go through a process of nucleic acid extraction and purification, and target detection to obtain results. The purity and quality of the extracted nucleic acid determine the effectiveness of the subsequent amplification, and the setting of the target is the basis for the specificity of detecting the GM maize sample. The method of obtaining the result is also based on intuition and convenience, according to the needs of different detection scenarios. For example, simple and rapid on-site testing methods are crucial for regulating the import and export of GM crops. In this context, the manner in which representative samples are collected, and the timing and reliability of the analysis, is critical for the smooth implementation of regulations and market surveillance.
2. Nucleic Acid 核酸提取技术
基于
Extraction Technology
DNA的测定用于检查加工产品的转基因状态,但必须确保-based assays are used to examine the transgenic status of processed products, but the quality of the DNA的质量。在研究不同种类材料的分子生物学检测方法时,DNA提取方法的选择直接影响其性能和效用。目前,植物基因组DNA的常用提取方法是十二烷基硫酸钠(SDS) must be ensured. In the study of molecular biology detection methods of different kinds of materials, the choice of DNA extraction methods directly affects its performance and utility. At present, the common extraction methods of plant genomic DNA are sodium dodecyl sulfate (SDS) [33],, cetyl trimethylammonium 十六烷基三甲基溴化铵 (CTAB)bromide (CTAB) [34],尿素提取法,, urea extraction method, Chelex-100法 method [35]、碱裂解法、聚乙烯吡咯烷酮, alkali lysis method, polyvinyl pyrrolidone-40 ((PVP-40)) [36]、高盐低, high-salt and low-pH萃取法 extraction method [37]和商用套件, and commercial kits [38]等。这些, etc. These DNA提取方法在原则上通常是一致的,包括细胞裂解以释放DNA和去除杂质以进行DNA纯化(图 extraction methods are generally consistent in principle, involving cell lysis for DNA release and the removal of impurities for DNA purification (Figure 1)。然而,不同植物材料中蛋白质、多糖、酚类等物质的含量并不相同,这给). However, the content of proteins, polysaccharides, phenols, and other substances in different plant materials is not the same, which causes great difficulties in the extraction, separation, and purification of DNA的提取、分离和纯化造成了很大的困难。对于植物来源的样品,应根据样品的特性选择合适的方法,并且可能需要对方法进行调整和优化。
. For plant-derived samples, the appropriate method should be selected according to the characteristics of the sample, and the methods may need to be adjusted and optimized.
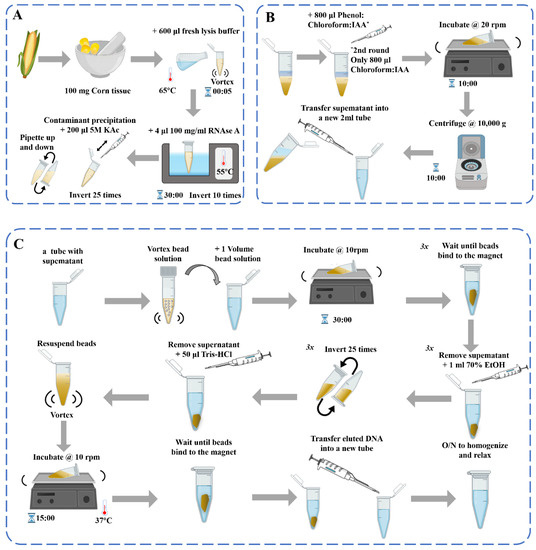
图Figure 1. Schematic diagram of the DNA提取过程示意图 extraction process [39]:(: (A)脱氧核糖核酸提取) DNA extraction;(b)污染物沉淀和苯酚:氯仿纯化 (B) Contaminant precipitation and Phenol: Chloroform purification 2×;( (C)用磁珠纯化基因组) Genomic DNA。 purification with beads.
在玉米样品的
In the process of DNA提取过程中,有针对各种植物组织的玉米原料的相关提取方法,如花粉、叶子、玉米丝 extraction of maize samples, there are related extraction methods of maize raw materials for various plant tissues, such as pollen, leaves, maize silk [40]、胚乳等, endosperm, and others [41].玉米是一种被子植物,其果实具有独特的结构,主要由果皮、种皮、胚乳和胚胎(胚芽、胚根、下胚轴和子叶)组成。有性繁殖玉米的果皮和种皮是由雌亲的子房壁和外皮发育而来的,所有的遗传物质都来自雌性亲本。成熟玉米的果皮和种皮几乎一起生长,很难分离。胚乳由受精极核发育而来,其遗传物质与胚胎和成株不同 Maize is an angiosperm, and its fruit has a unique structure, mainly composed of a pericarp, seed coat, endosperm, and embryo (germ, radicle, hypocotyl, and cotyledon). The pericarp and seed coat of sexually reproduced maize are developed from the ovary wall and the integument of the female parent, and all of the genetic material comes from the female parent. The pericarp and seed coat of mature maize almost grow together and are difficult to separate. The endosperm develops from the fertilized polar nucleus, and its genetic material is different from that of the embryo and adult plant;因此,在提取原料时应适当考虑。传统的基因筛选方法涉及从叶子中提取基因组 therefore, proper consideration should be given when extracting raw materials. The traditional gene-screening method involves extracting genomic DNA from the leaves [42][43]在田间种植大量玉米后单个植物的(或嫩叶)。单籽玉米 (or young leaves) of individual plants after planting large populations of maize in the field. The single-seed maize DNA提取方法主要涉及粉碎种子或利用种子胚乳提取DNA。玉米粗品的DNA提取方法主要用于饲料中转基因生物的检测 extraction method mainly involves crushing the seeds or using the seed endosperm to extract DNA. DNA extraction methods for crude maize products are mainly used in the detection of GMOs in feed [44][45].根据 According to the degree of DNA降解程度由小到大,相关食物可分为生玉米、冷冻玉米、罐装玉米、干包玉米样品 degradation from smallest to greatest, the relevant foods can be divided into raw maize, frozen maize, canned maize, and dry packet maize samples [46].深加工玉米产品主要是调味、膨化、油炸、糖化、发酵面粉产品,如玉米油 The deep-processed corn products are mainly seasoning, puffed, fried, saccharified, and fermented flour products, such as maize oil [47]、玉米淀粉、玉米片、爆米花、玉米棒、脆玉米角、哇头, maize starch, maize chip, popcorn, maize stick, crisp maize horn, wowtou [48]等。
细胞裂解,包括机械、酶和化学裂解,是从细胞组织中提取, etc.
Cell lysis, including mechanical, enzymatic, and chemical lysis, is an essential process for DNA的基本过程。机械开裂,如液氮研磨、热冲击、均质化、超声波处理等 extraction from cell tissues. Mechanical cracking, such as grinding in liquid nitrogen, heat shock, homogenization, and ultrasonic treatment [49],可能导致, may lead to DNA断裂,在实际应用中一般起到辅助作用。需要严格控制超声强度和间隔时间,以避免DNA过度断裂。酶裂解使用特定的酶(如果胶酶)来破坏细胞并释放核酸,这些核酸可以与化学裂解一起提取。
breakage, which generally plays an auxiliary role in practical applications. The ultrasonic intensity and gap time need to be strictly controlled to avoid excessive DNA breakage. Enzymatic cleavage uses specific enzymes (such as pectinases) to destroy cells and release nucleic acids that can be extracted in conjunction with chemical cleavage.
CTAB和 and SDS是两种最常用和最有效的化学裂解剂。通常,表面活性剂(如CTAB和SDS)倾向于与静电力,负轴力和疏水力驱动的聚合物(例如蛋白质和DNA)相互作用 are the two most commonly used and effective chemical lysing agents. In general, surfactants such as CTAB and SDS tend to interact with polymers (e.g., proteins and DNA) driven by electrostatic, diaxial, and hydrophobic forces [50]. CTAB能溶解细胞膜,在高盐(>0.7 can dissolve cell membranes, and in high-salt (>0.7 mol/L NaCl)溶液中,CTAB能与蛋白质和多糖形成可溶性稳定的复合物,但不能析出核酸) solutions, CTAB can form soluble and stable complexes with proteins and polysaccharides, but cannot precipitate nucleic acids [51].在低盐( In low-salt (0.1~0.5 mol/L NaCl)溶液中) solutions [52],, SDS可以在55~65°C的碱性条件下裂解细胞并使DNA游离,同时使蛋白质变性并与其结合 can, under alkaline conditions of 55~65 °C, lyse cells and make DNA free, while denaturing proteins and binding to them [53].最近,离子液体( Recently, ionic liquids (ILs)和磁性离子液体(MIL)已被探索为从复杂生物基质中提取DNA的新型溶剂) and magnetic ionic liquids (MIL) have been explored as novel solvents for extracting DNA from complex biological matrices [54]. IL 和 MIL 通过阳离子和带负电荷的磷酸盐主链之间的静电相互作用以及溶剂和 DNA 碱基的烷基链之间的疏水相互作用促进 DNAs and MILs promote DNA extraction through electrostatic interactions between cationic and negatively charged phosphate backbones, and hydrophobic interactions between the alkyl chains of the solvent and DNA bases 提取[55][56].微尺度电穿孔是一种释放细胞内材料的新兴技术 Microscale electroporation is an emerging technology for the release of intracellular materials [57].其作用机理是,当外加电脉冲的电场强度达到一定数量级时,细胞膜发生构型变化,出现大量微孔。这增加了细胞膜的通透性,有利于各种大分子物质(如 Its mechanism of action is that, when the electric field intensity of the applied electric pulse reaches a certain order of magnitude, the cell membrane undergoes a configurational change, and a large number of micropores appear. This increases the permeability of the cell membrane, which is conducive to the release of various macromolecular substances (such as DNA、RNA、蛋白质、化学小分子等)的释放。然而,调整施加的电压和脉冲长度以确定细胞裂解和DNA提取的最佳条件仍然需要实验验证。
粗, RNA, proteins, chemical small molecules, etc.). However, adjusting the applied voltage and pulse length to determine the optimal conditions for cell lysis and DNA extraction still requires experimental verification.
