Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 2 by Dean Liu and Version 1 by Jong-Wook Chung.
Genome editing has emerged as a powerful tool for accelerating crop improvement in horticultural crops by enabling precise modifications to their genetic makeup.
- CRISPR-Cas9
- crop improvement
- genome editing
- horticultural crops
1. Introduction to Genome Editing in Agriculture
Genome editing is a powerful biotechnological tool with transformative potential for horticultural crops. It involves the precise modification of plant DNA to enhance desired traits, increase crop yield, and confer resistance to pests, diseases, and environmental stresses. Molecular tools like CRISPR-Cas9, TALENs, and ZFNs act as molecular scissors, allowing for the targeted alteration of specific DNA sequences with unprecedented accuracy. This technology allows scientists to introduce or enhance beneficial traits in crops, including disease resistance, improved nutrition, and drought tolerance.
Horticulture is a critical component of global food production and human well-being as it involves the cultivation and management of plants for food, aesthetics, and medicinal purposes [1]. Horticultural crops, such as fruits, vegetables, ornamental plants, and medicinal herbs, not only contribute to the nutritional needs of populations worldwide but also enhance the visual appeal of our surroundings. However, the productivity and quality of horticultural crops face significant constraints due to challenges such as biotic and abiotic stresses, limited genetic variation, and increasing demands for improved traits [2].
Plant breeders have employed various techniques, such as hybridization, selection, and genetic manipulation, to enhance horticultural crops [3]. These approaches have contributed to improved yield, disease resistance, and other desirable traits [4]. However, traditional breeding methods have limitations, including long breeding cycles, limited genetic variation, and complex genetic architectures [5]. Genome editing has emerged as a transformative technology with the potential to revolutionize crop improvement, including horticultural crops [6]. By enabling precise modifications in an organism’s DNA, genome editing offers an efficient method to manipulate specific genes and traits [7]. This technology holds immense promise for expediting crop improvement, overcoming genetic barriers, and addressing specific challenges in horticultural crops.
The CRISPR-Cas9 system is widely used as a genome editing tool, utilizing guide RNA to direct the Cas9 enzyme for precise DNA cleavage and modifications [8]. TALENs and ZFNs are alternative genome editing tools also utilized in horticultural crop research [9]. These tools enable the precise editing of plant genomes by targeting specific genes associated with desired traits. CRISPR-Cas9 has proven effective in improving important traits in various horticultural crops through targeted modifications [10]. By designing specific gRNAs, researchers can direct the Cas9 enzyme to target genes associated with traits of interest, including disease resistance, abiotic stress tolerance, nutritional content, and yield-related characteristics [11]. The precise nature of CRISPR-Cas9 allows for the introduction of beneficial mutations or targeted gene knockouts, simulating natural genetic variations and accelerating the breeding process [12].
TALENs and ZFNs, along with CRISPR-Cas9, have been employed in horticultural crop research as genome editing tools (Figure 1). TALENs and ZFNs utilize engineered DNA-binding proteins that can be customized to target specific genomic sequences [13]. Similar to CRISPR-Cas9, these tools induce targeted DNA cleavage and subsequent modifications at the desired genomic sites. TALENs employ DNA-binding domains derived from transcription activator-like effectors (TALEs), which are naturally occurring proteins in plant pathogenic bacteria [14]. Engineered TALE domains are utilized to bind specific DNA sequences and are fused with a nuclease domain to induce DNA cleavage [15]. In contrast, ZFNs are hybrid proteins that combine engineered zinc finger DNA-binding domains with the FokI nuclease domain derived from the FokI restriction enzyme [16]. The zinc finger domains are designed to recognize targeted DNA sequences, and the FokI domain cleaves the DNA at the desired site [17].
