The advantages of human milk feeding, especially in preterm babies, are well recognized. Infants’ feeding with breast milk lowers the likelihood of developing a diverse range of non-communicable diseases later in life and it is also associated with improved neurodevelopmental outcomes. Although the precise mechanisms through which human milk feeding is linked with infants’ neurodevelopment are still unknown, potential epigenetic effects of breast milk through its bioactive components, including non-coding RNAs, stem cells and microbiome, could at least partly explain this association. Micro- and long-non-coding RNAs, enclosed in milk exosomes, as well as breast milk stem cells, survive digestion, reach the circulation and can cross the blood–brain barrier. Certain non-coding RNAs potentially regulate genes implicated in brain development and function, whereas nestin-positive stem cells can possibly differentiate into neural cells or/and act as epigenetic regulators in the brain.
- epigenetics
- human milk
- neurodevelopment
- miRNAs
- long non-coding RNAs
- stem cells
- microbiome
1. Introduction
2. MiRNAs
A number of recent publications have demonstrated that human milk contains components recently described as extracellular vesicles (EVs) [29]. Extracellular vesicles is a term for all phospholipid bilayer-enclosed particles that are released by cells into their environment and include exosomes and microvesicles [29]. Exosomes carry bioactive substances like proteins, DNA, messenger RNA (mRNA) and miRNAs [29][30]. Breast milk exosomes, being resistant to digestion [31], are able to transport their cargo and miRNAs to peripheral tissues via the systemic circulation and facilitate the epigenetic programming of various tissues and organs [20]. For this reason, they are considered important signaling molecules (signalosomes) between mother and child [20][32]. Since exosomes are also able to cross the blood–brain barrier, it is possible that the positive impact of breast milk on neurodevelopment is associated with miRNAs’ activity [33]. MiRNAs are small, single-stranded, non-coding RNA molecules containing 18 to 25 nucleotides. MiRNAs are also found in plants, animals and viruses, among others [34], and they are capable of controlling up to 60% of gene expression [35][36] by inhibiting mRNA translation into protein. These particles are, thus, involved in post-transcriptional gene regulation [37][38][39]. Breast milk has been categorized as one of the biological fluids that possesses a high concentration of miRNAs encapsulated in exosomes or as free molecules, with more than 1400 distinct miRNAs identified [36][40]. Not only is human milk highly enriched in miRNAs, but it has also the highest concentration of miRNAs compared to other body fluids, including plasma [20][40]. While previous research was focused on analyzing miRNAs in the skim fraction of breast milk, recent studies investigating the lipid and cell fractions of milk have revealed a larger quantity and diversity of miRNAs compared to the skim fraction [36]. A systematic review of 30 studies on non-coding RNAs of human breast milk showed that 10 miRNAs, including miR-148a-3p, miR-30a-5p, miR-30d-5p, miR-22-3p, miR-146b-5p, miR-200a-3p, miR-200c-3p, let-7a-5p, let-7b-5p and let-7f-5p, were the most abundant miRNAs in all breast milk fractions examined [19]. Overall, these findings show that miRNAs, which are well-established epigenetic modulators, are abundant in human breast milk and they are influenced by several factors relevant to lactation per se, maternal health and disease and preterm birth. They can reach the brain by crossing the blood–brain barrier, whereas several of them possess neuroprotective effects and can regulate the expression of genes implicated in infants’ brain development and function (Figure 1).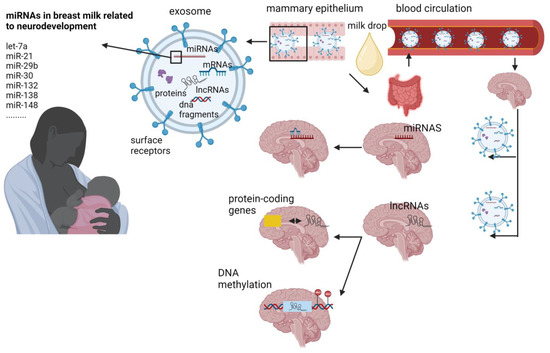
3. Long Non-Coding RNAs