Crude DNA 提取物含有大量蛋白质、extracts contain large amounts of protein, RNA、糖和其他杂质;因此,DNA纯化是必不可少的。大多数蛋白质在用氯仿或苯酚处理后可以通过变性和沉淀去除,这是蛋白质去除的常用方法, sugars, and other impurities; therefore, DNA purification is essential. Most proteins can be removed by denaturation and precipitation after treatment with chloroform or phenol, which are common methods of protein removal [58].苯酚和氯仿这两种不同的蛋白质变性剂交替使用,可以增强蛋白质去除的效果 The alternating use of phenol and chloroform, two different protein denaturants, can enhance the effect of protein removal [49].应该注意的是,这些化学物质,苯酚和氯仿,具有一定的氧化性,如果使用不当会严重损害 It should be noted that these chemicals, phenol and chloroform, have certain oxidizing properties and can seriously damage DNA。例如,鸟嘌呤对氧化特别敏感,接触苯酚/氯仿会导致形成8-氧鸟嘌呤 if used improperly. For example, guanine is particularly sensitive to oxidation, and exposure to phenol/chloroform can result in the formation of 8-oxoguanine [59].此外,苯酚和氯仿具有挥发性和毒性,根据美国卫生与公众服务部关于致癌物的报告,氯仿被归类为 Furthermore, phenol and chloroform are volatile and toxic, with chloroform classified as “根据实验动物研究中足够的致癌性证据,合理预期是人类致癌物”。reasonably expected to be a human carcinogen based on sufficient evidence of carcinogenicity in laboratory animal studies”, according to a U.S. Department of Health and Human Services report on carcinogens [60].
RNA可在约1°C下用 can be removed by digestion with RNase A消化2~37 h除去,或者通过氯化铯密度梯度离心纯化DNA,从而得到高质量的DNA制备 for 1~2 h at approximately 37 °C, or DNA can be purified by cesium chloride density gradient centrifugation, which results in a high-quality DNA preparation [58].玉米植物细胞具有增厚的次生壁和大液泡,储存大量次生物质,如多糖和多酚。表面活性剂 Maize plant cells have thickened secondary walls and large vacuoles that store a large number of secondary substances, such as polysaccharides and polyphenols. The surfactant CTAB在去除多糖方面优于 is better than SDS at removing polysaccharides [61][62].适当增加 Appropriately increasing the content of CTAB的含量(根据实际情况,如提高到3%,但使用量不低于 (according to the actual situation, such as increasing it to 3%, but not using less than 1% [63]) β-巯基乙醇(mercaptoethanol (0.2%–1%,根据实际情况确定), determined according to the actual situation) [64]能有效去除多糖等次生生物分子。同时,植物中所含的多酚在多酚氧化酶的催化作用下被氧化,导致提取的 can effectively remove polysaccharides and other secondary biomolecules. Simultaneously, the polyphenols contained in plants are oxidized under the catalytic action of polyphenol oxidase, resulting in a lower quality of the extracted DNA质量降低。去除多酚作用的主要方法包括在提取介质中添加抗氧化剂或在研磨过程中添加PVP或抗坏血酸. The main methods to remove the effects of polyphenols include adding antioxidants to the extraction medium or adding PVP or ascorbic acid during grinding [65].通过使用较高浓度的盐和酸性较低的介质可以防止酚类化合物的进一步氧化 Further oxidation of phenolic compounds can be prevented by using a higher concentration of salt and a less-acidic medium [66].已经开发了带有二氧化硅基离心柱的商用固相萃取试剂盒,以标准化程序并使其更有效率。这些试剂盒使用含有 Commercial solid-phase extraction kits with silica-based centrifugal columns have been developed to standardize procedures and make them more efficient. These kits use cleavage buffers containing CTAB或SDS的切割缓冲液,由解离盐组成的结合缓冲液,以促进DNA吸附到二氧化硅吸附剂上,以及含有有机溶剂的洗涤缓冲液来洗脱和纯化 or SDS, binding buffers consisting of dissociative salts to facilitate DNA adsorption onto silica adsorbents, and washing buffers containing organic solvents to eluate and purify the DNA [67].整个提取过程在室温下进行,实验要求严格性低,所得 The entire extraction process is conducted at room temperature, the rigor of the experimental requirements is low, and the concentration and purity of the DNA的浓度和纯度满足大多数当代分子生物学应用的要求。目前,用于植物DNA提取的商业试剂盒也采用功能化磁性材料,通过使用外部磁铁简化纯化步骤。
对于转基因玉米的 obtained meet the requirements of most contemporary molecular biology applications. Currently, commercial kits used for plant DNA extraction also employ functionalized magnetic materials to simplify the purification step through the use of external magnets.
For DNA提取,传统的 extraction of GM maize, the traditional CTAB方法是从高度加工的玉米面筋中提取可扩增DNA的最合适方法,这种面筋通常用作富含蛋白质的饲料成分。该方法可以在实验室测试中产生足够量的扩增DNA,以控制测试底物是否符合授权转基因玉米的事件容许限和标记阈值 method is the most suitable method for extracting amplifiable DNA from highly processed maize gluten, which is often used as a protein-rich feed ingredient. This method can produce sufficient amounts of amplified DNA in laboratory tests to control the compliance of the tested substrate with event tolerance limits and labeling thresholds for authorized GM maize [44].与C TAB方法相比,SDS提取方法通常具有更高的DNA产量,更好的细胞裂解效率,更低的DNA剪切速率和更高的多样性he SDS extraction method often results in a higher DNA yield, better cell lysis efficiency, a lower DNA shear rate, and higher diversity than the CTAB method [68].因此,在目前的众多 Therefore, among the many DNA提取方法中,CTAB和SDS方法的新改进目前正在开发中。 extraction methods at present, new improvements of both the CTAB and SDS methods are currently under development [53][69].
随着分子生物学、精准诊断和治疗领域的快速发展,对具有高通量、纯度和质量的新兴核酸提取技术的需求不断增加。磁珠(
With rapid developments in the field of molecular biology, precision diagnosis, and treatment, the demand for emerging nucleic acid extraction technologies with high throughput, purity, and quality is constantly increasing. As a simple, fast, reliable and automated nucleic acid extraction method, the magnetic beads(MBs)法作为一种简单、快速、可靠、自动化的核酸提取方法) method [70] for nucleic acid extraction has attracted more and more attention. The process of DNA extraction by the MBs method is simple, without repeated centrifugation, column separation, or vacuum filtration [71]. The entire process consists of four steps including lysis, binding, separation, and elution. Therefore, the process is fast and the extraction efficiency is high, culminating in extracted nucleic acids that are high in purity and concentration. Moreover, it is safe and non-toxic, does not use toxic reagents (such as phenol, etc.), reduces personnel hazards, and can be easily adapted for automated batch operation [72]. MBs and automatic nucleic acid extraction instruments can be used for high-throughput automated extraction of nucleic acids from large numbers of clinical samples. MBs with functionalized surfaces that can capture nucleic acids have been widely used to extract nucleic acids from biological samples, and multiple forms have been developed [73]. Examples include manual extraction using magnetic frames or microfluidic chips [74], automated robotic processing, and also the separation of ctDNA by superparamagnetic bead particles in microfluidic platforms for early cancer detection, etc. Combining them with traditional gene extraction methods can efficiently and quickly extract plant nucleic acids, and has an absolute advantage over other methods in the detection of low-content genetically modified components. Sebastian et al. [75] used centrifugal microfluidic technology, using continuous rotating magnetophoresis to facilitate magnetic bead integration and nucleic acid extraction where needed. This solution solves the drawbacks of magnetic bead-based solid-phase extraction that may cause nucleic acid loss due to the handling of magnetic beads when they are transferred from one chamber to another, resulting in higher yield and purity in the end. Jiang et al. [70] and others established a no-elution MB-based nucleic acid extraction method by introducing PEPPG F68 into the lysate and using NaOH solution instead of alcohol as the washing buffer. It avoids the dilution and loss of the target nucleic acid during the elution process, as well as the possible loss of sensitivity and false-negative results. At the same time, the detection sensitivity of loop-mediated isothermal amplification (LAMP) is significantly improved, which has broad application prospects. The current demand by molecular biologists is for convenient, rapid, and inexpensive DNA extraction and detection methods and more compact, portable equipment options to enhance real-time capabilities. The merging of extraction and microflow body technologies [76] to automate nucleic acid detection [77][78] is also in high demand.
3. Traditional Detection Technology
3.1. Variable-Temperature Amplification
Event-specific polymerase chain reaction (PCR) targeting unique sequences spanning the insert DNA and flanking genomic DNA has the highest level of specificity and is commonly used to confirm the identity and authorization status of GMO ingredients and to quantify GMO content [79]. DNA-based PCR methods are considered the most reliable and versatile techniques for the identification and quantification of GMOs, with the chief method being variable-temperature amplification. Variable-temperature amplification technology includes three steps, high-temperature denaturation, low-temperature annealing, and a suitable-temperature extension, and generally refers to PCR technology and its derivatives, such as real-time fluorescent PCR [80][81], droplet digital PCR (ddPCR) [82], nested or semi-nested PCR [83], multiplex PCR [84], etc.