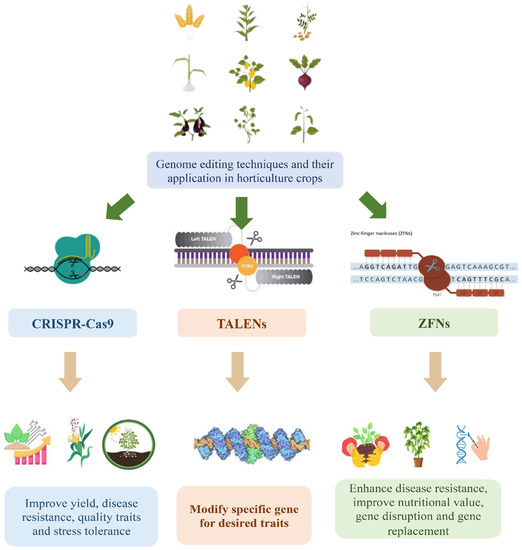
Figure 1. Genome editing in horticulture crops.
These genome editing tools offer researchers the ability to precisely modify genes associated with desired traits in horticultural crops, facilitating the development of improved varieties [18]. By utilizing these tools, scientists can expedite the enhancement of traits such as disease resistance, abiotic stress tolerance, nutritional content, and yield potential. In thiRes review, we eearchers examine the applications, methodologies, and potential impacts of genome editing tools, including CRISPR-Cas9, TALENs, and ZFNs, in horticultural crops.
.
The limited genetic diversity in horticultural crops poses challenges for developing improved varieties with enhanced traits [50][48]. CRISPR-Cas9 facilitates the accurate introduction of genetic variations, replicating the genetic diversity observed in wild relatives or closely related species [51][49]. Targeted modifications of specific genes or regulatory elements unlock untapped genetic potential and expand the available variation for crop enhancement [52][50], fostering the development of resilient, productive, and nutritionally valuable horticultural crops.
2. CRISPR-Cas9 in Horticultural Crops
The CRISPR-Cas9 system has gained prominence for its user-friendly nature, efficiency, and adaptability in genetic manipulation across organisms, including horticultural crops [21][19]. Based on bacteria’s defense mechanism against viral infections, CRISPR-Cas9 is a powerful tool [22][20] that enables precise genome editing in plants. This technology empowers researchers to target specific genes associated with desirable traits and introduce modifications to enhance agricultural characteristics [23][21]. This level of precision allows for the introduction of advantageous mutations, gene disruption, and the substitution of specific DNA sequences, leading to desired alterations in traits such as disease resistance, abiotic stress tolerance, nutritional composition, and yield-related attributes [24][22]. Table 21 presents examples of horticultural crops that have undergone genome editing using the CRISPR-Cas9 technique, along with the specific genes targeted and the resulting modified genetic traits. These modifications have led to significant improvements in fruit ripening, disease resistance, flowering time, tuberization, grain quality, and other desirable characteristics in the respective crops.Table 21.
Horticultural crops subjected to genome editing techniques, modified genetic traits in various plant species.
| Crops | Modified Gene(s) | Trait/Function | Reference |
|---|---|---|---|
| Tomato | E8, Phytoene desaturase (PDS), SlDELLA | Enhanced fruit ripening, delayed fruit senescence, reduced plant height | [25][23] |
| Potato | StCDF1 | Increased tuberization and yield | [26][24] |
| Wheat | TaGW2, Puroindoline genes | Enhanced thousand grain weight, improved grain quality | [27][25] |
| Citrus | CsPDS | Improved disease resistance, reduced ethylene production | [28][26] |
| Strawberry | FaTM6 | Petal and stamen development | [29][27] |
| Grape | VvWRKY52, VvWRKY2 | Enhanced disease resistance, improved abiotic stress tolerance | [30][28] |
| Brassica oleracea | XccR5-89.2 | Improved resistance to blackleg disease | [31][29] |
| Mushroom (Agaricus bisporus) | Polyphenol oxidase (PPO) genes | Reduced browning and improved shelf life | [32][30] |
| Banana | MaACO1 | Promotes the shelf life of banana | [33][31] |
| Carrot | DcCCD4 | Different colored taproots in carrots | [34][32] |
| Strawberry | FaGAST1 | Increased fruit size | [35][33] |
| Cucumis melo | CmACO1 | Extends the shelf-life | [36][34] |
| Capsicum annuum | CaERF28 | Anthracnose resistance | [37][35] |
| Rose | RhEIN2 | Ethylene insensitivity in rose | [38][36] |
| Melon | eIF4E | Virus resistance and male sterility | [39][37] |
| Tomato | SlMAPK3 | Reduced drought tolerance | [40][38] |
| Brassica napus | FAD2 | Catalyzes the desaturation of oleic acid | [41][39] |
| Kiwifruit | AcBFT | Reduce plant dormancy | [42][40] |