In addition to miRNAs, breast milk also contains other types of regulatory non-coding RNAs, such as long non-coding RNAs (lncRNAs). Long non-coding RNAs are RNA molecules that are typically composed of at least 200 nucleotides [41]. They are often formed through the splicing of two or more exons derived from genomic regions located near protein-coding genes [19]. LncRNAs have a crucial role in processes such as neurogenesis, synaptogenesis, and the development of the brain (Figure 1). The utilization of high-throughput technologies has revealed their specific expression in distinct cell types, subcellular compartments, and various brain regions [42][43]. Numerous lncRNAs exhibit expression patterns that vary with age [44] and actively contribute to the determination of neural cell fate [45]. Given their involvement in these essential processes, any abnormal expression of these transcripts has the potential to lead to neurodevelopmental or neuropsychiatric disorders, including, but not limited to, autism spectrum disorder and schizophrenia [45][46]. Compared to miRNAs, lncRNAs of breast milk have been much less studied to date and only from an immunological and metabolic point of view. NORAD, referred to as “the guardian of the human genome” and shown to have a neuroprotective epigenetic role, was found to be abundant in human breast milk; however, it was downregulated in preterm compared to term human milk. Further studies are needed to investigate human milk non-coding RNAs related to brain development and neurodevelopment in full-term and preterm babies.4. Stem Cells
Stem cells possess a remarkable capacity for both self-renewal, sustaining their undifferentiated state, and differentiation into various cell types and tissues in specific conditions [47][48][49]. In contrast, adult cells traditionally maintain their lineage commitment, yet recent studies have revealed promising approaches to induce cellular plasticity, allowing them to potentially transform into diverse cell types. This breakthrough holds significant implications for cell-based therapies in the field of regenerative medicine [49]. The discovery of stem cells within human milk dates back to 2007 [50], highlighting their presence in this unique fluid. Breastfeeding has long been recognized for its protective effects against diseases that may arise later in life, although their precise mechanism remains elusive. The presence of stem cells in both preterm and term human breast milk [51] offers one potential explanation for these beneficial effects. Interestingly, in animal studies, breast milk stem cells survive digestion and enter into the circulation and the brain, where they can be differentiated into neuronal and glial cells [52]. Stem cells from human milk contain both genetic material and bioactive molecules, such as microRNAs, which can act as epigenetic regulators [53]. The beneficial effects of breast milk stem cells may also be mediated through the paracrine action of exosomes released by these cells [54][55]. Moreover, by using the marker nestin, Cregan et al. identified nestin-positive putative stem cells in human breast milk [50]. Nestin (acronym for neuroepithelial stem cell protein) is a marker for multipotent stem cells that can differentiate into neural cells [56]. Indeed, Hosseini et al. [57] showed that human breast milk derived stem cells can differentiate into neural lineages (oligodendrocytes, astrocytes, and neurons). This differentiation capacity of milk stem cells offers valuable insights into the beneficial effects of human milk on neurodevelopment. That discovery also indicates the potential use of these cells as a suitable and easy source for cell replacement therapies targeting brain diseases. Thus, breast milk stem cells, either through their differentiation into neural cells or/and by acting as epigenetic regulators in the brain (Figure 2), seem to have opened up new horizons in the explanation of the positive short- and long-term impact of human milk. However, further research is required to elucidate their exact mechanism(s) of action after breastfeeding and define the extent of their capabilities.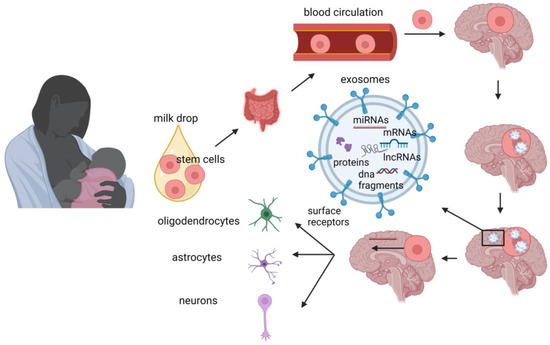
5. Microbiome
The microbiome, which encompasses the genomes of all microorganisms, symbiotic and pathogenic, in a specific environment, has been extensively studied [58][59]. Previous assumptions regarding the existence of bacteria in human milk attributed their presence to contamination or mastitis [60][61]. However, during the early 2000s, research emerged revealing the existence of commensal bacteria in human milk and provided evidence that the DNA of these bacteria differed from that found on the surface of the breast skin, indicating that they were distinct entities [62][63][64]. By using next-generation sequencing techniques, it was found that half of the microorganism population was the same in all milk samples composing the core