Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Julio José Jiménez-Alonso and Version 3 by Lindsay Dong.
Cancer cells cannot proliferate and survive unless they obtain sufficient levels of the 20 proteinogenic amino acids (AAs). Unlike normal cells, cancer cells have genetic and metabolic alterations that may limit their capacity to obtain adequate levels of the 20 AAs in challenging metabolic environments. However, since normal diets provide all AAs at relatively constant levels and ratios, these potentially lethal genetic and metabolic defects are eventually harmless to cancer cells. If the normal diet of cancer patients is temporarily replaced with artificial diets in which the levels of specific AAs are manipulated, cancer cells may be unable to proliferate and survive.
- cancer metabolism
- anticancer activity
- artificial diets
1. Introduction
The first metabolic alteration of cancer cells was discovered almost one century ago by the German biochemist Otto Warburg. He observed that, unlike normal cells, cancer cells convert high amounts of glucose into lactate in the presence of normal oxygen levels [1]. This alteration in glucose metabolism, known as aerobic glycolysis or the Warburg effect, is now widely used in diagnostic imaging to trace cancers and evaluate cancer treatment responses [2][3][2,3]. The clinical use of FDG-based PET imaging has continually shown that most primary and metastatic cancers have a significant increase in glucose uptake compared to normal tissues [2][3][2,3].
Over many decades, the Warburg effect was considered to be an irrelevant oddity of cancer cells, probably because it was unknown why cancer cells used this primitive form of energy production when the availability of oxygen allows for a much more effective way of producing energy: oxidative phosphorylation. The explanation of the Warburg effect is simple when one realizes that glycolysis not only serves to produce energy, but also to produce building blocks to generate new cells [4]. Cancer cells have high glycolytic rates because the breakdown of glucose molecules generates the building blocks needed to produce many cellular components for the new cancer cells created during cell division. One cell cannot divide to produce two cells unless glucose is broken down into these building blocks. Since both glucose and oxygen are supplied together through the blood, cancer cells have no choice but to activate glycolysis in the presence of oxygen in order to proliferate. Since oxygen inhibits glycolysis (Pasteur Effect) indirectly via ATP generation, cancer cells partially uncouple oxygen utilization from ATP production to activate glycolysis in the presence of normal oxygen levels. By deviating oxygen metabolism from the route that generates ATP to the route that produces reactive oxygen species (ROS), cancer cells manage to keep sustained glycolytic rates under aerobic conditions [4][5][6][4,5,6]. The subsequent increased production of ROS, such as superoxide anion and hydrogen peroxide, leads to a state of increased basal oxidative stress, which represents another metabolic hallmark of cancer cells [5][7][8][9][10][5,7,8,9,10].
Targeting the Warburg effect for cancer therapy is difficult because cells from different normal tissues also need glucose for their survival and proliferation. However, understanding the Warburg effect is important to realize that the genetic alterations in cancer cells are insufficient for cancer cell proliferation and survival. Cancer cells also need to take glucose and other nutrients, such as amino acids (AAs), from the extracellular environment to proliferate and survive under conditions of elevated oxidative stress. Although the metabolic changes in cancer cells play an important role in carcinogenesis and cancer progression, these changes can also be exploited to develop new cancer therapies [10][11][12][13][14][15][10,11,12,13,14,15].