| Tomato | SlMYC2 | Fruit Resistance to Botrytis cinerea | [43][41] |
| Soybean | GmFATB1 | Reduce saturated fatty acids | [44][42] |
| Kiwi fruit | AcCBF3 | Dwarf plants and enhanced freezing tolerance | [45][43] |
| Sweet Potato | IbGBSSI and IbSBEII | Improvement of starch quality | [46][44] |
| Papaya | phytoene desaturase (CpPDS) | Inducing a visually scorable albino phenotype | [47][45] |
| Eggplant | SmelPPO4, SmelPPO5, and SmelPPO6 | Reduces fruit flesh browning | [48][46] |
| Cassava | eIF4E | Reduces cassava brown streak disease symptom | [49][47] |
3. TALENs (Transcription Activator-like Effector Nucleases)
TALENs, along with CRISPR-Cas9, are widely employed as genome editing tools for precise gene modifications in horticultural crops [114][51]. TALENs are engineered nucleases capable of inducing double-strand breaks (DSBs) at specific DNA sequences, enabling targeted gene editing [128][52]. Comprising a customizable DNA-binding domain derived from transcription activator-like effectors (TALEs) and a nuclease domain typically derived from the FokI endonuclease, TALENs offer a dual-component design [129][53]. The DNA-binding domain of TALENs is constructed using multiple repeats of TALEs, each recognizing a specific nucleotide in the target DNA sequence [130][54]. These TALEs consist of repeat units typically containing 33–35 amino acids in a central repeat region [131][55]. The specificity of TALENs is achieved through customizable repeat variable di-residues (RVDs) within each repeat unit, where different RVDs recognize different nucleotides, enabling the design of highly specific TALENs [132][56]. The nuclease domain of TALENs is derived from the FokI endonuclease, which requires dimerization for its DNA cleavage activity [133][57]. TALENs are designed as pairs, with each TALEN targeting one DNA strand [134][58]. Upon binding to their target sites, the FokI nuclease domains of the TALENs dimerize, forming a functional nuclease complex that induces double-strand breaks (DSBs) at the target site [135][59].4. ZFNs (Zinc Finger Nucleases)
ZFNs, an engineered nuclease class, have been utilized for genome editing in horticultural crops [159][60]. Comprising two main components, ZFPs and a nuclease domain from FokI endonuclease, ZFNs exhibit sequence-specific DNA recognition [160,161][61][62]. Each zinc finger module, approximately 30 amino acids in length, targets three DNA bases, and multiple modules enable precise targeting of specific DNA sequences [162,163][63][64]. ZFNs are employed in pairs, with each ZFN targeting one DNA strand [164][65]. The binding of the ZFNs to their target sites leads to FokI nuclease domain dimerization, generating a functional nuclease complex that induces double-strand breaks (DSBs) at the target site [135][59].5. Regulatory and Ethical Considerations
Genome editing has emerged as a promising approach for sustainable agriculture and crop enhancement, aiming to create transgene-free plants [186][66]. However, to ensure responsible and safe utilization of this technology, it is imperative to address key regulatory and ethical aspects. As genome editing advances, a thorough understanding of international regulations, regional policies, and ethical implications becomes essential in promoting its widespread adoption for crop improvement in agriculture.References
- Okunlola, A.I.; Adepoju, A.O.; Akinpetide, E.O. The significant role of horticulture in environmental aesthetics and management. Int. J. Hortic. 2016, 6, 17.
- Salgotra, R.K.; Chauhan, B.S. Genetic Diversity, Conservation, and Utilization of Plant Genetic Resources. Genes 2023, 14, 174.
- Borlaug, N.E. Contributions of Conventional Plant-Breeding to Food-Production. Science 1983, 219, 689–693.
- Sharma, H.C.; Crouch, J.H.; Sharma, K.K.; Seetharama, N.; Hash, C.T. Applications of biotechnology for crop improvement: Prospects and constraints. Plant Sci. 2002, 163, 381–395.
- Beaver, J.S.; Osorno, J.M. Achievements and limitations of contemporary common bean breeding using conventional and molecular approaches. Euphytica 2009, 168, 145–175.