bacterial microbiota (bacteriome) [65]. The predominant phyla reported in human milk are Proteobacteria, Firmicutes, Actinobacteria, and Bacteroidetes. When examining the genus level, the most abundant taxa include Bifidobacterium, Lactobacillus, Streptococcus, Staphylococcus, Ralstonia, Bacteroides, Enterobacter, and Enterococcus, among others [66][67]. The composition of breast milk microbiota may be influenced by various factors. Among them, the impact of the stage of lactation on the composition of microbiota in breast milk has been investigated in several studies [68][69][70][71][72]. Findings have been inconsistent, with some studies reporting higher total bacterial loads in colostrum compared to mature milk [68][69], while others have observed an increase in bacterial loads throughout the lactation period [70][71]. On the contrary, certain studies did not detect significant alterations in bacterial numbers in breast milk samples collected within the first month after delivery, suggesting stability in microbial composition during this early period [72]. These varying results highlight the complexity and diversity of microbiota present in breast milk. The complexity and diversity of breast milk microbiota have implications for understanding the influence of other factors on its composition. Probiotic administration during pregnancy did not influence the composition of the microbiota of breast milk, according to three separate studies involving participant sizes of 84, 125, and 20 women [67][73][74][75]. Similarly, the impact of smoking on the diversity and composition of the breast milk microbiota was examined in a study involving 393 participants, revealing no significant effects [66]. When considering milk expression methods, it was observed that using a breast pump rather than manual expression was associated with lower bacterial richness in breast milk; this could be attributed to the non-aseptic protocol used for milk collection [66]. There is evidence that the gut microbiome during early life contributes to the establishment of epigenetic modifications and it is also associated with brain development and neurodevelopment [76][77][78]. The colonization of the infant’s intestine after birth, influenced by maternal flora, delivery method, early skin-to-skin contact, and neonatal diet, results in specific epigenetic patterns that can influence the protective function of the gut mucosa against future insults [79]. Furthermore, the gut microorganisms secrete molecules which can reach the brain via the circulatory system after absorption and affect the brain’s development (Figure 3), especially during sensitive periods (gut–brain axis) [80]. Interestingly, in a recent study in a humanized mouse model, the aberrant gut microbiome of preterm infants had negative effects on brain organization and maturation, and brain metabolism, as well as on behavior and memory [77]. The connection between the gut microbiome and brain function has led to investigations into its potential role in neurobehavioral disorders, such as autism spectrum disorder (ASD), anxiety and attention-deficit-hyperactivity disorder [81]. It has been reported that children with ASD have a dysbiotic microbiome with an abundance of Bacteroidetes in feces [82]. The presence of these bacteria in fecal samples could potentially explain the occurrence of gastrointestinal symptoms in certain individuals with ASD [83][84]. As neurodevelopmental impairments are often linked to the degree of prematurity, optimizing the microbial environment in early life becomes crucial for promoting healthy neurodevelopment in this vulnerable population [85]. Considering that the maternal breast milk microbiome colonizes the infant’s gut and presents similar species to the gut microbiome of the infants, it can possibly be extrapolated that mother’s breast milk microbiome also has epigenetic influences and it is associated with infants’ brain function and neurodevelopment. The precise mechanisms through which the breast milk microbiome carries out such effects on infants’ brains remain to be elucidated.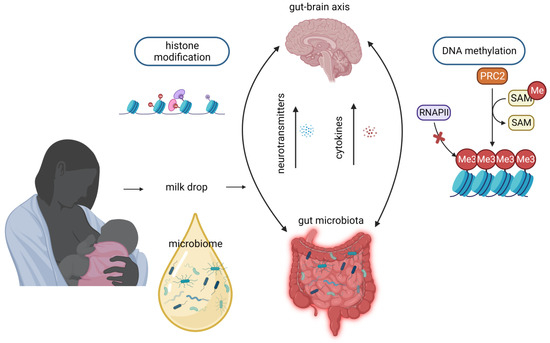
References
- GBD 2015 Obesity Collaborators; Afshin, A.; Forouzanfar, M.H.; Reitsma, M.B.; Sur, P.; Estep, K.; Lee, A.; Marczak, L.; Mokdad, A.H.; Moradi-Lakeh, M.; et al. Health Effects of Overweight and Obesity in 195 Countries over 25 Years. N. Engl. J. Med. 2017, 377, 13–27.