2. Protein Restriction
Dietary protein restriction can increase life expectancy [16][28] and reduce the incidence of age-related diseases such as cancer [17][29]. It is well known that proliferating cancer cells must produce new proteins for the new cells created during cell division. Since dietary proteins provide the AAs needed to generate the proteins of the new cancer cells, it is not surprising that low-protein diets can restrict tumor growth in animal models [18][30]. A reduction in IGF-1 levels has been proposed as a key mechanism by which low-protein diets induce anticancer activity. Murine models of melanoma and breast cancer have revealed that mice fed with a low-protein diet (4% kcal protein) had reduced IFG-1 levels and a reduced tumor progression compared to those fed with a high-protein diet (18% kcal protein) [19][31]. Weight loss was observed in elderly mice but not in young mice. Similarly, a low-protein diet reduced the IGF-1 levels in patients aged 50–65 years and reduced the risk of cancer death, while a low-protein diet increased the mortality among elderly patients (+65 years old) [19][31]. This suggests that low-protein diets might induce anticancer activity in middle-aged adults but not in elderly patients. Low-protein diets are not active in all cancer types. Enhanced cancer immunosurveillance is another possible mechanism by which low-protein diets induce anticancer activity. A reduction in dietary proteins (17–15% protein vs. 19% protein) induced IRE1α-dependent endoplasmic reticulum (ER) stress in cancer cells, which resulted in cytokine production and improved the anticancer immune response [20][33]. A lower protein intake (12%) reversed this anticancer effect, which suggested that a certain level of protein intake was needed for activity [20][33]. However, other studies have shown that diets with a lower protein intake (7% protein vs. 21% protein) inhibited cancer progression and induced a synergistic effect when combined to anti-PD-1 immunotherapy [21][34]. Low-protein diets also induced anticancer activity in immunosuppressed mice, therefore suggesting that the anticancer activity of protein restriction is not necessarily mediated by the immune system [22][35]. The type of protein can also modulate this cancer progression. Mice fed with a 20% plant-based protein diet showed a reduced tumor growth in xenograft and syngeneic cancer models when compared to mice fed with a 20% animal-based protein diet [22][23][35,36]. The anticancer activity of diets based on plant proteins was explained by declines in the plasma levels of IFG-1 and insulin, which decreased the activity of the IGF/AKT/mTOR pathway and led to epigenetic modifications that restricted tumor growth [22][23][35,36].3. Essential Amino Acids
3.1. Leucine
Leucine (Leu) is one of the nine EAAs for humans; this means that we cannot biosynthesize it from other nutrients and we must take it from the diet. Like all 20 proteinogenic AAs, Leu is necessary for protein synthesis. Leu is also important for other cellular functions. For example, Leu is a key intracellular sensor of AAs under starvation conditions and it regulates protein turnover through mTORC1 signaling [24][37]. Like isoleucine (Ile) and valine (Val), Leu is a branched-chain amino acid (BCAA); these AAs can regulate the lipid metabolism in cancer cells by providing carbon skeletons for fatty-acid biosynthesis [25][38]. The dietary restriction of Leu can induce in vivo anticancer effects. In 1956, Sugimura et al. [26][39] found that dietary deprivation of Leu for 5 days reduced the growth rate of Walker tumors in rats by 24%; however, it also induced body weight loss. In 1971, reducing the dietary levels of Leu from 0.8% to 0.1% for 3 weeks significantly reduced tumor growth in mice with breast adenocarcinomas; the tumor weights were 32 ± 10 g in mice fed with a standard AA-based diet (0.80% Leu), 38 ± 4 g for those on a 0.50% Leu diet, 32 ± 4 g for those on a 0.25% Leu diet, and 16 ± 6 g for those on a 0.10% Leu diet [27][40]. Body weight loss was observed in the mice fed with the 0.10% Leu diet [27][40]. Mechanistically, Leu limitation restricts protein synthesis, cell division, and tumor growth. In addition, Leu restriction can reduce Leu catabolism and limit the fatty acid biosynthesis and lipogenesis in cancer cells. BCAAs catabolism plays an important role in pancreatic cancer growth by regulating lipogenesis [25][38]. BCAT2 and BCKDHA knockdown impaired pancreatic cancer cell proliferation in vitro and in vivo by inhibiting fatty acid synthesis [25][38]. Since Leu restriction can reduce tumor growth, it makes sense to think that Leu