The PCR method is the most commonly used molecular detection technique and is the standard method for detecting GMOs [85]. Standard qualitative PCR is the most commonly used PCR detection method. The mechanism is as follows: after the primer and template DNA are specifically combined in accordance with the principle of complementary base pairing, under the catalysis of Taq DNA polymerase, using deoxyribonucleotide triphosphates (dNTPs) as the raw material, a new DNA strand is synthesized according to the principle of semi-conservative replication. After “n” times of amplification, the total number of progeny DNA is 2n, and finally achieves a million-fold amplification of the number of target DNA fragments, which facilitates the detection of subsequent target DNA fragments [86] and, finally, achieves a million-fold amplification of the number of target DNA fragments, which facilitates the detection of subsequent target DNA fragments. Standard PCR is used for the amplification of transgenic crop genes owing to its simple operation, high efficiency, and low cost. Quantitative PCR (qPCR), based on a standard curve, is considered the gold standard technique for the analysis of GMOs because of its high sensitivity and good stability [87]. Real-time fluorescent qPCR [88] is a technology that adds fluorescent groups to the PCR reaction system and uses the accumulation of fluorescent signals to monitor the entire PCR reaction process in real time. This technology uses the strength of the fluorescent signal to determine the number of specific amplification products over time, and an unknown template is quantitatively analyzed using a standard curve. This technology can quantitatively analyze DNA templates and has the characteristics of high sensitivity, specificity, and reliability; low pollution; and timely and accurate detection. It can perform both absolute and relative quantifications and is widely used to inspect GM maize [89]. Various forms of qPCR are constantly being developed to meet the needs of practical applications, focusing on duplex and multiplex reactions to improve detection throughput and efficiency.
The concept of digital PCR (dPCR) was first proposed by Vogelstein et al. in 1999 [90]. In dPCR, the reaction mixture is divided into many individual reactions called partitions, and each reaction does not contain one or more copies of the target. Reads are partitioned as negative or positive at the endpoints, and DNA concentrations are calculated using a Poisson distribution [91]. The partitioning of reaction volumes using wells on a chip in microfluidics/chip-based dPCR [92] and droplets in emulsion/ddPCR [93] are the two main approaches. In cdPCR, reactions are divided into hundreds or thousands of chambers in a single plate or array. Many studies have used chip-based platforms, such as the microwell chip-based QuantStudio 12k flex dPCR and 3D dPCR (Life Technologies), for the detection of GMOs in the field [94][95]. The Constellation system (Formulatrix) is a plate-based microfluidic dPCR system that offers five-color multiplexing. The biggest difference between the cdPCR platforms is the number of partitions created per sample and the number of samples analyzed in one run [96]. ddPCR has a synergistic effect on droplet microfluidics. It improves the sensitivity of PCR at the single-molecule level by dividing tens of microliters of PCR mixture into tens of thousands of droplets and it can perform absolute quantification without a standard curve, thus avoiding the amplification efficiency bias observed in qPCR. It enables accurate target determination even at low copy numbers and can be significantly cost-effective when combined with multiplexing [97]. However, the droplet reaction generator used for ddPCR is bulky and complicated, which is an important limitation for its use in on-site detection. Thus far, microfluidic platforms for droplet generation using centrifugal forces, such as those utilizing ferrofluids, electromagnets [97], and surface acoustic waves [98]. Using the working principle of the indirect pressurization method, Park et al. [99] developed a pushbutton-activated microfluidic dropenser (droplet dispenser). Its use for sample preparation in ddPCR eliminates the need for benchtop droplet generators and automated pipetting, making ddPCR an on-site molecular diagnostic tool. In conclusion, dPCR has proven to be an effective tool for the quantification of maize and soybean GMOs and GMOs in complex matrix samples with precision and accuracy the same as or better than qPCR methods [100]. Recently developed multiplex dPCR methods [87][101] may be useful for analyzing samples containing multiple genetic modification events.
More types of PCR technologies are constantly being developed to meet the requirements for on-site rapid detection and high throughput. The use of an ultrafast PCR system can significantly reduce PCR run times and the number of reagents required for analysis. Therefore, ultrafast PCR systems have recently been studied and applied in various fields. The latest example of an ultrafast PCR system is a system used in rice detection [102]. The analysis principle is the same as that of real-time fluorescent qPCR, based on SYBR green [103], except that Evagreen dye is used as the intercalating dye instead of SYBR green. It requires 18% of the detection time of traditional PCR and 23% of the detection time of real-time PCR, and it can support small portable analyzers, thus providing a new strategy for the on-site detection of GM maize.
3.2. Isothermal Amplification
Isothermal nucleic acid amplification technology is used for nucleic acid amplification at a constant temperature. According to the different methods of single-stranded template formation, isothermal amplification can be divided into the following four categories: (1) strand-displacing DNA polymerase-mediated reactions such as Loop-Mediated Isothermal Amplification (LAMP) [104], Rolling Circle Amplification (RCA) [105], Cross-Primed Amplification (CPA) [106], Nucleic Acid Sequence-Based Amplification (NASBA [107]), and Multiple Displacement Amplification (MDA) [108]; (2) Enzymatic unwinding primer annealing reaction, such as Helicase-Dependent Amplification (HDA) [109], Recombinase Polymerase Amplification (RPA) [110], and Ligase Chain Reaction (LCR) [111]; (3) RNA transcription-based amplification, such as Transcription Mediated Amplification (TMA); and (4) Requires reactions assisted by single-strand cleavage enzymes, such as Strand Displacement Amplification (SDA) [112], and Isothermal Strand Displacement Amplification (iSDA) [113]. LAMP, RPA, and CPA are widely used for the detection of GM crops.
LAMP employs a DNA polymerase and a set of four specially designed primers that recognize a total of six different sequences in the target DNA. Internal primers containing the sequences of the sense and antisense strands of the target DNA initiate LAMP [104]. The product is a mixture of stem-loop DNA with stems of various sizes and cauliflower-like structures. Multiple loops are induced by annealing between alternating inverted repeats of the target sequence in the same strand. This enables simpler and more selective detection. For example, through a mechanism similar to multivalent antigen antibody interactions, the target sequence has a higher degree of specificity [104]. This method is insensitive to inhibitors, and can be used with crude DNA samples. LAMP is a simple and reliable GM detection method that can be performed on a thermal cycler, heating block, or portable constant temperature real-time amplification system. After the reaction is completed, using nucleic acid staining or fluorescent dyes, such as SYBR® Green and hydroxynaphthol blue, the LAMP products can be visualized and monitored using turbidity analysis or real-time LAMP [114]. Since the first report on the LAMP method in 2000, the number of studies using LAMP to detect GM ingredients has increased annually [115][116][117].
RPA technology was first proposed in 2006 by Piepenburg et al. [118]. In RPA, isothermal amplification of specific DNA fragments is achieved by the binding of reverse oligonucleotide primers to the template DNA and extension via DNA polymerase. It does not require the orientation of the primers to their complementary target sequences. RPA can be used to amplify cDNA generated by the reverse transcription of double-stranded DNA, single-stranded DNA, methylated DNA, RNA, or miRNA, and multiple reverse transcriptases are used for RPA [110]. When using RPA directly in milk [119] or seed powder [120], only thermal lysis, nuclease-free water lysis, or EzWayTM Direct PCR buffer are required to release the desired nucleic acid. With the assistance of a variety of enzymes, the in vitro amplification of nucleic acid can be completed at a constant temperature of 31–37 °C for 20 minutes [110]. Traditional in vitro nucleic acid amplification techniques do not have a rapid response, high sensitivity, high specificity, or low equipment dependence. Compared with SDA, RCA, and LAMP, RPA does not require an initial denaturation step to generate single-stranded (ss)DNA from double-stranded (ds)DNA targets, highlighting its suitability for use in the field [121]. In 2014, a commercial RPA kit launched by the British company, Twist DX [122], made the detection more convenient. Simultaneously, a variety of probes can be combined to expand the application range of RPA technology. RPA appears to be particularly well suited for multiplexing, where different targets can be verified with different efficiencies; however, it currently requires laborious optimization steps. Clustered regularly interspaced short palindromic repeats (CRISPR)/CRISPER-associated protein [123], lateral flow assays [124][125]和微流体 and microfluidics [126]为转基因的现场即时检测提供了额外的选择(图 have provided additional options for the on-site point-of-care detection of transgenes (Figure 2)。
).