- Wilson, J. The Barker hypothesis. An analysis. Aust. N. Z. J. Obstet. Gynaecol. 1999, 39, 1–7.
- Calkins, K.; Devaskar, S.U. Fetal origins of adult disease. Curr. Probl. Pediatr. Adolesc. Health Care 2011, 41, 158–176.
- Barker, D.J.; Osmond, C.; Kajantie, E.; Eriksson, J.G. Growth and chronic disease: Findings in the Helsinki Birth Cohort. Ann. Hum. Biol. 2009, 36, 445–458.
- Barker, D.J. The developmental origins of adult disease. J. Am. Coll. Nutr. 2004, 23, 588S–595S.
- Goyal, D.; Limesand, S.W.; Goyal, R. Epigenetic responses and the developmental origins of health and disease. J. Endocrinol. 2019, 242, T105–T119.
- Holme, A.M.; Sitras, V. Developmental origin of health and disease—Evidence and time for action. Acta Obstet. Gynecol. Scand. 2020, 99, 961–962.
- Heindel, J.J.; Balbus, J.; Birnbaum, L.; Brune-Drisse, M.N.; Grandjean, P.; Gray, K.; Landrigan, P.J.; Sly, P.D.; Suk, W.A.; Cory Slechta, D.; et al. Developmental Origins of Health and Disease: Integrating Environmental Influences. Endocrinology 2015, 156, 3416–3421.
- Petronis, A. Epigenetics as a unifying principle in the aetiology of complex traits and diseases. Nature 2010, 465, 721–727.
- Deichmann, U. Epigenetics: The origins and evolution of a fashionable topic. Dev. Biol. 2016, 416, 249–254.
- Chen, M.; Zhang, L. Epigenetic mechanisms in developmental programming of adult disease. Drug Discov. Today 2011, 16, 1007–1018.
- Banik, A.; Kandilya, D.; Ramya, S.; Stünkel, W.; Chong, Y.S.; Dheen, S.T. Maternal Factors that Induce Epigenetic Changes Contribute to Neurological Disorders in Offspring. Genes 2017, 8, 150.
- Barker, D.J.P. Sir Richard Doll Lecture. Developmental origins of chronic disease. Public Health 2012, 126, 185–189.
- Plagemann, A. Maternal diabetes and perinatal programming. Early Hum. Dev. 2011, 87, 743–747.
- Horta, B.L.; de Lima, N.P. Breastfeeding and Type 2 Diabetes: Systematic Review and Meta-Analysis. Curr. Diab. Rep. 2019, 19, 1.
- Camacho-Morales, A.; Caba, M.; García-Juárez, M.; Caba-Flores, M.D.; Viveros-Contreras, R.; Martínez-Valenzuela, C. Breastfeeding Contributes to Physiological Immune Programming in the Newborn. Front. Pediatr. 2021, 9, 744104.
- Ozkan, H.; Tuzun, F.; Taheri, S.; Korhan, P.; Akokay, P.; Yılmaz, O.; Duman, N.; Özer, E.; Tufan, E.; Kumral, A.; et al. Epigenetic Programming Through Breast Milk and Its Impact on Milk-Siblings Mating. Front. Genet. 2020, 11, 569232.
- Chutipongtanate, S.; Morrow, A.L.; Newburg, D.S. Human Milk Extracellular Vesicles: A Biological System with Clinical Implications. Cells 2022, 11, 2345.
