Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 3 by Conner Chen and Version 2 by Conner Chen.
Starting from the morphological function of fish, there are many swimming modes of fish in nature, the most authoritative classification of which, based on the organ of propulsion, was first proposed by Breder and Webb [38]. There are two main categories of propulsion methods, body caudal fin (BCF) propulsion and median paired fin (MPF) propulsion. In 1978, Lindsey classified the different families of fish species according to their mode of propulsion, which provided the essential foundation for the improvement of the overall performance of undulating propulsion biomimetic robots.
- underwater biomimetic robots
- undulating propulsion robots
- undulating propulsion mechanism
1. Introduction
Underwater robots have an important role to play in future marine development and underwater activities and have broad application prospects and great potential value [1][2][3][4][5]. Researchers have designed various underwater robots for underwater missions, such as remotely operated vehicles (ROVs) and autonomous underwater vehicles (AUVs) [6][7] (Figure 1). Traditional underwater robots are composed of rigid materials and most are propelled by propellers [8][9]. However, propeller propulsion is unsuitable for implementation in harsh marine environments as it can damage aquatic life or the seabed or even become trapped in weeds [10][11]. In addition, the low efficiency and stability of propeller-driven craft in low-speed attitude adjustment conditions greatly limit the applications of propeller propulsion. Therefore, novel underwater propulsion methods are desired and are currently being investigated [7].
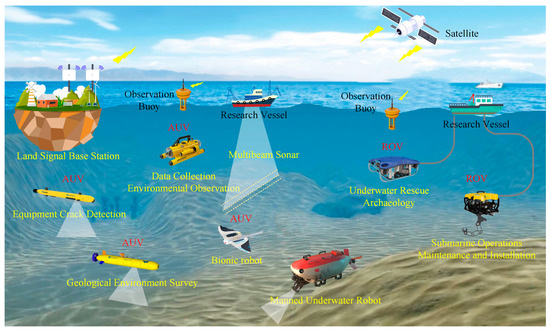
Figure 1.
Overview of ROV and AUV application scenarios.
At present, increasing attention is being paid to bionics-based propulsion mechanisms. This propulsive method is validated in marine environments as its principle is inspired by various marine organisms. The soft bodies, good flexibility and strong environmental adaptability of marine organisms inspire new ideas for the development of underwater robots [12]. After a long period of evolution, fish have adapted to exploit the principle of hydrodynamics to move forward using tail fins or pectoral fins [13][14][15], which has become the best way to move in water [16][17]. The propulsion method employed by fish can maintain high maneuverability and change posture according to the changing environment. Inspired by the fish propulsion method, the first underwater bionic robot RoboTuna [18] was unveiled in 1994, and various types of underwater bionic robots continue to emerge. Equipped with various propulsion methods, underwater robots present positive development trends and broad application prospects [19][20][21][22][23] The term underwater biomimetic robot generally refers to robots that imitate the structure, function, or type of movement of fish species [24]. They have the basic movement characteristics of biomimetic fish and are continuously being optimized and improved on this basis. They have diverse functions, strong environmental adaptability and stability [25]. Undulating propulsion biomimetic robots are propelled by the undulating of pectoral fins, caudal fins and the body, an approach that has superior motion performance and can realize multiple motion modes [26]. In summary, undulating propulsion underwater biomimetic robots are increasingly favored by scholars due to their good maneuverability and stability.
2. The Swimming Mode of Fish
Starting from the morphological function of fish, there are many swimming modes of fish in nature, the most authoritative classification of which, based on the organ of propulsion, was first proposed by Breder and Webb [27]. There are two main categories of propulsion methods, body caudal fin (BCF) propulsion and median paired fin (MPF) propulsion. In 1978, Lindsey [28] classified the different families of fish species according to their mode of propulsion, which provided the essential foundation for the improvement of the overall performance of undulating propulsion biomimetic robots. Figure 2 shows the fish species that employ BCF mode propulsion [29]. For this propulsion mode, the main body types are anguilliform, subcarangiform, carangiform, thunniform and ostraciiform, and the representative fish species for these body types are the eel, trout, herring, tuna and longhorn cowfish, respectively. Fish employing BCF mode propulsion gradually transition from undulating to oscillatory propulsion according to their different driving methods, as shown by the dashed line in Figure 2. In the figure, the first three propulsion modes are undulating and the last two are oscillatory. BCF propulsion modes are characterized by a high swimming speed—swordfish can reach a speed of 110 km/h—and good energy efficiency—tuna can maintain 90% energy efficiency while swimming for a long time [30]. Fish with the anguilliform body type employ fluctuations of the whole body and tail fin. This type of fish has a slender body and a faster swimming speed. Subcarangiform and carangiform fish species are relatively similar; the difference is that the former uses 1/2 of their body length for fluctuations, while the latter uses only about 1/3 of their body length for propulsion [31][32]. Compared with the anguilliform model, the fluctuation range is small, but the caudal fluctuation frequency is high. The propulsion of thunniform fish comes from the tail fin [33]. Based on hydrodynamic models, thunniform propulsion has been estimated to produce mechanical efficiencies in the range of 80–90% [18][34][35]. The fastest fish (tuna, dolphin) use this propulsion mode [36][37][38][39]. Fish species with the ostraciiform body type adopt an oscillatory propulsion mode. This type of fish simply oscillates the caudal fin, which induces only a small thrust; thus, the speed of travel is relatively slow [40].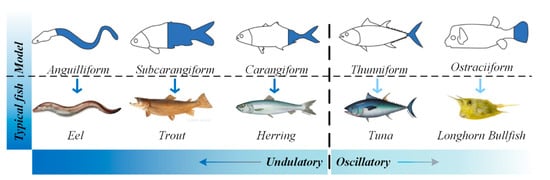
Figure 2. BCF propulsion mode [41].