supplementation may facilitate cancer progression. A study showed that a 5% Leu supplementation increased cancer growth in a syngeneic model of pancreatic cancer [28][49]. Maintaining high Leu levels may be important for preventing proteolysis, which could be beneficial in certain circumstances. Leu is a critical intracellular sensor of AAs under starvation conditions. This AA activates mTORC1 signaling and inhibits autophagy and proteasome-mediated proteolysis. Supplementing Leu may therefore prevent intracellular and extracellular proteolysis [29][30][31][32][50,51,52,53]. If muscle and liver proteolysis is not prevented, the lysis of proteins in these organs would supply any AA restricted in the diet [33][34][35][26,27,54]. The inhibition of proteolysis is also important in avoiding weight loss and cachexia. Cachexia is a syndrome of progressive body weight loss with reductions in skeletal muscle and fat mass [36][55]. Ultimately, cachexia reduces the tolerability of anticancer treatments and leads to a reduced life expectancy and quality of life [36][37][55,56]. Leu supplementation can alleviate cancer cachexia by activating mTORC1 and decreasing protein degradation [30][36][38][39][51,55,57,58].3.2. Isoleucine
Like all proteinogenic AAs, the BCAA Ile is necessary for protein synthesis. Ile also participates in other biological processes, including lipogenesis and immune function regulation [24][25][40][37,38,69]. Experiments conducted several decades ago revealed that a complete dietary Ile restriction for 5 days inhibited tumor growth by 40% in Walker tumor-bearing rats; however, this force-fed intervention caused the animals to lose 1–2 g per day [26][39]. Dietary Ile restriction (from 0.5% to 0.05%) also resulted in tumor growth inhibition in C57BL/6 mice with BW10232 mammary carcinomas [27][40].3.3. Valine
Like Leu and Ile, Val is an essential and proteinogenic BCAA. It is also involved in other cellular functions, including the regulation of lipid and glucose metabolism [24][25][41][37,38,70]. A dietary depletion of Val for 5 days reduced tumor growth by 41% in Walker tumor-bearing rats [26][39]. However, all the animals on this Val-free diet rapidly sickened and failed to survive beyond 9 days on this diet. A dietary limitation of Val (from 0.7% to 0.1%) significantly decreased tumor growth in mice with breast adenocarcinomas, but also induced body weight loss [27][40].3.4. Threonine
Threonine (Thr) is an essential and proteinogenic AA. Like other AAs, Thr catabolism can also provide amino groups for the synthesis of NEAAs and carbon skeletons for biosynthesis and energy production [24][37]. The force feeding of a diet lacking in Thr for 5 days reduced tumor growth by 28% in Walker tumor-bearing rats [26][39]. This diet caused the animals to lose between 0.2 and 1.0 g/day over an 11-day period [26][39].3.5. Lysine
Lys is an essential and proteinogenic AA, whose deficiency can trigger severe malnutrition [24][42][37,71]. Lys is also used for carnitine production and participates in protein methylation, acetylation, ubiquitination, and glycosylation [24][37]. The anticancer activity of Lys restriction was evaluated 80 years ago in mice with spontaneous breast cancer [43][72]. The author of this research first devised a Lys-deficient diet suitable for human consumption (palatable, adequate in calories, minerals, and vitamins, and sufficient for keeping nitrogen balance). After observing in two healthy humans that nitrogen equilibrium could be maintained with this diet, he obtained and reproduced a strain of mice characterized by a high incidence of spontaneous mammary carcinomas. The mice that developed tumors were fed with the Lys-deficient diet. The diet inhibited the growth rate of the tumors, but also the rate of normal growth in the mice. These inhibitory effects were abolished upon the addition of Lys, therefore indicating that Lys was essential for both normal and malignant growth. When the Lys-deficient diet was fed to them for several weeks, the antitumor effect wore off and the tumors resumed rapid growth.3.6. Phenylalanine
Phenylalanine (Phe) is an essential and proteinogenic AA with an aromatic group in its structure. Phe can be used to synthesize tyrosine (Tyr), a proteinogenic NEAA that produces important molecules such as catecholamines (dopamine, epinephrine, and norepinephrine) and melanin [24][37]. Dietary Phe limitation is used in people with phenylketonuria, an inborn disease caused by the inactivity of the enzyme phenylalanine hydroxylase, which converts Phe into Tyr; the accumulation of Phe can lead to seizures and intellectual disability [44][73].3.7. Histidine