- Tingö, L.; Ahlberg, E.; Johansson, L.; Pedersen, S.A.; Chawla, K.; Sætrom, P.; Cione, E.; Simpson, M.R. Non-Coding RNAs in Human Breast Milk: A Systematic Review. Front. Immunol. 2021, 12, 725323.
- Melnik, B.C.; Stremmel, W.; Weiskirchen, R.; John, S.M.; Schmitz, G. Exosome-Derived MicroRNAs of Human Milk and Their Effects on Infant Health and Development. Biomolecules 2021, 11, 851.
- Koh, K. Maternal breastfeeding and children’s cognitive development. Soc. Sci. Med. 2017, 187, 101–108.
- Wallenborn, J.T.; Levine, G.A.; Santos, A.C.D.; Grisi, S.; Brentani, A.; Fink, G. Breastfeeding, Physical Growth, and Cognitive Development. Pediatrics 2021, 147, e2020008029.
- Lewallen, L.P. Breastfeeding is important for cognitive development in term and preterm infants. Evid. Based Nurs. 2012, 15, 85–86.
- Horta, B.L.; de Mola, C.L.; Victora, C.G. Breastfeeding and intelligence: A systematic review and meta-analysis. Acta Paediatr. 2015, 104, 14–19.
- Kramer, M.S.; Aboud, F.; Mironova, E.; Vanilovich, I.; Platt, R.W.; Matush, L.; Igumnov, S.; Fombonne, E.; Bogdanovich, N.; Ducruet, T.; et al. Breastfeeding and child cognitive development: New evidence from a large randomized trial. Arch. Gen. Psychiatry 2008, 65, 578–584.
- Chetta, K.E.; Schulz, E.V.; Wagner, C.L. Outcomes improved with human milk intake in preterm and full-term infants. Semin. Perinatol. 2021, 45, 151384.
- Sullivan, G.; Vaher, K.; Blesa, M.; Galdi, P.; Stoye, D.Q.; Quigley, A.J.; Thrippleton, M.J.; Norrie, J.; Bastin, M.E.; Boardman, J.P. Breast Milk Exposure is Associated with Cortical Maturation in Preterm Infants. Ann. Neurol. 2022, 93, 591–603.
- Xu, J.; Shin, J.; McGee, M.; Unger, S.; Bando, N.; Sato, J.; Vandewouw, M.; Patel, Y.; Branson, H.M.; Paus, T.; et al. Intake of mother’s milk by very-low-birth-weight infants and variation in DNA methylation of genes involved in neurodevelopment at 5.5 years of age. Am. J. Clin. Nutr. 2022, 116, 1038–1048.
- Hu, Y.; Thaler, J.; Nieuwland, R. Extracellular Vesicles in Human Milk. Pharmaceuticals 2021, 14, 1050.
- He, S.; Liu, G.; Zhu, X. Human breast milk-derived exosomes may help maintain intestinal epithelial barrier integrity. Pediatr. Res. 2021, 90, 366–372.
- Shandilya, S.; Rani, P.; Onteru, S.K.; Singh, D. Small Interfering RNA in Milk Exosomes Is Resistant to Digestion and Crosses the Intestinal Barrier In Vitro. J. Agric. Food Chem. 2017, 65, 9506–9513.
- Melnik, B.C.; John, S.M.; Schmitz, G. Milk is not just food but most likely a genetic transfection system activating mTORC1 signaling for postnatal growth. Nutr. J. 2013, 12, 103.
- Chen, C.C.; Liu, L.; Ma, F.; Wong, C.W.; Guo, X.E.; Chacko, J.V.; Farhoodi, H.P.; Zhang, S.X.; Zimak, J.; Ségaliny, A.; et al. Elucidation of Exosome Migration across the Blood–Brain Barrier Model In Vitro. Cell. Mol. Bioeng. 2016, 9, 509–529.
- Green, D.; Dalmay, T.; Chapman, T. Microguards and micromessengers of the genome. Heredity 2016, 116, 125–134.
- Ying, S.-Y.; Chang, D.C.; Lin, S.-L. The microRNA (miRNA): Overview of the RNA genes that modulate gene function. Mol. Biotechnol. 2008, 38, 257–268.