- Xiong, J.S.; Ding, J.; Li, Y. Genome-editing technologies and their potential application in horticultural crop breeding. Hortic. Res. 2015, 2, 15019.
- Doudna, J.A.; Charpentier, E. The new frontier of genome engineering with CRISPR-Cas9. Science 2014, 346, 1258096.
- Thurtle-Schmidt, D.M.; Lo, T.W. Molecular biology at the cutting edge: A review on CRISPR/CAS9 gene editing for undergraduates. Biochem. Mol. Biol. Edu. 2018, 46, 195–205.
- Xu, J.M.; Hua, K.; Lang, Z.B. Genome editing for horticultural crop improvement. Hortic. Res. 2019, 6, 113.
- Erpen-Dalla Corte, L.; Mahmoud, L.M.; Moraes, T.S.; Mou, Z.L.; Grosser, J.W.; Dutt, M. Development of Improved Fruit, Vegetable, and Ornamental Crops Using the CRISPR/Cas9 Genome Editing Technique. Plants 2019, 8, 601.
- Zhang, D.Q.; Zhang, Z.Y.; Unver, T.; Zhang, B.H. CRISPR/Cas: A powerful tool for gene function study and crop improvement. J. Adv. Res. 2021, 29, 207–221.
- Rani, R.; Yadav, P.; Barbadikar, K.M.; Baliyan, N.; Malhotra, E.V.; Singh, B.K.; Kumar, A.; Singh, D. CRISPR/Cas9: A promising way to exploit genetic variation in plants. Biotechnol. Lett. 2016, 38, 1991–2006.
- Gaj, T.; Gersbach, C.A.; Barbas, C.F. ZFN, TALEN, and CRISPR/Cas-based methods for genome engineering. Trends Biotechnol. 2013, 31, 397–405.
- Sun, N.; Zhao, H.M. Transcription activator-like effector nucleases (TALENs): A highly efficient and versatile tool for genome editing. Biotechnol. Bioeng. 2013, 110, 1811–1821.
- Joung, J.K.; Sander, J.D. INNOVATION TALENs: A widely applicable technology for targeted genome editing. Nat. Rev. Mol. Cell Bio. 2013, 14, 49–55.
- Li, T.; Huang, S.; Jiang, W.Z.; Wright, D.; Spalding, M.H.; Weeks, D.P.; Yang, B. TAL nucleases (TALNs): Hybrid proteins composed of TAL effectors and FokI DNA-cleavage domain. Nucleic Acids Res. 2011, 39, 359–372.
- Pattanayak, V.; Ramirez, C.L.; Joung, J.K.; Liu, D.R. Revealing off-target cleavage specificities of zinc-finger nucleases by in vitro selection. Nat. Methods 2011, 8, 765–770.
- Osakabe, Y.; Osakabe, K. Genome Editing with Engineered Nucleases in Plants. Plant Cell Physiol. 2015, 56, 389–400.
- Noman, A.; Aqeel, M.; He, S.L. CRISPR-Cas9: Tool for Qualitative and Quantitative Plant Genome Editing. Front. Plant Sci. 2016, 7, 1740.
- Rao, M.J.; Wang, L.Q. CRISPR/Cas9 technology for improving agronomic traits and future prospective in agriculture. Planta 2021, 254, 68.
- Rasheed, A.; Barqawi, A.A.; Mahmood, A.; Nawaz, M.; Shah, A.N.; Bay, D.H.; Alahdal, M.A.; Hassan, M.U.; Qari, S.H. CRISPR/Cas9 is a powerful tool for precise genome editing of legume crops: A review. Mol. Biol. Rep. 2022, 49, 5595–5609.
- Urnov, F.D.; Rebar, E.J.; Holmes, M.C.; Zhang, H.S.; Gregory, P.D. Genome editing with engineered zinc finger nucleases. Nat. Rev. Genet. 2010, 11, 636–646.
- Wai, A.H.; Naing, A.H.; Lee, D.J.; Kim, C.K.; Chung, M.Y. Molecular genetic approaches for enhancing stress tolerance and fruit quality of tomato. Plant Biotechnol. Rep. 2020, 14, 515–537.