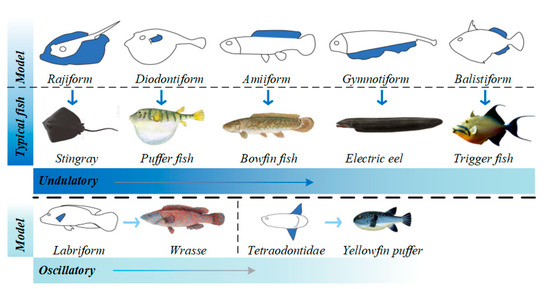
Figure 3. MPF propulsion mode [41].
References
- Isaka, K.; Tadami, N.; Fujiwara, A.; Watanabe, T.; Sugesawa, M.; Yamada, Y.; Yoshida, H.; Nakamura, T. Study on Drilling Resistance Reduction of a Seafloor Robotic Explorer Based on the Drilling Properties of Underwater Ground. In Proceedings of the 2019 IEEE/SICE International Symposium on System Integration (SII), Paris, France, 14–16 January 2019; pp. 718–723.
- Yamada, D.; Takebayashi, T.; Kato, H.; Sakagami, N.; Kawamura, S. Underwater Robot with Negative Pressure Effect Plates for Maintenance of Underwater Structures. In Proceedings of the 2019 IEEE/ASME International Conference On Advanced Intelligent Mechatronics (AIM), Hong Kong, China, 8–12 July 2019; pp. 1092–1097.
- Ru, J.Y.; Yu, H.; Liu, H.; Liu, J.Y.; Zhang, X.Y.; Xu, H.L. A Bounded Near-Bottom Cruise Trajectory Planning Algorithm for Underwater Vehicles. J. Mar. Sci. Eng. 2023, 11, 7.
- Leng, D.X.; Shao, S.; Xie, Y.C.; Wang, H.H.; Liu, G.J. A brief review of recent progress on deep sea mining vehicle. Ocean. Eng. 2021, 228, 108565.
- Zheng, J.Z.; Wang, J.X.; Guo, X.; Huntrakul, C.; Wang, C.; Xie, G.M. Biomimetic Electric Sense-Based Localization: A Solution for Small Underwater Robots in a Large-Scale Environment. IEEE Robot. Autom. Mag. 2022, 29, 50–65.
- Colgate, J.E.; Lynch, K.M. Mechanics and control of swimming: A review. IEEE J. Ocean. Eng. 2004, 29, 660–673.
- Bu, K.L.; Gong, X.B.; Yu, C.L.; Xie, F. Biomimetic Aquatic Robots Based on Fluid-Driven Actuators: A Review. J. Mar. Sci. Eng. 2022, 10, 735.
- Chutia, S.; Kakoty, N.M.; Deka, D. A Review of Underwater Robotics, Navigation, Sensing Techniques and Applications. In Proceedings of the Advances in Robotics (AIR’17), New Delhi, India, 28 June–2 July 2017.
- Wang, J.; Wu, Z.X.; Dong, H.J.; Tan, M.; Yu, J.Z. Development and Control of Underwater Gliding Robots: A Review. IEEE CAA J. Autom. Sin. 2022, 9, 1543–1560.
- Castano, M.L.; Tan, X.B. Model Predictive Control-Based Path-Following for Tail-Actuated Robotic Fish. J. Dyn. Syst. Meas. Control. Trans. Asme 2019, 141, 071012.