- Carrillo-Lozano, E.; Sebastián-Valles, F.; Knott-Torcal, C. Circulating microRNAs in Breast Milk and Their Potential Impact on the Infant. Nutrients 2020, 12, 3066.
- Bartel, D.P. Metazoan MicroRNAs. Cell 2018, 173, 20–51.
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297.
- Wang, Z. MicroRNA: A matter of life or death. World J. Biol. Chem. 2010, 1, 41–54.
- Hatmal, M.M.; Al-Hatamleh, M.A.I.; Olaimat, A.N.; Alshaer, W.; Hasan, H.; Albakri, K.A.; Alkhafaji, E.; Issa, N.N.; Al-Holy, M.A.; Abderrahman, S.M.; et al. Immunomodulatory Properties of Human Breast Milk: MicroRNA Contents and Potential Epigenetic Effects. Biomedicines 2022, 10, 1219.
- Aliperti, V.; Skonieczna, J.; Cerase, A. Long Non-Coding RNA (lncRNA) Roles in Cell Biology, Neurodevelopment and Neurological Disorders. Non Coding RNA 2021, 7, 36.
- Ng, S.Y.; Lin, L.; Soh, B.S.; Stanton, L.W. Long noncoding RNAs in development and disease of the central nervous system. Trends Genet. 2013, 29, 461–468.
- Shi, C.; Zhang, L.; Qin, C. Long non-coding RNAs in brain development, synaptic biology, and Alzheimer’s disease. Brain Res. Bull. 2017, 132, 160–169.
- Lipovich, L.; Tarca, A.L.; Cai, J.; Jia, H.; Chugani, H.T.; Sterner, K.N.; Grossman, L.I.; Uddin, M.; Hof, P.R.; Sherwood, C.C.; et al. Developmental changes in the transcriptome of human cerebral cortex tissue: Long noncoding RNA transcripts. Cereb. Cortex 2014, 24, 1451–1459.
- Li, L.; Zhuang, Y.; Zhao, X.; Li, X. Long Non-coding RNA in Neuronal Development and Neurological Disorders. Front. Genet. 2018, 9, 744.
- Wilkinson, B.; Campbell, D.B. Contribution of long noncoding RNAs to autism spectrum disorder risk. Int. Rev. Neurobiol. 2013, 113, 35–59.
- Weissman, I.L. Translating stem and progenitor cell biology to the clinic: Barriers and opportunities. Science 2000, 287, 1442–1446.
- Kersin, S.G.; Ozek, E. Breast milk stem cells: Are they magic bullets in neonatology? Turk. Arch. Pediatr. 2021, 56, 187–191.
- Briere, C.E.; McGrath, J.M.; Jensen, T.; Matson, A.; Finck, C. Breast Milk Stem Cells: Current Science and Implications for Preterm Infants. Adv. Neonatal. Care 2016, 16, 410–419.
- Cregan, M.D.; Fan, Y.; Appelbee, A.; Brown, M.L.; Klopcic, B.; Koppen, J.; Mitoulas, L.R.; Piper, K.M.E.; Choolani, M.A.; Chong, Y.-S.; et al. Identification of nestin-positive putative mammary stem cells in human breastmilk. Cell Tissue Res. 2007, 329, 129–136.
- Briere, C.E.; Jensen, T.; McGrath, J.M.; Young, E.E.; Finck, C. Stem-Like Cell Characteristics from Breast Milk of Mothers with Preterm Infants as Compared to Mothers with Term Infants. Breastfeed. Med. 2017, 12, 174–179.
- Aydın, M.Ş.; Yiğit, E.N.; Vatandaşlar, E.; Erdoğan, E.; Öztürk, G. Transfer and Integration of Breast Milk Stem Cells to the Brain of Suckling Pups. Sci. Rep. 2018, 8, 14289.
- Belkozhayev, A.M.; Al-Yozbaki, M.; George, A.; Ye Niyazova, R.; Sharipov, K.O.; Byrne, L.J.; Wilson, C.M. Extracellular Vesicles, Stem Cells and the Role of miRNAs in Neurodegeneration. Curr. Neuropharmacol. 2022, 20, 1450–1478.