- Gonzales, L.R.; Shi, L.; Bergonzi, S.B.; Oortwijn, M.; Franco-Zorrilla, J.M.; Solano-Tavira, R.; Visser, R.G.F.; Abelenda, J.A.; Bachem, C.W.B. Potato CYCLING DOF FACTOR 1 and its lncRNA counterpart StFLORE link tuber development and drought response. Plant J. 2021, 105, 855–869.
- Henry, R.J.; Furtado, A.; Rangan, P. Wheat seed transcriptome reveals genes controlling key traits for human preference and crop adaptation. Curr. Opin. Plant Biol. 2018, 45, 231–236.
- Yang, W.; Ren, J.; Liu, W.; Liu, D.; Xie, K.; Zhang, F.; Wang, P.; Guo, W.; Wu, X. An efficient transient gene expression system for protein subcellular localization assay and genome editing in citrus protoplasts. Hortic. Plant J. 2023, 9, 425–436.
- Martin-Pizarro, C.; Trivino, J.C.; Pose, D. Functional analysis of the TM6 MADS-box gene in the octoploid strawberry by CRISPR/Cas9-directed mutagenesis. J. Exp. Bot. 2019, 70, 885–895.
- Capriotti, L.; Baraldi, E.; Mezzetti, B.; Limera, C.; Sabbadini, S. Biotechnological Approaches: Gene Overexpression, Gene Silencing, and Genome Editing to Control Fungal and Oomycete Diseases in Grapevine. Int. J. Mol. Sci. 2020, 21, 5701.
- Afrin, K.S.; Rahim, M.A.; Jung, H.J.; Park, J.I.; Kim, H.T.; Nou, I.S. Development of Molecular Marker through Genome Realignment for Specific Detection of Xanthomonas campestris pv. campestris Race 5, a Pathogen of Black Rot Disease. J. Microbiol. Biotechnol. 2019, 29, 785–793.
- Waltz, E. CRISPR-edited crops free to enter market, skip regulation. Nat. Biotechnol. 2016, 34, 582.
- Hu, C.H.; Sheng, O.; Deng, G.M.; He, W.D.; Dong, T.; Yang, Q.S.; Dou, T.X.; Li, C.Y.; Gao, H.J.; Liu, S.W.; et al. CRISPR/Cas9-mediated genome editing of MaACO1 (aminocyclopropane-1-carboxylate oxidase 1) promotes the shelf life of banana fruit. Plant Biotechnol. J. 2021, 19, 654–656.
- Li, T.; Deng, Y.J.; Liu, J.X.; Duan, A.Q.; Liu, H.; Xiong, A.S. DcCCD4 catalyzes the degradation of alpha-carotene and beta-carotene to affect carotenoid accumulation and taproot color in carrot. Plant J. 2021, 108, 1116–1130.
- Abdullah; Faraji, S.; Mehmood, F.; Malik, H.M.T.; Ahmed, I.; Heidari, P.; Poczai, P. The GASA Gene Family in Cacao (Theobroma cacao, Malvaceae): Genome Wide Identification and Expression Analysis. Agronomy 2021, 11, 1425.
- Nonaka, S.; Ito, M.; Ezura, H. Targeted modification of CmACO1 by CRISPR/Cas9 extends the shelf-life of Cucumis melo var. reticulatus melon. Front. Genome Ed. 2023, 5, 1176125.
- Mishra, R.; Mohanty, J.N.; Mahanty, B.; Joshi, R.K. A single transcript CRISPR/Cas9 mediated mutagenesis of CaERF28 confers anthracnose resistance in chilli pepper (Capsicum annuum L.). Planta 2021, 254, 5.
- Wang, C.P.; Li, Y.; Wang, N.; Yu, Q.; Li, Y.H.; Gao, J.P.; Zhou, X.F.; Ma, N. An efficient CRISPR/Cas9 platform for targeted genome editing in rose (Rosa hybrida). J. Integr. Plant Biol. 2023, 65, 895–899.
- Pechar, G.S.; Donaire, L.; Gosalvez, B.; Garcia-Almodovar, C.; Sanchez-Pina, M.A.; Truniger, V.; Aranda, M.A. Editing melon eIF4E associates with virus resistance and male sterility. Plant Biotechnol. J. 2022, 20, 2006–2022.