- Ma, Y.T.; Ye, R.D.; Zheng, R.; Geng, L.B.; Yang, Y. A highly mobile ducted underwater robot for subsea infrastructure inspection. In Proceedings of the 2016 IEEE International Conference on Cyber Technology in Automation, Control, and Intelligent Systems (CYBER), Chengdu, China, 19–22 June 2016; pp. 397–400.
- Kruiper, R.; Vincent, J.F.V.; Abraham, E.; Soar, R.C.; Konstas, I.; Chen-Burger, J.; Desmulliez, M.P.Y. Towards a Design Process for Computer-Aided Biomimetics. Biomimetics 2018, 3, 14.
- Bianchi, G.; Maffi, L.; Tealdi, M.; Cinquemani, S. A Bioinspired Cownose Ray Robot for Seabed Exploration. Biomimetics 2023, 8, 30.
- Shao, H.; Dong, B.B.; Zheng, C.Z.; Li, T.; Zuo, Q.Y.; Xu, Y.H.; Fang, H.T.; He, K.; Xie, F.R. Thrust Improvement of a Biomimetic Robotic Fish by Using a Deformable Caudal Fin. Biomimetics 2022, 7, 113.
- Kocak, M.; Yazici, M.V.; Akdal, E.; Can, F.C.; Gezgin, E. Utilization of Function Generation Synthesis on Biomimetics: A Case Study on Moray Eel Double Jaw Design. Biomimetics 2022, 7, 145.
- Salazar, R.; Campos, A.; Fuentes, V.; Abdelkefi, A. A review on the modeling, materials, and actuators of aquatic unmanned vehicles. Ocean. Eng. 2019, 172, 257–285.
- Silva, A.T.; Baerum, K.M.; Hedger, R.D.; Baktoft, H.; Fjeldstad, H.P.; Gjelland, K.O.; Okland, F.; Forseth, T. The effects of hydrodynamics on the three-dimensional downstream migratory movement of Atlantic salmon. Sci. Total Environ. 2020, 705, 135773.
- Triantafyllou, M.S.; Triantafyllou, G.S. An efficient swimming machine. Sci. Am. 1995, 272, 64–70.
- Yu, J.Z.; Liu, J.C.; Wu, Z.X.; Fang, H. Depth Control of a Bioinspired Robotic Dolphin Based on Sliding-Mode Fuzzy Control Method. IEEE Trans. Ind. Electron. 2018, 65, 2429–2438.
- Meng, Y.; Wu, Z.X.; Li, Y.T.; Chen, D.; Tan, M.; Yu, J.Z. Vision-Based Underwater Target Following Control of an Agile Robotic Manta With Flexible Pectoral Fins. IEEE Robot. Autom. Lett. 2023, 8, 2277–2284.
- Yu, J.Z.; Wang, C.; Xie, G.M. Coordination of Multiple Robotic Fish With Applications to Underwater Robot Competition. IEEE Trans. Ind. Electron. 2016, 63, 1280–1288.
- Yang, Q.; Li, G.; Mu, W.; Liu, G.; Sun, H. Identification of crack length and angle at the center weld seam of offshore platforms using a neural network approach. J. Mar. Sci. Eng. 2020, 8, 40.
- Leng, D.X.; Liu, D.; Li, H.Y.; Jin, B.; Liu, G.J. Internal flow effect on the cross-flow vortex-induced vibration of marine risers with different support methods. Ocean Eng. 2022, 257, 111487.
- Szymak, P.; Praczyk, T.; Naus, K.; Szturomski, B.; Malec, M.; Morawski, M. Research on Biomimetic Underwater Vehicles for Underwater ISR. In Proceedings of the Ground/Air Multisensor Interoperability, Integration, and Networking for Persistent ISR VII, Baltimore, MD, USA, 18–20 April 2016.
- Wang, R.; Wang, S.; Wang, Y.; Cheng, L.; Tan, M. Development and Motion Control of Biomimetic Underwater Robots: A Survey. IEEE Trans. Syst. Man Cybern. Syst. 2022, 52, 833–844.
- Wang, A.; Liu, G.; Wang, X.; Fu, B. Development and Analysis of Body and/or Caudal Fin Biomimetic Robot Fish. J. Mech. Eng. 2016, 52, 137–146.
- Webb, P.W. Form and function in fish swimming. Sci. Am. 1984, 251, 72–83.
- Lindsey, C.C. Form, function and locomotory habits in fish. Fish Physiol. 1978, 12–17.
- Sfakiotakis, M.; Lane, D.M.; Davies, J.B.C. Review of fish swimming modes for aquatic locomotion. IEEE J. Ocean. Eng. 1999, 24, 237–252.