- Davis, C.; Savitz, S.I.; Satani, N. Mesenchymal Stem Cell Derived Extracellular Vesicles for Repairing the Neurovascular Unit after Ischemic Stroke. Cells 2021, 10, 767.
- Chang, Y.-H.; Wu, K.-C.; Harn, H.-J.; Lin, S.-Z.; Ding, D.-C. Exosomes and Stem Cells in Degenerative Disease Diagnosis and Therapy. Cell Transplant. 2018, 27, 349–363.
- Lendahl, U.; Zimmerman, L.B.; McKay, R.D.G. CNS stem cells express a new class of intermediate filament protein. Cell 1990, 60, 585–595.
- Hosseini, S.M.; Talaei-Khozani, T.; Sani, M.; Owrangi, B. Differentiation of human breast-milk stem cells to neural stem cells and neurons. Neurol. Res. Int. 2014, 2014, 807896.
- Turnbaugh, P.J.; Ley, R.E.; Hamady, M.; Fraser-Liggett, C.M.; Knight, R.; Gordon, J.I. The human microbiome project. Nature 2007, 449, 804–810.
- Berg, G.; Rybakova, D.; Fischer, D.; Cernava, T.; Vergès, M.-C.C.; Charles, T.; Chen, X.; Cocolin, L.; Eversole, K.; Corral, G.H.; et al. Microbiome definition re-visited: Old concepts and new challenges. Microbiome 2020, 8, 103.
- Gavin, A.; Ostovar, K.; Gavin, A.; Ostovar, K. Microbiological Characterization of Human Milk (1). J. Food Prot. 1977, 40, 614–616.
- Jones, C.A. Maternal transmission of infectious pathogens in breast milk. J. Paediatr. Child Health 2001, 37, 576–582.
- Martín, R.; Langa, S.; Reviriego, C.; Jimínez, E.; Marín, M.L.; Xaus, J.; Fernández, L.; Rodríguez, J.M. Human milk is a source of lactic acid bacteria for the infant gut. J. Pediatr. 2003, 143, 754–758.
- Collado, M.; Delgado, S.; Maldonado, A.; Rodríguez, J. Assessment of the bacterial diversity of breast milk of healthy women by quantitative real-time PCR. Lett. Appl. Microbiol. 2009, 48, 523–528.
- Hunt, K.M.; Foster, J.A.; Forney, L.J.; Schütte, U.M.E.; Beck, D.L.; Abdo, Z.; Fox, L.K.; Williams, J.E.; McGuire, M.K.; McGuire, M.A. Characterization of the diversity and temporal stability of bacterial communities in human milk. PLoS ONE 2011, 6, e21313.
- Yi, D.Y.; Kim, S.Y. Human Breast Milk Composition and Function in Human Health: From Nutritional Components to Microbiome and MicroRNAs. Nutrients 2021, 13, 3094.
- Moossavi, S.; Sepehri, S.; Robertson, B.; Bode, L.; Goruk, S.; Field, C.J.; Lix, L.M.; de Souza, R.J.; Becker, A.B.; Mandhane, P.J.; et al. Composition and Variation of the Human Milk Microbiota Are Influenced by Maternal and Early-Life Factors. Cell Host Microbe 2019, 25, 324–335.e4.
- Zimmermann, P.; Curtis, N. Breast milk microbiota: A review of the factors that influence composition. J. Infect. 2020, 81, 17–47.
- Damaceno, Q.S.; Souza, J.P.; Nicoli, J.R.; Paula, R.L.; Assis, G.B.; Figueiredo, H.C.; Azevedo, V.; Martins, F.S. Evaluation of Potential Probiotics Isolated from Human Milk and Colostrum. Probiotics Antimicrob. Proteins 2017, 9, 371–379.
- Solís, G.; de los Reyes-Gavilan, C.G.; Fernández, N.; Margolles, A.; Gueimonde, M. Establishment and development of lactic acid bacteria and bifidobacteria microbiota in breast-milk and the infant gut. Anaerobe 2010, 16, 307–310.