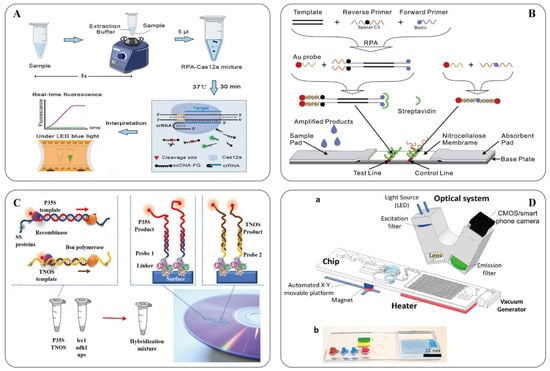
图Figure 2.( (A)) Schematic showing the 用于快速和视觉核酸检测的ORCD检测系统示意图ORCD assay system for rapid and visual nucleic acid detection [123]; (B)) Schematic of the LFNAA(侧流核酸测定)示意图 (lateral flow nucleic acid assay) [125]; (C)) Scheme of the assay for GMOs detection based on 基于多重 RPA 扩增的转基因生物检测方法方案(左)和阵列格式的杂交测定(右)multiplex RPA amplification (left) and hybridization assay in the array format (right) [127]; (D)) Microfluidic platform for sample-to-answer 用于样本到答案数字digital RT-RPA 的微流控平台[126]:: (a) Schematic illustration of the total system consisting of a magnetic platform, heater unit, vacuum generator, detection part with camera, (a) 由磁性平台、加热器单元、真空发生器、带摄像头的检测部件和发光二极管 (LED) 组成的整个系统的示意图。一次性芯片有三个操作区域,用于制备和混合试剂以及检测病原体。(b) 用于芯片可视化的染料装载室。棕色、蓝色和红色分别表示用于样品制备的裂解室、洗涤室和洗脱室。绿色和黄色腔室分别显示RPA混合物和矿物油室。and light-emitting diode (LED). The disposable chip has three operational zones to prepare and mix the reagents and detect the pathogen agent. (b) Dye-loaded chambers for visualization of chips. Brown, blue, and red show lysis, washing and elution chambers, respectively, for sample preparation. Green and yellow chambers show RPA mixture and mineral oil chambers, respectively.
CPA技术是由美塔生物科技有限公司开发的等温 technology is an isothermal DNA扩增系统 amplification system developed by Us-Tar Biotechnology Co., Ltd [106].系统仅依靠一个环结构进行复制 The system relies on only one ring structure for replication [128]. The CPA测定能够在恒定温度下扩增核酸序列,并且只需要一种具有链置换活性的酶和一组五个引物即可进行CPA反应,而无需初始变性步骤或添加切口酶 assay enables the amplification of nucleic acid sequences at a constant temperature and requires only an enzyme with strand displacement activity and a set of five primers to perform the CPA reaction, without the need for an initial denaturation step or the addition of a nickase [129].在 At an assay temperature of 63 °C的测定温度下,DNA模板中瞬时自发变性气泡下引物模板杂交体的形成比相对于模板DNA的高浓度引物对模板链的重新退火更有利。通过退火与模板链不互补的5'末端的交叉引物以及交叉引物上游置换引物的结合来促进链置换。由此产生的靶DNA指数扩增具有高度特异性和灵敏度, the formation of primer template hybrids under transient spontaneous denaturing bubbles in the DNA template are more favorable than the re-annealing of the template strands by high concentrations of primers relative to the template DNA. Strand displacement is facilitated by annealing cross-primers with 5’ ends that are not complementary to the template strand and the binding of displacement primers upstream of the cross-primers. The resulting exponential amplification of the target DNA is highly specific and sensitive [106].传统上, CPA通过昂贵的荧光技术,繁琐的凝胶电泳程序或使用分光光度计测量浊度来产生结果。这些方法需要复杂而笨重的光学元件或暴露于致癌染料,这限制了它们在资源有限的实验室中的更广泛应用 has traditionally yielded results through expensive fluorescence-based techniques, tedious gel electrophoresis procedures, or the measurement of turbidity using a spectrophotometer. These methods require complex and bulky optics or exposure to carcinogenic dyes, which limits their wider use in resource-limited laboratories [128].此外,比色指示剂,如 In addition, colorimetric indicators such as pH敏感染料(中性红)、孔雀石绿(MG)和羟基萘酚蓝(HNB)-sensitive dyes (neutral red), malachite green (MG), and hydroxynaphthol blue (HNB) [130]也被用作监测 were also used as complementary techniques to monitor CPA反应的补充技术。
responses.
3.3. 基因芯片技术
基因芯片技术是美国公司
Gene Chip Technology
Gene chip technology is a GM food detection technology developed by the American company Affymetrix在 in the 1990年代开发的一种转基因食品检测技术。近年来,它作为一种高科技分子生物学工具迅速发展。基因芯片也称为DNA芯片或微阵列s. It has rapidly developed as a high-tech molecular biology tool in recent years. Gene chips are also known as DNA chips or microarrays [131].该技术涉及在固相载体上以有序密度排列大量 This technology involves arranging a large number of DNA片段或寡核苷酸片段,使用固相载体表面的特定探针与标记样品杂交,以及使用芯片扫描仪检测和分析杂交信号 fragments or oligonucleotide fragments at an orderly density on a solid-phase carrier, using specific probes on the surface of the solid-phase carrier to hybridize with labeled samples, and using a chip scanner to detect and analyze hybridization signals [132].它可以定性和定量地准确检测样品中不同类型的 It can accurately detect different types of DNA序列。当微阵列杂交用于基因分析时,可以从很少的实验样品中获得有关基因表达差异的信息。当使用该技术进行检测时,对样品进行预处理和纯化以获得高纯度的DNA样品,然后通过PCR扩增,用荧光标记,并与基因芯片上的DNA探针杂交。然后读取信号以获得结果 sequences in samples qualitatively and quantitatively. When microarray hybridization is used for gene analysis, information on differences in gene expression can be obtained from very few experimental samples. When using this technology for detection, the sample is pretreated and purified to obtain a highly pure DNA sample, which is then amplified by PCR, labeled with fluorescence, and hybridized with the DNA probe on the gene chip. The signal is then read to obtain the result [133].基因芯片可以固定大量针对不同靶基因的寡核苷酸探针 Gene chips can immobilize a large number of oligonucleotide probes for different target genes;因此,基因芯片技术可以同时检测数十甚至数百个基因 therefore, gene chip technology can detect dozens or even hundreds of genes simultaneously [134]并且可以检测转基因食品中的多种成分。卢等 and can detect multiple components in GM foods. Lu et al. [135]采用基因芯片检测方法结合多重 used a gene chip detection method combined with multiplex PCR,在单个芯片上同时检测多对基因。检测了七种转基因玉米成分:Bt176、Bt11、GA21、 to simultaneously detect multiple pairs of genes on a single chip. Seven types of GM maize components were detected: Bt176, Bt11, GA21, Mon810、, Mon863、, TC1507和NK603。该方法大大提高了检测的准确性和效率,灵敏度高达0.01%。图尔克等, and NK603. This method greatly improves the accuracy and efficiency of detection, with a sensitivity as high as 0.01%. Turkec et al. [131]测试了 tested 1830种不同的探针,并为12个转基因品种(1个玉米和<>个大豆品种)开发了高密度寡核苷酸微阵列平台。该方法消除了PCR扩增步骤的需要,简化了分析并允许对每个检测到的GMO进行定量,从而能够以<>%(DNA浓度)的灵敏度对每种转基因作物进行特异性检测。
基因芯片技术具有高并行性、高通量、高特异性、高灵敏度和自动化等优势。但由于该技术开发起步较晚,综合性强,专业性强,成本高,制造基因芯片的过程相对复杂,存在一定的背景干扰等问题。因此,该技术在实际应用中的普及和推广受到限制 different probes and developed a high-density oligonucleotide microarray platform for 12 GM varieties (nine maize and three soybean varieties). This method eliminates the need for a PCR amplification step, which simplifies the analysis and allows the quantification of each detected GMO, enabling the specific detection of each GM crop with a sensitivity of 1% (DNA concentration).
Gene chip technology offers advantages, such as a high level of parallelism, high throughput, high specificity, high sensitivity, and automation. However, due to the late start of the development of this technology, strong comprehensiveness, strong professionalism, and high cost, the process of making gene chips is relatively complicated, and there are certain problems, such as background interference. Therefore, the popularization and promotion of this technology in practical applications are limited [136].目前,全球种植的转基因食用农产品种类有限,转基因成分的检测往往只需要检测几十个靶基因,而基因芯片技术可以进行多基因甚至全基因检测。 At present, the types of GM edible agricultural products grown worldwide is limited, and the detection of GM ingredients often only requires the detection of dozens of target genes, whereas gene chip technology can perform multigene or even whole-gene detection [134].因此,考虑到实验成本和利用率,该技术目前不适合转基因检测。一般来说,基因芯片技术需要极小的样本,具有快速、省时、无污染、准确、适合自动化操作等优点 Therefore, considering the experimental cost and utilization rate, this technology is currently unsuitable for transgene detection. In general, gene chip technology requires extremely small samples and has the advantages of being fast, time-saving, pollution-free, accurate, and suitable for automated operation;然而,目前使用该技术的研究是在疾病诊断和微生物检测领域,转基因检测的研究更集中在微流控芯片上。 however, current research using this technology is in the field of disease diagnosis and microbial detection, and research on transgenic detection is focused more on microfluidic chips [137].微流控芯片通过微处理技术将多个操作平台集成到一个芯片中,样品和试剂消耗少,反应速度快,并行工艺大量。因此,它具有更大的发展潜力。
Microfluidic chips integrate multiple operating platforms into one chip through micro-processing technology, with less consumption of samples and reagents, a fast reaction speed, and a large number of parallel processes. Therefore, it has greater development potential.
References
- FAOSTAT. Available online: https://www.fao.org/faostat/es/#data/QCL (accessed on 21 May 2023).
- Spatial-Temporal Evolution of Scientific Production about Genetically Modified Maize. Agriculture 2021, 11, 246.