Histidine (His) is an aromatic EAA required for protein synthesis. This AA is involved in other cellular functions, including the synthesis of histamine and carnitine [24][45][37,92]. The force feeding of a diet lacking in His for 5 days reduced tumor growth by 19% in the Walker rat model [26][39]. More recently, the dietary limitation of His was found to selectively limit the growth of MYC-dependent neural tumors in a Drosophila model [46][93].3.8. Tryptophan
Although tryptophan (Trp) is the least abundant EAA in the diet, it is necessary for protein synthesis and the production of a variety of biologically active compounds, including serotonin, melatonin, and niacin (a component of NAD and NADP) [24][47][37,95]. In addition, Trp and its catabolic derivatives modulate the immune function and play key roles in autoimmune diseases and antitumor immunity [47][48][95,96]. In 1959, a total Trp restriction for 5 days inhibited tumor growth by 19% in Walker tumor-bearing rats, with moderate weight loss in the animals [26][39]. In 1971, a Trp limitation (from 0.10% to 0.02%) reduced tumor growth in C57BL/6 mice with BW10232 mammary carcinomas [27][40]. The tumor weights were 33 ± 6 g in mice fed with a standard AA-based diet (0.15% Trp), 33 ± 13 g for those on a 0.10% Trp diet, 31 ± 10 g for those on a 0.05% Trp diet, and 16 ± 8 g for those on a 0.02% Trp diet [27][40]. The mice fed with the 0.02% Trp diet lost 28% of their weight in 3 weeks [27][40]. A moderate dietary limitation of Trp (0.05%) did not show anticancer activity in C3H mice bearing mammary adenocarcinomas [49][97].3.9. Methionine
Methionine (Met) is an essential and proteinogenic AA that contains a sulfur atom in its structure. Met is the precursor of S-adenosyl methionine (SAM), which is a methyl donor involved in DNA methylation and epigenetics. Met also produces Cys through the irreversible transsulfuration pathway, which, in turn, produces several sulfur-containing molecules with important cellular roles, including glutathione (GSH), hydrogen sulfide (H2S), and taurine (Tau) [24][50][51][52][24,37,103,104]. Dietary Met restriction has shown anticancer activity in numerous preclinical studies [21][26][27][33][34][53][54][55][56][57][58][59][60][61][62][63][64][65][66][67][68][69][70][71][72][73][26,27,34,39,40,105,106,107,108,109,110,111,112,113,114,115,116,117,118,119,120,121,122,123,124,125]. A dietary Met depletion (0%) induced anticancer activity in rats [26][69][70][71][72][39,121,122,123,124] and mice [53][54][55][56][57][58][59][73][105,106,107,108,109,110,111,125] with different types of cancer. The force feeding of a diet lacking in Met for 5 days reduced tumor growth by 39% in the Walker rat model, but this diet caused the animals to lose l–2 g weight/day [26][39]. Met restriction (from 0.60% to 0.10%) reduced tumor growth in C57BL mice with BW10232 mammary carcinomas [27][40]. The tumor weights were 36 ± 13 g in mice fed with a standard AA-based diet (0.90% Met; 0.2% Cys), 33 ± 11 g for those on a 0.60% Met diet, 30 ± 6 g for those on a 0.40% Met diet, 29 ± 7 g for those on a 0.20% Met diet, and 16 ± 7 g for those on a 0.10% Met diet [27][40]. The mice fed with the 0.10% Met diet lost 10% of their initial weight in 3 weeks [27][40].4. Non-Essential Amino Acids
4.1. Cysteine
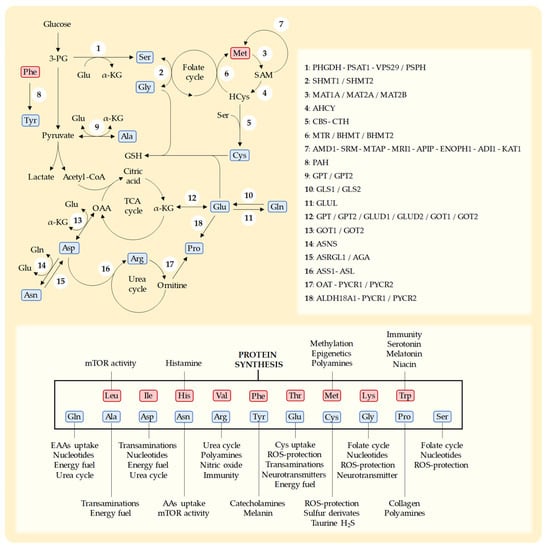
Cys is a sulfur-containing NEAA with multiple cellular roles [74][169]. Humans can biosynthesize Cys from the EAA Met through the transsulfuration pathway [50][74][75][24,169,170] (Figure 12). In addition to being necessary for protein synthesis, Cys is essential for the production of a variety of sulfur-containing molecules with important biological roles [75][170]. These include the iron–sulfur clusters found in enzymes of the electron transport chain (ETC) [76][171], coenzyme A, and thioredoxins [77][172]. Cys also produces taurine [24][37] and hydrogen sulfide (H2S) [24][37] and is the rate-limiting AA for the production of the tripeptide GSH (Glu-Cys-Gly) [74][75][169,170]. GSH is essential for protecting cells against the toxic effects ROS [74][169].Figure 12. Schematic representation of key metabolic pathways for the biosynthesis of NEAAs, the enzymes in each pathway, and the main functions of each AA [35][78][54,173]. NEAAs are represented in blue and EAAs in red. Leucine (Leu), isoleucine (Ile), histidine (His), valine (Val), phenylalanine (Phe), threonine (Thr), methionine (Met), lysine (Lys), tryptophan (Trp), glutamine (Gln), alanine (Ala), aspartate (Asp), asparagine (Asn), arginine (Arg), tyrosine (Tyr), glutamate (Glu), cysteine (Cys), glycine (Gly), proline (Pro), and serine (Ser). 