- Khodayar-Pardo, P.; Mira-Pascual, L.; Collado, M.C.; Martínez-Costa, C. Impact of lactation stage, gestational age and mode of delivery on breast milk microbiota. J. Perinatol. 2014, 34, 599–605.
- Dahaban, N.M.; Romli, M.F.; Roslan, N.R.; Kong, S.S.-S.; Cheah, F.-C. Bacteria in expressed breastmilk from mothers of premature infants and maternal hygienic status. Breastfeed. Med. 2013, 8, 422–423.
- Boix-Amorós, A.; Collado, M.C.; Mira, A. Relationship between Milk Microbiota, Bacterial Load, Macronutrients, and Human Cells during Lactation. Front. Microbiol. 2016, 7, 492.
- Hermansson, H.; Kumar, H.; Collado, M.C.; Salminen, S.; Isolauri, E.; Rautava, S. Breast Milk Microbiota Is Shaped by Mode of Delivery and Intrapartum Antibiotic Exposure. Front. Nutr. 2019, 6, 4.
- Simpson, M.R.; Avershina, E.; Storrø, O.; Johnsen, R.; Rudi, K.; Øien, T. Breastfeeding-associated microbiota in human milk following supplementation with Lactobacillus rhamnosus GG, Lactobacillus acidophilus La-5, and Bifidobacterium animalis ssp. lactis Bb-12. J. Dairy Sci. 2018, 101, 889–899.
- Gueimonde, M.; Laitinen, K.; Salminen, S.; Isolauri, E. Breast milk: A source of bifidobacteria for infant gut development and maturation? Neonatology 2007, 92, 64–66.
- Vuong, H.E. Intersections of the microbiome and early neurodevelopment. Int. Rev. Neurobiol. 2022, 167, 1–23.
- Lu, J.; Drobyshevsky, A.; Lu, L.; Yu, Y.; Caplan, M.S.; Claud, E.C. Microbiota from Preterm Infants Who Develop Necrotizing Enterocolitis Drives the Neurodevelopment Impairment in a Humanized Mouse Model. Microorganisms 2023, 11, 1131.
- Xia, J.; Claud, E.C. Gut Microbiome-Brain Axis as an Explanation for the Risk of Poor Neurodevelopment Out-come in Preterm Infants with Necrotizing Enterocolitis. Microorganisms 2023, 11, 1035.
- Indrio, F.; Martini, S.; Francavilla, R.; Corvaglia, L.; Cristofori, F.; Mastrolia, S.A.; Neu, J.; Rautava, S.; Spena, G.R.; Raimondi, F.; et al. Epigenetic Matters: The Link between Early Nutrition, Microbiome, and Long-term Health Development. Front. Pediatr. 2017, 5, 178.
- Tognini, P. Gut Microbiota: A Potential Regulator of Neurodevelopment. Front. Cell. Neurosci. 2017, 11, 25.
- Laue, H.E.; Coker, M.O.; Madan, J.C. The Developing Microbiome from Birth to 3 Years: The Gut-Brain Axis and Neurodevelopmental Outcomes. Front. Pediatr. 2022, 10, 815885.
- Ihekweazu, F.D.; Versalovic, J. Development of the Pediatric Gut Microbiome: Impact on Health and Disease. Am. J. Med Sci. 2018, 356, 413–423.
- Valicenti-McDermott, M.; McVicar, K.; Rapin, I.; Wershil, B.K.; Cohen, H.; Shinnar, S. Frequency of gastrointestinal symptoms in children with autistic spectrum disorders and association with family history of autoimmune disease. J. Dev. Behav. Pediatr. 2006, 27, S128–S136.
- Hsiao, E.Y.; McBride, S.W.; Hsien, S.; Sharon, G.; Hyde, E.R.; McCue, T.; Codelli, J.A.; Chow, J.; Reisman, S.E.; Petrosino, J.F.; et al. Microbiota modulate behavioral and physiological abnormalities associated with neurodevelopmental disorders. Cell 2013, 155, 1451–1463.
- Lu, J.; Claud, E.C. Connection between gut microbiome and brain development in preterm infants. Dev. Psychobiol. 2019, 61, 739–751.