- EFSA Panel on Genetically Modified Organisms (GMO); Naegeli, H.; Birch, A.N.; Casacuberta, J.; De Schrijver, A.; Gralak, M.A.; Guerche, P.; Jones, H.; Manachini, B.; Messéan, A.; et al. Scientific Opinion on an Application by DOW AgroSciences LLC (EFSA-GMO-NL-2010-89) for Placing on the Market the Genetically Modified Herbicidetolerant Maize DAS-40278-9 for Food and Feed Uses, Import and Processing under Regulation (EC) No 1829/2003. EFSA J. 2016, 14, e04633.
- Sutradhar, M.; Mandal, N. Reasons and Riddance of Agrobacteriumtumefaciens Overgrowth in Plant Transformation. Transgenic Res. 2023, 32, 33–52.
- Dong, O.X.; Ronald, P.C. Targeted DNA Insertion in Plants. Proc. Natl. Acad. Sci. USA 2021, 118, e2004834117.
- Muppala, S.; Gudlavalleti, P.K.; Malireddy, K.R.; Puligundla, S.K.; Dasari, P. Development of Stable Transgenic Maize Plants Tolerant for Drought by Manipulating ABA Signaling through Agrobacterium-Mediated Transformation. J. Genet. Eng. Biotechnol. 2021, 19, 96.
- Snow, A.A.; Palma, P.M. Commercialization of Transgenic Plants: Potential Ecological Risks. BioScience 1997, 47, 86–96.
- Maize (Zea mays L.) GM Events|GM Approval Database—ISAAA.Org. Available online: https://www.isaaa.org/gmapprovaldatabase/crop/default.asp?CropID=6&Crop=Maize (accessed on 13 July 2023).
- Global Status of Commercialized Biotech/GM Crops: 2019—ISAAA Brief 55-2019|ISAAA.Org. Available online: https://www.isaaa.org/resources/publications/briefs/55/default.asp (accessed on 13 July 2023).
- Anderson, J.A.; Herman, R.A.; Carlson, A.; Mathesius, C.; Maxwell, C.; Mirsky, H.; Roper, J.; Smith, B.; Walker, C.; Wu, J. Hypothesis-Based Food, Feed, and Environmental Safety Assessment of GM Crops: A Case Study Using Maize Event DP-202216-6. GM Crops Food-Biotechnol. Agric. Food Chain 2021, 12, 282–291.
- Avsar, B.; Sadeghi, S.; Turkec, A.; Lucas, S.J. Identification and Quantitation of Genetically Modified (GM) Ingredients in Maize, Rice, Soybean and Wheat-Containing Retail Foods and Feeds in Turkey. J. Food Sci. Technol. 2020, 57, 787–793.
- Hashemzadeh, H.; Karbasi, A.; Mohammadi, H.; Firoozzare, A.; Boccia, F. Investigating the Effect of Nudges on Consumers’ Willingness to Pay for Genetically Modified Corn Oil. Sustainability 2022, 14, 12705.
- Boccia, F.; Punzo, G. A Choice Experiment on Consumer Perceptions of Three Generations of Genetically Modified Foods. Appetite 2021, 161, 105158.
- Zhang, R.; Ma, S.; Li, L.; Zhang, M.; Tian, S.; Wang, D.; Liu, K.; Liu, H.; Zhu, W.; Wang, X. Comprehensive Utilization of Corn Starch Processing By-Products: A Review. Grain Oil Sci. Technol. 2021, 4, 89–107.
- Delgado-Valerio, P.; Ramón-Amado, A.; Piñeyro-Nelson, A.; Álvarez-Buylla, E.R.; Ayala-Angulo, N.M.; Molina-Sánchez, A. Presencia de secuencias transgénicas en masa para tortillas de poblados urbanos y rurales de la meseta purépecha, michoacán, méxico. Rev. Fitotec. Mex. 2022, 45, 283.
- Ala-Kokko, K.; Nalley, L.L.; Shew, A.M.; Tack, J.B.; Chaminuka, P.; Matlock, M.D.; D’Haese, M. Economic and Ecosystem Impacts of GM Maize in South Africa. Glob. Food Secur. Agric. Policy Econ. Environ. 2021, 29, 100544.
- Brara, Z.; Costa, J.; Villa, C.; Grazina, L.; Bitam, A.; Mafra, I. Surveying Genetically Modified Maize in Foods Marketed in Algeria. Food Control 2020, 109, 106928.
- EUR-Lex. Regulation (EC) No 1830/2003 of the European Parliament and of the Council of 22 September 2003 Concerning the Traceability and Labelling of Genetically Modified Organisms and the Traceability of Food and Feed Products Produced from Genetically Modified Organisms and Amending Directive 2001/18/EC; Publications Office of the EU: Luxembourg, 2003; Volume 268.
- Decree of the Ministry of Agriculture of the People’s Republic of China (No. 10) Measures for the Administration of the Labeling of Agricultural Genetically Modified Organisms__State Council Gazette No. 2002 of 35_Chinese Government Website. Available online: https://www.gov.cn/gongbao/content/2002/content_61835.htm (accessed on 13 July 2023).
- 7 CFR Part 66—Part 66—National Bioengineered Food Disclosure Standard. Available online: https://www.law.cornell.edu/cfr/text/7/part-66 (accessed on 13 July 2023).
- Branch, L.S. Consolidated Federal Laws of Canada, Food and Drug Regulations. Available online: https://laws-lois.justice.gc.ca/eng/regulations/C.R.C.,_c._870/page-54.html#h-574622 (accessed on 13 July 2023).
- Federal Register of Legislation—Australian Government. Available online: https://www.legislation.gov.au/Series/F2015L00404 (accessed on 13 July 2023).
- FSSAI. Available online: https://fssai.gov.in/cms/food-safety-and-standards-regulations.php (accessed on 13 July 2023).
- Genesiska; Suratmi, R.C. Detection of Promoter Designed for Transgenic Plant in Local Soybean. IOP Conf. Ser. Earth Environ. Sci. 2020, 458, 012011.
- He, Y.; Fan, Z. A Novel Biosensor Based on DNA Hybridization for Ultrasensitive Detection of NOS Terminator Gene Sequences. Sens. Actuators B Chem. 2018, 257, 538–544.
- Zadeh, R.B.; Safaeian, S.; Moslemi, E.; Nadushen, R.M.; Esfahani, K. Monitoring of Infant Formula and Baby Food for the Pat and NOS Terminator of Genetically Modified Maize and Soybean by Real-Time PCR in Iran. Iran. J. Pharm. Res. 2022, 21, e126921.
- Jiao, P.; Jin, S.; Chen, N.; Wang, C.; Liu, S.; Qu, J.; Guan, S.; Ma, Y. Improvement of Cold Tolerance in Maize (Zea mays L.) Using Agrobacterium-Mediated Transformation of ZmSAMDC Gene. GM Crops Food-Biotechnol. Agric. Food Chain 2022, 13, 131–141.
- Zeng, H.; Wang, J.; Jia, J.; Wu, G.; Yang, Q.; Liu, X.; Tang, X. Development of a Lateral Flow Test Strip for Simultaneous Detection of BT-Cry1Ab, BT-Cry1Ac and CP4 EPSPS Proteins in Genetically Modified Crops. Food Chem. 2021, 335, 127627.
- Xu, J.-M.; Zhu, J.-S.; Li, M.-Z.; Hu, H.; Mao, C.-Z. Progress on Methods for Acquiring Flanking Genomic Sequence. Yi Chuan Hered. 2022, 44, 313–321.
- Siddique, K.; Wei, J.; Li, R.; Zhang, D.; Shi, J. Identification of T-DNA Insertion Site and Flanking Sequence of a Genetically Modified Maize Event IE09S034 Using Next-Generation Sequencing Technology. Mol. Biotechnol. 2019, 61, 694–702.
- Li, X.; Shen, K.; Yang, L.; Wang, S.; Pan, L.; Zhang, D. Applicability of a Novel Reference Molecule Suitable for Event-Specific Detections of Maize NK603 Based on Both 5′ and 3′ Flanking Sequences. Food Control 2010, 21, 927–934.
- Bhoge, R.K.; Chhabra, R.; Randhawa, G.; Sathiyabama, M.; Singh, M. Event-Specific Analytical Methods for Six Genetically Modified Maize Events Using Visual and Real-Time Loop-Mediated Isothermal Amplification. Food Control 2015, 55, 18–30.
- Tai, T.H.; Tanksley, S.D. A Rapid and Inexpensive Method for Isolation of Total DNA from Dehydrated Plant Tissue. Plant Mol. Biol. Report. 1990, 8, 297–303.
- Murray, M.G.; Thompson, W.F. Rapid Isolation of High Molecular Weight Plant DNA. Nucleic Acids Res. 1980, 8, 4321–4326.
- Giraffa, G.; Rossetti, L.; Neviani, E. An Evaluation of Chelex-Based DNA Purification Protocols for the Typing of Lactic Acid Bacteria. J. Microbiol. Methods 2000, 42, 175–184.
- Kim, C.S.; Lee, C.H.; Shin, J.S.; Chung, Y.S.; Hyung, N.I. A Simple and Rapid Method for Isolation of High Quality Genomic DNA from Fruit Trees and Conifers Using PVP. Nucleic Acids Res. 1997, 25, 1085–1086.
- Padmalatha, K.; Prasad, M. Optimization of DNA Isolation and PCR Protocol for RAPD Analysis of Selected Medicinal and Aromatic Plants of Conservation Concern from Peninsular India. Afr. J. Biotechnol. 2006, 5, 230–234.