- Wang, C.; Lu, J.; Ding, X.; Jiang, C.; Yang, J.; Shen, J. Design, modeling, control, and experiments for a fish-robot-based IoT platform to enable smart ocean. IEEE Internet Things J. 2021, 8, 9317–9329.
- Guanrong, H.; Zhenlong, W.; Jian, L.I.; Yangwei, W. Development of a Caudal-Fin-Propelled Micro Robot Fish Based on Flexible Fins. Robot 2008, 30, 171–175,181.
- Butail, S.; Bartolini, T.; Porfiri, M. Collective Response of Zebrafish Shoals to a Free-Swimming Robotic Fish. PLoS ONE 2013, 8, e76123.
- Mitin, I.; Korotaev, R.; Ermolaev, A.; Mironov, V.; Lobov, S.A.; Kazantsev, V.B. Bioinspired Propulsion System for a Thunniform Robotic Fish. Biomimetics 2022, 7, 215.
- Fish, F.E. Transitions from drag-based to lift-based propulsion in mammalian swimming. Am. Zool. 1996, 36, 628–641.
- Rohr, J.J.; Fish, F.E. Strouhal numbers and optimization of swimming by odontocete cetaceans. J. Exp. Biol. 2004, 207, 1633–1642.
- Wiguna, T.; Heo, S.; Park, H.C.; Goo, N.S. Design and Experimental Parameteric Study of a Fish Robot Actuated by Piezoelectric Actuators. J. Intell. Mater. Syst. Struct. 2009, 20, 751–758.
- Edwards, E.F. Duration of unassisted swimming activity for spotted dolphin (Stenella attenuata) calves: Implications for mother-calf separation during tuna purse-seine sets. Fish. Bull. 2006, 104, 125–135.
- Brill, R.W. Selective advantages conferred by the high performance physiology of tunas, billfishes, and dolphin fish. Comp. Biochem. Physiol. A Physiol. 1996, 113, 3–15.
- Yu, J.Z.; Wu, Z.X.; Su, Z.S.; Wang, T.Z.; Qi, S.W. Motion Control Strategies for a Repetitive Leaping Robotic Dolphin. IEEE-Asme Trans. Mechatron. 2019, 24, 913–923.
- Chan, W.L.; Kang, T.; Lee, Y.J. Experiments and identification of an ostraciiform fish robot. In Proceedings of the 2007 IEEE International Conference on Robotics and Biomimetics, Macau, China, 5–8 December 2017; pp. 530–534.
- Li, Y.; Xu, Y.; Wu, Z.; Ma, L.; Guo, M.; Li, Z.; Li, Y. A comprehensive review on fish-inspired robots. Int. J. Adv. Robot. Syst. 2022, 19.
- Ikeda, M.; Hikasa, S.; Watanabe, K.; Nagai, I. Propulsive force analysis of a pectoral fin for rajiform type fish robots from fluid dynamic aspects. In Proceedings of the Eighteenth International Symposium on Artificial Life and Robotics (AROB 18th ‘13), Daejeon, Republic of Korea, 30 January–1 February 2013; pp. 65–68.
- Miyazaki, H.; Onoda, A.; Terada, H.; Nakajima, M. Species Identification of Pufferfish Products Using RAPD Analysis. Food Hyg. Saf. Sci. 2017, 58, 75–79.
- Bogan, S.; Taverne, L.; Agnolin, F. First Triassic and oldest record of a South American amiiform fish: Caturus sp from the Los Menucos Group (lower Upper Triassic), Rio Negro province, Argentina. Geol. Belg. 2013, 16, 191–195.
- Crampton, W.G.R. Gymnotiform fish: An important component of Amazonian floodplain fish communities. J. Fish Biol. 1996, 48, 298–301.
- George, A.B.; Westneat, M.W. Three-dimensional kinematic analyses reveal asymmetries in Xanthichthys auromarginatus (Balistidae) median fin biomechanics during steady balistiform swimming. Integr. Comp. Biol. 2021, 61, E306–E307.
- Zhang, R.; Hu, W. The numerical study on the propulsive mechanism of balistiform. Chin. J. Hydrodynomics 2020, 35, 258–265.
- Chinook Salmon. Oncorhynchus Tshawytscha. Feb. NOAA Fisheries. Available online: http://www.nmfs.noaa.gov/pr/species/fish/chinook-salmon.html (accessed on 7 July 2023).
- Sagong, W.; Jeon, W.-P.; Choi, H. Hydrodynamic characteristics of the sailfish (Istiophorus platypterus) and swordfish (Xiphias gladius) in gliding postures at their cruise speeds. PLoS ONE 2013, 8, e81323.
- Stingray. A-z Animals. Feb. Available online: https://a-z-animals.com/animals/stingray/?r (accessed on 7 July 2023).
More