- Wang, L.; Chen, L.; Li, R.; Zhao, R.R.; Yang, M.J.; Sheng, J.P.; Shen, L. Reduced Drought Tolerance by CRISPR/Cas9-Mediated SlMAPK3 Mutagenesis in Tomato Plants. J. Agr. Food Chem. 2017, 65, 8674–8682.
- Okuzaki, A.; Ogawa, T.; Koizuka, C.; Kaneko, K.; Inaba, M.; Imamura, J.; Koizuka, N. CRISPR/Cas9-mediated genome editing of the fatty acid desaturase 2 gene in Brassica napus. Plant Physiol. Biochem. 2018, 131, 63–69.
- Herath, D.; Voogd, C.; Mayo-Smith, M.; Yang, B.; Allan, A.C.; Putterill, J.; Varkonyi-Gasic, E. CRISPR-Cas9-mediated mutagenesis of kiwifruit BFT genes results in an evergrowing but not early flowering phenotype. Plant Biotechnol. J. 2022, 20, 2064–2076.
- Shu, P.; Li, Z.Y.; Min, D.D.; Zhang, X.H.; Ai, W.; Li, J.Z.; Zhou, J.X.; Li, Z.L.; Li, F.J.; Li, X.A. CRISPR/Cas9-Mediated SlMYC2 Mutagenesis Adverse to Tomato Plant Growth and MeJA-Induced Fruit Resistance to Botrytis cinerea. J. Agr. Food Chem. 2020, 68, 5529–5538.
- Ma, J.; Sun, S.; Whelan, J.; Shou, H.X. CRISPR/Cas9-Mediated Knockout of GmFATB1 Significantly Reduced the Amount of Saturated Fatty Acids in Soybean Seeds. Int. J. Mol. Sci. 2021, 22, 3877.
- Wang, R. What Makes ‘Hayward’ Kiwifruit Store so Well? The Biological Basis for the Postharvest Behaviour of ‘Hayward’ Kiwifruit. Ph.D. Thesis, The University of Auckland, Auckland, New Zealand, 2021.
- Wang, H.X.; Wu, Y.L.; Zhang, Y.D.; Yang, J.; Fan, W.J.; Zhang, H.; Zhao, S.S.; Yuan, L.; Zhang, P. CRISPR/Cas9-Based Mutagenesis of Starch Biosynthetic Genes in Sweet Potato (Ipomoea Batatas) for the Improvement of Starch Quality. Int. J. Mol. Sci. 2019, 20, 4702.
- Brewer, S.E.; Chambers, A.H. CRISPR/Cas9-mediated genome editing of phytoene desaturase in Carica papaya L. J. Hortic. Sci. Biotechnol. 2022, 97, 580–592.
- Maioli, A.; Gianoglio, S.; Moglia, A.; Acquadro, A.; Valentino, D.; Milani, A.M.; Prohens, J.; Orzaez, D.; Granell, A.; Lanteri, S.; et al. Simultaneous CRISPR/Cas9 Editing of Three PPO Genes Reduces Fruit Flesh Browning in Solanum melongena L. Front. Plant Sci. 2020, 11, 607161.
- Gomez, M.A.; Lin, Z.D.; Moll, T.; Chauhan, R.D.; Hayden, L.; Renninger, K.; Beyene, G.; Taylor, N.J.; Carrington, J.C.; Staskawicz, B.J.; et al. Simultaneous CRISPR/Cas9-mediated editing of cassava eIF4E isoforms nCBP-1 and nCBP-2 reduces cassava brown streak disease symptom severity and incidence. Plant Biotechnol. J. 2019, 17, 421–434.
- Krishna, H.; Alizadeh, M.; Singh, D.; Singh, U.; Chauhan, N.; Eftekhari, M.; Sadh, R.K. Somaclonal variations and their applications in horticultural crops improvement. 3 Biotech 2016, 6, 54.
- Søren, K.; Toni, W.; Christoph, D.; Hanne, C.T.; Magnus, R.; Morten Egevang, J.; Qiongxian, L.; Cynthia, V.; Emiko, M.; Jeppe Thulin, Ø.; et al. FIND-IT: Ultrafast mining of genome diversity. bioRxiv 2021.