- Bashalkhanov, S.; Rajora, O.P. Protocol: A High-Throughput DNA Extraction System Suitable for Conifers. Plant Methods 2008, 4, 20.
- Russo, A.; Mayjonade, B.; Frei, D.; Potente, G.; Kellenberger, R.T.; Frachon, L.; Copetti, D.; Studer, B.; Frey, J.E.; Grossniklaus, U.; et al. Low-Input High-Molecular-Weight DNA Extraction for Long-Read Sequencing From Plants of Diverse Families. Front. Plant Sci. 2022, 13, 883897.
- Nadeem, S.A.; Mughal, D.; Butt, N.A.; Ahmed, S.; Khan, I.A. Utilization of Corn Silk for GMO Detection Through Real-Time PCR. Waste Biomass Valorization 2023.
- SanJuan-Badillo, A.; Galvez, A.; Plasencia, J.; Quirasco, M. Assessment of DNA extraction methods from various maize (Zea mays L.) tissues for environmental GMO monitoring in Mexico. Part I: Detection by end-point PCR. Agrociencia 2014, 48, 17–33.
- Gao, S.; Martinez, C.; Skinner, D.J.; Krivanek, A.F.; Crouch, J.H.; Xu, Y. Development of a Seed DNA-Based Genotyping System for Marker-Assisted Selection in Maize. Mol. Breed. 2008, 22, 477–494.
- Leach, K.A.; McSteen, P.C.; Braun, D.M. Genomic DNA Isolation from Maize (Zea mays) Leaves Using a Simple, High-Throughput Protocol. Curr. Protoc. Plant Biol. 2016, 1, 15–27.
- Matthes, N.; Westphal, K.; Haldemann, C.; Egert, M.; Jokisch, C.; Speck, B. Validation of a Modified CTAB Method for DNA Extraction from Protein-Rich Maize Feedstuffs. J. Consum. Prot. Food Saf. 2020, 15, 331–340.
- Turkec, A.; Kazan, H.; Karacanli, B.; Lucas, S.J. DNA Extraction Techniques Compared for Accurate Detection of Genetically Modified Organisms (GMOs) in Maize Food and Feed Products. J. Food Sci. Technol. 2015, 52, 5164–5171.
- Takabatake, R.; Noritake, H.; Noguchi, A.; Nakamura, K.; Kondo, K.; Akiyama, H.; Teshima, R.; Mano, J.; Kitta, K. Comparison of DNA Extraction Methods for Sweet Corn and Processed Sweet Corns. Food Hyg. Saf. Sci. 2013, 54, 309–315.
- Kishine, M.; Noguchi, A.; Mano, J.; Takabatake, R.; Nakamura, K.; Kondo, K.; Kitta, K. Detection of DNA in Highly Processed Foods. Food Hyg. Saf. Sci. 2018, 59, 151–156.
- Singh, M.; Sodhi, K.K.; Paliwal, A.; Sharma, S.; Randhawa, G. Efficient DNA Extraction Procedures for Processed Food Derivatives-a Critical Step to Ensure Quality for GMO Analysis. Food Anal. Methods 2021, 14, 2249–2261.
- Liu, Y.; Liu, X.; Cui, Y.; Yuan, W. Ultrasound for Microalgal Cell Disruption and Product Extraction: A Review. Ultrason. Sonochem. 2022, 87, 106054.
- Xia, Y.; Chen, F.; Du, Y.; Liu, C.; Bu, G.; Xin, Y.; Liu, B. A Modified SDS-Based DNA Extraction Method from Raw Soybean. Biosci. Rep. 2019, 39, BSR20182271.
- Jones, A.S. The Isolation of Bacterial Nucleic Acids Using Cetyltrimethylammonium Bromide (Cetavlon). Biochim. Biophys. Acta 1953, 10, 607–612.
- Masoodi, K.Z.; Lone, S.M.; Rasool, R.S. Chapter 7—Genomic DNA Extraction from the Plant Leaves Using the CTAB Method. In Advanced Methods in Molecular Biology and Biotechnology; Masoodi, K.Z., Lone, S.M., Rasool, R.S., Eds.; Academic Press: Cambridge, MA, USA, 2021; pp. 37–44. ISBN 978-0-12-824449-4.
- Chabi Sika, K.; Kefela, T.; Adoukonou-Sagbadja, H.; Ahoton, L.; Saidou, A.; Baba-Moussa, L.; Jno Baptiste, L.; Kotconi, S.O.; Gachomo, E.W. A Simple and Efficient Genomic DNA Extraction Protocol for Large Scale Genetic Analyses of Plant Biological Systems. Plant Gene 2015, 1, 43–45.
- Wang, X.; Liu, M.; Ding, X. Guanidinium Hydrophobic Magnetic Ionic Liquid-Based Dispersive Droplet Extraction for the Selective Extraction of DNA. Langmuir 2021, 37, 11665–11675.
- Emaus, M.N.; Cagliero, C.; Gostel, M.R.; Johnson, G.; Anderson, J.L. Simple and Efficient Isolation of Plant Genomic DNA Using Magnetic Ionic Liquids. Plant Methods 2022, 18, 37.
- Marengo, A.; Cagliero, C.; Sgorbini, B.; Anderson, J.L.; Emaus, M.N.; Bicchi, C.; Bertea, C.M.; Rubiolo, P. Development of an Innovative and Sustainable One-Step Method for Rapid Plant DNA Isolation for Targeted PCR Using Magnetic Ionic Liquids. Plant Methods 2019, 15, 1–11.
- Choi, S.-E.; Khoo, H.; Hur, S.C. Recent Advances in Microscale Electroporation. Chem. Rev. 2022, 122, 11247–11286.
- Wu, X.; Gong, F.; Wang, W. Protein Extraction from Plant Tissues for 2DE and Its Application in Proteomic Analysis. Proteomics 2014, 14, 645–658.
- Nishii, K.; Möller, M.; Foster, R.G.; Forrest, L.L.; Kelso, N.; Barber, S.; Howard, C.; Hart, M.L. A High Quality, High Molecular Weight DNA Extraction Method for PacBio HiFi Genome Sequencing of Recalcitrant Plants. Plant Methods 2023, 19, 41.
- Barbier, F.F.; Chabikwa, T.G.; Ahsan, M.U.; Cook, S.E.; Powell, R.; Tanurdzic, M.; Beveridge, C.A. A Phenol/Chloroform-Free Method to Extract Nucleic Acids from Recalcitrant, Woody Tropical Species for Gene Expression and Sequencing. Plant Methods 2019, 15, 62.
- Valizadeh, N.; Holasou, H.A.; Mohammadi, S.A.; Khawar, K.M. A Comparison of Genomic DNA Extraction Protocols in Artemisia Annua L. for Large Scale Genetic Analyses Studies. Iran. J. Sci. Technol. Trans. Sci. 2021, 45, 1587–1595.
- Springer, N.M. Isolation of Plant DNA for PCR and Genotyping Using Organic Extraction and CTAB. Cold Spring Harb. Protoc. 2010, 2010, pdb.prot5515.
- Spadoni, A.; Sion, S.; Gadaleta, S.; Savoia, M.A.; Piarulli, L.; Fanelli, V.; di Rienzo, V.; Taranto, F.; Miazzi, M.; Montemurro, C.; et al. A Simple and Rapid Method for Genomic DNA Extraction and Microsatellite Analysis in Tree Plants. J. Agric. Sci. Technol. 2019, 21, 1215–1226.
- Koh, R.B.L.; Barbosa, C.F.C.; Aquino, V.M.; Galvez, L.C. Extraction of High Molecular Weight DNA Suitable for Next-Generation Sequencing from the Fiber Crop Abaca. Ind. Crops Prod. 2021, 161, 113194.
- Ali, Q.; Salisu, I.B.; Raza, A.; Shahid, A.A.; Rao, A.Q.; Husnain, T. A Modified Protocol for Rapid DNA Isolation from Cotton (Gossypium spp.). Methods X 2019, 6, 259–264.
- Lee, B.-J.; Kim, S.; Lee, J.-W.; Lee, H.-M.; Eo, S.H. Technical Note: Polyvinylpyrrolidone (PVP) and Proteinase-K Improve the Efficiency of DNA Extraction from Japanese Larch Wood and PCR Success Rate. Forensic Sci. Int. 2021, 328, 111005.
- Brandão, W.Q.; da Silva, R.J.; Mojica-Sánchez, L.C.; Maciel, B.G.; Ratkovski, G.P.; de Melo, C.P. Use of Polypyrrole-Polystyrene Membranes for Extracting DNA from Plant Tissues. Biomater. Biosyst. 2022, 7, 100060.
- Natarajan, V.P.; Zhang, X.; Morono, Y.; Inagaki, F.; Wang, F. A Modified SDS-Based DNA Extraction Method for High Quality Environmental DNA from Seafloor Environments. Front. Microbiol. 2016, 7, 986.
- Allen, G.C.; Flores-Vergara, M.A.; Krasynanski, S.; Kumar, S.; Thompson, W.F. A Modified Protocol for Rapid DNA Isolation from Plant Tissues Using Cetyltrimethylammonium Bromide. Nat. Protoc. 2006, 1, 2320–2325.