3-phospho-D-glycerate (3-PG), S-adenosylmethionine (SAM), homocysteine (HCys), glutathione (GSH), α-ketoglutarate (α-KGlu), tricarboxylic acid cycle (TCA), oxaloacetate (OAA), reactive oxygen species (ROS). D-3-phosphoglycerate dehydrogenase (PHGDH), phosphoserine aminotransferase-1 (PSAT1), vacuolar protein sorting-associated protein-29 (VPS29), phosphoserine phosphatase (PSPH), serine hydroxymethyltransferase-1 (SHMT1), serine hydroxymethyltransferase-2 (SHMT2), S-adenosylmethionine synthase isoform type-1 (MAT1A), S-adenosylmethionine synthetase isoform type-2 (MAT2A), methionine adenosyltransferase 2 subunit beta (MAT2B), adenosylhomocysteinase (AHCY), cystathionine β-synthase (CBS), cystathionine γ-lyase (CTH), methionine synthase (MTR), betaine-homocysteine methyltransferase (BHMT), betaine-homocysteine methyltransferase-2 (BHMT2), S-adenosylmethionine decarboxylase (AMD1), spermidine synthase (SRM), 5′-methylthioadenosine phosphorylase (MTAP), methylthioribose-1-phosphate isomerase (MRI1), methylthioribulose 1-phosphate dehydratase (APIP), enolase-phosphatase (ENOPH1), 1,2-dihydroxy-3-keto-5-methylthiopentene dioxygenase (ADI1), 2-oxo-4-methylthiobutanoate aminotransferase (KYAT1), phenylalanine hydroxylase (PAH), alanine aminotransferase-1 (GPT), alanine aminotransferase-2 (GPT2), glutaminase-1 (GLS1), glutaminase-2 (GLS2), glutamine synthetase (GLUL), glutamate dehydrogenase-1 (GLUD1), glutamate dehydrogenase-2 (GLUD2), aspartate aminotransferase-1 (GOT1), aspartate aminotransferase-2 (GOT2), asparagine synthetase (ASNS), asparaginase (ASRGL1), aspartylglucosaminidase (AGA), argininosuccinate synthase (ASS1), argininosuccinate lyase (ASL), ornithine aminotransferase (OAT), pyrroline-5-carboxylate reductase-1 (PYCR1), pyrroline-5-carboxylate reductase-2 (PYCR2), and δ-1-pyrroline-5-carboxylate synthase (ALDH18A1). Enzymes that participate in consecutive steps in a metabolic pathway are separated by “-” and enzymes that catalyze the same step in a metabolic pathway are separated by “/”.
The importance of Cys in tumor growth was first reported in 1936 [79][133]. In this study, Voegtlin et al. observed that a diet deficient in Cys/Met reduced tumor growth in mice with spontaneous breast cancer, and the addition of Cys abruptly stimulated tumor growth [79][133]. Since Met can produce Cys, Met has usually been restricted in many studies evaluating the anticancer activity of Cys depletion/restriction [21][27][33][34][59][61][68][69][70][79][26,27,34,40,111,113,120,121,122,133]. Intravenous parenteral nutrition with double Cys/Met restriction showed anticancer activity in rats with sarcoma [69][70][121,122] and inhibited the cancer proliferation in mice xenografted with human glioma cells [59][111].
Mechanistically, Cys restriction may induce anticancer activity by reducing the capacity of cancer cells to eliminate ROS. Cancer cells produce high levels of ROS, which may accumulate and produce cell death [8]. Cancer cells rely on GSH to reduce these ROS levels [50][24]. A dietary Cys restriction can decrease Cys plasma levels [80][176], reduce GSH biosynthesis [80][81][175,176], and increase the ROS levels in cancer cells [80][81][82][174,175,176].
Since Cys is necessary for immune cells, Cys restriction may reduce the ability of the immune system to eliminate cancer cells. Cys is essential for T-cell activation and function [83][177]. High CysS plasma levels have been associated with a higher probability of response to immune checkpoint inhibitors in patients with lung cancer [84][85][178,179]. However, the negative effect of Cys restriction on the immune antitumor response is controversial, because other studies have suggested that Cys restriction can increase the antitumor immune response [21][86][34,180].
Pharmacological approaches based on an enzymatic depletion of Cys and inhibition of Cys transporters support the idea that Cys restriction has potential for cancer therapy. These pharmacological interventions have been useful for understanding the possible mechanisms by which Cys restriction induces in vivo anticancer effects. In 2017, an optimized human cyst(e)inase enzyme was able to reduce the Cys and CysS plasma levels in mice and primates without causing toxicity [87][181]. Cyst(e)inase has shown anticancer activity in mouse models of a variety of cancers, including prostate, breast, chronic lymphocytic leukemia, pancreas, lung, renal, melanoma, and ovarian cancer [86][87][88][89][90][91][92][180,181,182,183,184,185,186]. Cyst(e)inase administration increases ROS levels, depletes the intracellular levels of GSH, and triggers ferroptosis in cancer cells [87][88][89][90][91][92][181,182,183,184,185,186].