- Wagh, S.G.; Pohare, M.B. Current and Future Prospects of Plant Breeding with CRISPR/Cas. Current. J. Appl. Sci. Technol. 2019, 38, 1–17.
- Mahfouz, M.M.; Piatek, A.; Stewart, C.N. Genome engineering via TALENs and CRISPR/Cas9 systems: Challenges and perspectives. Plant Biotechnol. J. 2014, 12, 1006–1014.
- Lee, J.; Chung, J.H.; Kim, H.M.; Kim, D.W.; Kim, H. Designed nucleases for targeted genome editing. Plant Biotechnol. J. 2016, 14, 448–462.
- Sanjana, N.E.; Cong, L.; Zhou, Y.; Cunniff, M.M.; Feng, G.P.; Zhang, F. A transcription activator-like effector toolbox for genome engineering. Nat. Protoc. 2012, 7, 171–192.
- Morbitzer, R.; Romer, P.; Boch, J.; Lahaye, T. Regulation of selected genome loci using de novo-engineered transcription activator-like effector (TALE)-type transcription factors. Proc. Natl. Acad. Sci. USA 2010, 107, 21617–21622.
- Miller, J.C.; Tan, S.Y.; Qiao, G.J.; Barlow, K.A.; Wang, J.B.; Xia, D.F.; Meng, X.D.; Paschon, D.E.; Leung, E.; Hinkley, S.J.; et al. A TALE nuclease architecture for efficient genome editing. Nat. Biotechnol. 2011, 29, 143–148.
- Christian, M.L.; Demorest, Z.L.; Starker, C.G.; Osborn, M.J.; Nyquist, M.D.; Zhang, Y.; Carlson, D.F.; Bradley, P.; Bogdanove, A.J.; Voytas, D.F. Targeting G with TAL Effectors: A Comparison of Activities of TALENs Constructed with NN and NK Repeat Variable Di-Residues. PLoS ONE 2012, 7, e45383.
- Xiao, A.; Wu, Y.D.; Yang, Z.P.; Hu, Y.Y.; Wang, W.Y.; Zhang, Y.T.; Kong, L.; Gao, G.; Zhu, Z.Y.; Lin, S.; et al. EENdb: A database and knowledge base of ZFNs and TALENs for endonuclease engineering. Nucleic Acids Res. 2013, 41, D415–D422.
- Cermak, T.; Doyle, E.L.; Christian, M.; Wang, L.; Zhang, Y.; Schmidt, C.; Baller, J.A.; Somia, N.V.; Bogdanove, A.J.; Voytas, D.F. Efficient design and assembly of custom TALEN and other TAL effector-based constructs for DNA targeting. Nucleic Acids Res. 2011, 39, 7879.
- Puchta, H.; Fauser, F. Synthetic nucleases for genome engineering in plants: Prospects for a bright future. Plant J. 2014, 78, 727–741.
- Govindan, G.; Ramalingam, S. Programmable Site-Specific Nucleases for Targeted Genome Engineering in Higher Eukaryotes. J. Cell Physiol. 2016, 231, 2380–2392.
- Durai, S.; Mani, M.; Kandavelou, K.; Wu, J.; Porteus, M.H.; Chandrasegaran, S. Zinc finger nucleases: Custom-designed molecular scissors for genome engineering of plant and mammalian cells. Nucleic Acids Res. 2005, 33, 5978–5990.
- Mandell, J.G.; Barbas, C.F. Zinc finger tools: Custom DNA-binding domains for transcription factors and nucleases. Nucleic Acids Res. 2006, 34, W516–W523.
- Hall, T.M.T. Multiple modes of RNA recognition by zinc finger proteins. Curr. Opin. Struct. Biol. 2005, 15, 367–373.
- Cathomen, T.; Joung, J.K. Zinc-finger nucleases: The next generation emerges. Mol. Ther. 2008, 16, 1200–1207.
- Kim, H.; Kim, J.S. A guide to genome engineering with programmable nucleases. Nat. Rev. Genet. 2014, 15, 321–334.
- Gao, C. Genome engineering for crop improvement and future agriculture. Cell 2021, 184, 1621–1635.
More