- Jiang, Q.; Li, Y.; Huang, L.; Guo, J.; Wang, A.; Ma, C.; Shi, C. Direct Capture and Amplification of Nucleic Acids Using a Universal, Elution-Free Magnetic Bead-Based Method for Rapid Pathogen Detection in Multiple Types of Biological Samples. Anal. Bioanal. Chem. 2023, 415, 427–438.
- Oberacker, P.; Stepper, P.; Bond, D.M.; Hoehn, S.; Focken, J.; Meyer, V.; Schelle, L.; Sugrue, V.J.; Jeunen, G.-J.; Moser, T.; et al. Bio-On-Magnetic-Beads (BOMB): Open Platform for High-Throughput Nucleic Acid Extraction and Manipulation. PLoS Biol. 2019, 17, e3000107.
- Fort, A.; Guiry, M.D.; Sulpice, R. Magnetic Beads, a Particularly Effective Novel Method for Extraction of NGS-Ready DNA from Macroalgae. Algal Res. 2018, 32, 308–313.
- Fei, Z.; Cheng, C.; Wei, R.; Tan, G.; Xiao, P. Reversible Superhydrophobicity Unyielding Magnetic Beads of Flipping-Triggered (SYMBOL) Regulate the Binding and Unbinding of Nucleic Acids for Ultra-Sensitive Detection. Chem. Eng. J. 2022, 431, 133953.
- Balakrishnan, S.G.; Ahmad, M.R.; Koloor, S.S.R.; Petrů, M. Separation of CtDNA by Superparamagnetic Bead Particles in Microfluidic Platform for Early Cancer Detection. J. Adv. Res. 2021, 33, 109–116.
- Hin, S.; Paust, N.; Rombach, M.; Lueddecke, J.; Specht, M.; Zengerle, R.; Mitsakakis, K. Magnetophoresis in Centrifugal Microfluidics at Continuous Rotation for Nucleic Acid Extraction. Micromachines 2022, 13, 2112.
- Wu, J.; Kodzius, R.; Cao, W.; Wen, W. Extraction, Amplification and Detection of DNA in Microfluidic Chip-Based Assays. Microchim. Acta 2014, 181, 1611–1631.
- Tian, W.; Liu, Y.; Wang, S.; Ye, J.; Liu, H.; Wang, Y.; Zhou, M. Automated and Rapid Easy-to-Use Magnetic Solid-Phase Extraction System for Five Heavy Metals in Cereals and Feeds. Foods 2022, 11, 3944.
- Tong, R.; Zhang, L.; Hu, C.; Chen, X.; Song, Q.; Lou, K.; Tang, X.; Chen, Y.; Gong, X.; Gao, Y.; et al. An Automated and Miniaturized Rotating-Disk Device for Rapid Nucleic Acid Extraction. Micromachines 2019, 10, 204.
- Li, J.; Gao, H.; Li, Y.; Xiao, F.; Zhai, S.; Wu, G.; Wu, Y. Event-Specific PCR Methods to Quantify the Genetically Modified DBN9936 Maize. J. Food Compos. Anal. 2022, 105, 104236.
- Spanea, E.; Tsironi, T.; Tsakali, E.; Batrinou, A.; Stefanou, V.; Antonopoulos, D.; Koussissis, S.; Tsaknis, J.; Impe, J.V.; Houhoula, D. Evaluation of a Real Time PCR Assay Method for the Detection of Genetically Modified Organisms in Food Products. J. Food Res. 2020, 9, 1.
- Kaur, J.; Radu, S.; Ghazali, F.M.; Kqueen, C.Y. Real-Time PCR-Based Detection and Quantification of Genetically Modified Maize in Processed Feeds Commercialised in Malaysia. Food Control 2010, 21, 1536–1544.
- Dong, S.; Zhang, D.; Yu, C.; Zhang, Z.; Liu, Y. Using Droplet Digital PCR to Detect Plant DNA in Tissues of Zebrafish (Danio Rerio) Fed Genetically Modified Maize. Aquac. Res. 2021, 52, 4467–4474.
- Zimmermann, A.; Hemmer, W.; Liniger, M.; Luthy, J.; Pauli, U. A Sensitive Detection Method for Genetically Modified MaisGard (TM) Corn Using a Nested PCR-System. Food Sci. Technol.-Lebensm.-Wiss. Technol. 1998, 31, 664–667.
- Dong, L.; Long, L.; Xing, Z.; Li, C.; He, Y.; Yan, W.; Xia, W.; Li, F. Establishment of Multi-Fluorescence Real-Time PCR Assay for GM Soybean Detection. Int. J. Agric. Biol. 2020, 24, 292–298.
- Godbey, W.T. Chapter 10—The Polymerase Chain Reaction (PCR). In Biotechnology and its Applications, 2nd ed.; Godbey, W.T., Ed.; Academic Press: Cambridge, MA, USA, 2022; pp. 219–246. ISBN 978-0-12-817726-6.
- Stephenson, F.H. Chapter 8—The Polymerase Chain Reaction. In Calculations for Molecular Biology and Biotechnology, 3rd ed.; Stephenson, F.H., Ed.; Academic Press: Cambridge, MA, USA, 2016; pp. 171–213. ISBN 978-0-12-802211-5.
- Dobnik, D.; Spilsberg, B.; Bogožalec Košir, A.; Štebih, D.; Morisset, D.; Holst-Jensen, A.; Žel, J. Multiplex Droplet Digital PCR Protocols for Quantification of GM Maize Events. Methods Mol. Biol. Clifton NJ 2018, 1768, 69–98.
- Heid, C.A.; Stevens, J.; Livak, K.J.; Williams, P.M. Real Time Quantitative PCR. Genome Res. 1996, 6, 986–994.
- Fengjun, W.; Qiang, C.; Sudan, Y.; Haifeng, X.; Hongxia, X.; Yiping, Y.; Yexi, Z. Rapid Identification of Transgenic Maize Lines and Transformants by Quadruplex Real-Time PCR. Zhongguo Liangyou Xuebao 2021, 36, 128–135.
- McNutt, M.K.; Baulcombe, D.; Benfey, P.N.; Bergmann, D.; Coruzzi, G.M.; Estelle, M.; Kieber, J.J.; Long, S.P.; Niyogi, K.K.; Schroeder, J.I.; et al. Digital PCR. Proc. Natl. Acad. Sci. USA 1999, 96, 9236–9241.
- Tellinghuisen, J. DPCR vs. QPCR: The Role of Poisson Statistics at Low Concentrations. Anal. Biochem. 2020, 611, 113946.
- Chen, S.; Sun, Y.; Fan, F.; Chen, S.; Zhang, Y.; Zhang, Y.; Meng, X.; Lin, J.-M. Present Status of Microfluidic PCR Chip in Nucleic Acid Detection and Future Perspective. TrAC Trends Anal. Chem. 2022, 157, 116737.
- Bogožalec Košir, A.; Demšar, T.; Štebih, D.; Žel, J.; Milavec, M. Digital PCR as an Effective Tool for GMO Quantification in Complex Matrices. Food Chem. 2019, 294, 73–78.
- Dong, L.; Meng, Y.; Sui, Z.; Wang, J.; Wu, L.; Fu, B. Comparison of Four Digital PCR Platforms for Accurate Quantification of DNA Copy Number of a Certified Plasmid DNA Reference Material. Sci. Rep. 2015, 5, 13174.
- Wan, J.; Song, L.; Wu, Y.; Brzoska, P.; Keys, D.; Chen, C.; Valliyodan, B.; Shannon, J.G.; Nguyen, H.T. Application of Digital PCR in the Analysis of Transgenic Soybean Plants. Adv. Biosci. Biotechnol. 2016, 7, 403–417.
- Demeke, T.; Dobnik, D. Critical Assessment of Digital PCR for the Detection and Quantification of Genetically Modified Organisms. Anal. Bioanal. Chem. 2018, 410, 4039–4050.
- Kahkeshani, S.; Kong, J.E.; Wei, Q.; Tseng, D.; Garner, O.B.; Ozcan, A.; Di Carlo, D. Ferrodrop Dose-Optimized Digital Quantification of Biomolecules in Low-Volume Samples. Anal. Chem. 2018, 90, 8881–8888.
- Park, J.; Han, D.H.; Hwang, S.-H.; Park, J.-K. Reciprocating Flow-Assisted Nucleic Acid Purification Using a Finger-Actuated Microfluidic Device. Lab. Chip 2020, 20, 3346–3353.
- Park, J.; Lee, K.G.; Han, D.H.; Lee, J.-S.; Lee, S.J.; Park, J.-K. Pushbutton-Activated Microfluidic Dropenser for Droplet Digital PCR. Biosens. Bioelectron. 2021, 181, 113159.
- Cottenet, G.; Blancpain, C.; Chuah, P.F. Performance Assessment of Digital PCR for the Quantification of GM-Maize and GM-Soya Events. Anal. Bioanal. Chem. 2019, 411, 2461–2469.
- Shen, C.; Yin, H.; Tong, Z.; Qiu, S.; Lu, Y.; Wu, Z.; Mao, H. Digital Microfluidic Chip Based on Direct Ink Writing For Nucleic Acid Multiplex PCR Detection. In Proceedings of the 2022 IEEE 35th International Conference on Micro Electro Mechanical Systems Conference (MEMS), Tokyo, Japan, 9–13 January 2022; pp. 365–368.
- Shin, M.K.; Jeon, S.M.; Koo, Y.E. Development of a Rapid Detection Method for Genetically Modified Rice Using the Ultra-Fast PCR System. Food Sci. Biotechnol. 2022, 31, 175–182.
- Zhou, H.; Lei, Y.; Wang, P.; Liu, M.; Hu, X. Development of SYBR Green Real-Time PCR and Nested RT-PCR for the Detection of Potato Mop-Top Virus (PMTV) and Viral Surveys in Progeny Tubers Derived from PMTV Infected Potato Tubers. Mol. Cell. Probes 2019, 47, 101438.
- Notomi, T.; Okayama, H.; Masubuchi, H.; Yonekawa, T.; Watanabe, K.; Amino, N.; Hase, T. Loop-Mediated Isothermal Amplification of DNA. Nucleic Acids Res. 2000, 28, E63.
- Lizardi, P.M.; Huang, X.; Zhu, Z.; Bray-Ward, P.; Thomas, D.C.; Ward, D.C. Mutation Detection and Single-Molecule Counting Using Isothermal Rolling-Circle Amplification. Nat. Genet. 1998, 19, 225–232.
- Xu, G.; Hu, L.; Zhong, H.; Wang, H.; Yusa, S.; Weiss, T.C.; Romaniuk, P.J.; Pickerill, S.; You, Q. Cross Priming Amplification: Mechanism and Optimization for Isothermal DNA Amplification. Sci. Rep. 2012, 2, 246.
- Compton, J. Nucleic-Acid Sequence-Based Amplification. Nature 1991, 350, 91–92.
- Lasken, R.S. Genomic DNA Amplification by the Multiple Displacement Amplification (MDA) Method. Biochem. Soc. Trans. 2009, 37, 450–453.
- Vincent, M.; Xu, Y.; Kong, H.M. Helicase-Dependent Isothermal DNA Amplification. Embo Rep. 2004, 5, 795–800.
- Lobato, I.M.; O’Sullivan, C.K. Recombinase Polymerase Amplification: Basics, Applications and Recent Advances. TrAC Trends Anal. Chem. 2018, 98, 19–35.
- Chalmers, F.M.; Curnow, K.M. Scaling Up the Ligase Chain Reaction-Based Approach to Gene Synthesis. BioTechniques 2001, 30, 249–252.
- Ehses, S.; Ackermann, J.; McCaskill, J.S. Optimization and Design of Oligonucleotide Setup for Strand Displacement Amplification. J. Biochem. Biophys. Methods 2005, 63, 170–186.
- Toley, B.J.; Covelli, I.; Belousov, Y.; Ramachandran, S.; Kline, E.; Scarr, N.; Vermeulen, N.; Mahoney, W.; Lutz, B.R.; Yager, P. Isothermal Strand Displacement Amplification (ISDA): A Rapid and Sensitive Method of Nucleic Acid Amplification for Point-of-Care Diagnosis. Analyst 2015, 140, 7540–7549.
- Singh, M.; Pal, D.; Sood, P.; Randhawa, G. Loop-Mediated Isothermal Amplification Assays: Rapid and Efficient Diagnostics for Genetically Modified Crops. Food Control 2019, 106, 106759.
- Xu, J.; Zheng, Q.; Yu, L.; Liu, R.; Zhao, X.; Wang, G.; Wang, Q.; Cao, J. Loop-Mediated Isothermal Amplification (LAMP) Method for Detection of Genetically Modified Maize T25. Food Sci. Nutr. 2013, 1, 432–438.
- Li, R.; Chen, J.; Zhang, X.; Cui, J.; Tao, S.; Yang, L. Mini-Disk Capillary Array Coupling with LAMP for Visual Detection of Multiple Nucleic Acids Using Genetically Modified Organism Analysis as an Example. J. Agric. Food Chem. 2020, 68, 899–906.
- Kaymaz, S.V.; Elitas, M. Optimization of Loop-Mediated Isothermal Amplification (LAMP) Reaction Mixture for Biosensor Applications. MethodsX 2021, 8, 101282.
- Piepenburg, O.; Williams, C.H.; Stemple, D.L.; Armes, N.A. DNA Detection Using Recombination Proteins. PLoS Biol. 2006, 4, e204.
- Choi, G.; Jung, J.H.; Park, B.H.; Oh, S.J.; Seo, J.H.; Choi, J.S.; Kim, D.H.; Seo, T.S. A Centrifugal Direct Recombinase Polymerase Amplification (Direct-RPA) Microdevice for Multiplex and Real-Time Identification of Food Poisoning Bacteria. Lab. Chip 2016, 16, 2309–2316.
- Chandu, D.; Paul, S.; Parker, M.; Dudin, Y.; King-Sitzes, J.; Perez, T.; Mittanck, D.W.; Shah, M.; Glenn, K.C.; Piepenburg, O. Development of a Rapid Point-of-Use DNA Test for the Screening of Genuity® Roundup Ready 2 Yield® Soybean in Seed Samples. BioMed Res. Int. 2016, 2016, 3145921.
- Daher, R.K.; Stewart, G.; Boissinot, M.; Bergeron, M.G. Recombinase Polymerase Amplification for Diagnostic Applications. Clin. Chem. 2016, 62, 947–958.
- Versatile Isothermal DNA/RNA Amplification by TwistDx. Available online: https://www.twistdx.co.uk/ (accessed on 17 July 2023).
- Lin, K.; Guo, J.; Guo, X.; Li, Q.; Li, X.; Sun, Z.; Zhao, Z.; Weng, J.; Wu, J.; Zhang, R.; et al. Fast and Visual Detection of Nucleic Acids Using a One-Step RPA-CRISPR Detection (ORCD) System Unrestricted by the PAM. Anal. Chim. Acta 2023, 1248, 340938.
- Li, K.; Luo, Y.; Huang, K.; Yang, Z.; Wan, Y.; Xu, W. Single Universal Primer Recombinase Polymerase Amplification-Based Lateral Flow Biosensor (SUP-RPA-LFB) for Multiplex Detection of Genetically Modified Maize. Anal. Chim. Acta 2020, 1127, 217–224.
- Liu, R.; Wang, Z.; Liu, X.; Chen, A.; Yang, S. Rapid On-Site Detection of Salmonella Pullorum Based on Lateral Flow Nucleic Acid Assay Combined with Recombinase Polymerase Amplification Reaction. Poult. Sci. 2020, 99, 7225–7232.
- Seder, I.; Coronel-Tellez, R.; Helalat, S.H.; Sun, Y. Fully Integrated Sample-in-Answer-out Platform for Viral Detection Using Digital Reverse Transcription Recombinase Polymerase Amplification (DRT-RPA). Biosens. Bioelectron. 2023, 237, 115487.
- Tortajada-Genaro, L.A.; Maquieira, A. Multiple Recombinase Polymerase Amplification and Low-Cost Array Technology for the Screening of Genetically Modified Organisms. J. Food Compos. Anal. 2021, 103, 104083.
- Glökler, J.; Lim, T.S.; Ida, J.; Frohme, M. Isothermal Amplifications—A Comprehensive Review on Current Methods. Crit. Rev. Biochem. Mol. Biol. 2021, 56, 543–586.
- Wang, Y.; Sun, L.; Li, J.; Wang, Z.; Jiao, W.; Xiao, J.; Shen, C.; Xu, F.; Qi, H.; Wang, Y.; et al. Label-Free Cross-Priming Amplification Coupled with Endonuclease Restriction and Nanoparticles-Based Biosensor for Simultaneous Detection of Nucleic Acids and Prevention of Carryover Contamination. Front. Chem. 2019, 7, 322.
- Nguyen, H.Q.; Nguyen, V.D.; Van Nguyen, H.; Seo, T.S. Quantification of Colorimetric Isothermal Amplification on the Smartphone and Its Open-Source App for Point-of-Care Pathogen Detection. Sci. Rep. 2020, 10, 15123.
- Turkec, A.; Lucas, S.J.; Karacanli, B.; Baykut, A.; Yuksel, H. Assessment of a Direct Hybridization Microarray Strategy for Comprehensive Monitoring of Genetically Modified Organisms (GMOs). Food Chem. 2016, 194, 399–409.
- Wei, B.-R.; Savellano, D.F.; Hu, C.-H. Analysis of Status Quo and Research Progress in Nursing Care for Different Typed Coronavirus Disease 2019. J. Integr. Nurs. 2020, 2, 160–171.
- Morisset, D.; Dobnik, D.; Hamels, S.; Zel, J.; Gruden, K. NAIMA: Target Amplification Strategy Allowing Quantitative on-Chip Detection of GMOs. Nucleic Acids Res. 2008, 36, e118.
- Ki, E.Y.; Lee, Y.K.; Lee, A.; Park, J.S. Comparison of the PANArray HPV Genotyping Chip Test with the Cobas 4800 HPV and Hybrid Capture 2 Tests for Detection of HPV in ASCUS Women. Yonsei Med. J. 2018, 59, 662–668.
- Lu, X.-B.; Wu, H.-B.; Wang, M.; Li, B.-D.; Yang, C.-L.; Sun, H.-W. Developing a Method of Oligonucleotide Microarray for Event Specific Detec-Tion of Transgenic Maize. ACTA Agron. Sin. 2009, 35, 1432–1438.
- Liu-Stratton, Y.; Roy, S.; Sen, C.K. DNA Microarray Technology in Nutraceutical and Food Safety. Toxicol. Lett. 2004, 150, 29–42.
- Gao, H.; Yan, C.; Wu, W.; Li, J. Application of Microfluidic Chip Technology in Food Safety Sensing. Sensors 2020, 20, 1792.
More