2. Mouse Models of ASD
2.1. Mouse Models of Psychiatric Disorders
Currently, no animal models have been proven to suffer from known psychiatric diseases, share their causes and symptoms, and respond similarly to treatments validated in humans. Thus, homologous models of psychiatric diseases are rare. Previously, the main utility of animal models in pre-clinical psychiatry was to validate drugs that target psychiatric symptoms. For example, rodents exposed to an open-field environment experience a conflict between their fear-induced thigmotaxis and their desire to explore the new surroundings
[19][20]. A fearful animal would mainly explore the periphery of the field and avoid the center, particularly if it is brightly lit. For many years, this test, which is known as the open-field test, has been utilized to evaluate candidate anxiolytic drugs, demonstrating a high predictive value. Indeed, drugs that increased the exploration of the center of an open field in rodents demonstrated anxiolytic effects when administered to humans, although some exceptions were also observed
[20][21].
Despite the lack of homologous models of psychiatric diseases, recent efforts have allowed the development of valuable animal models that serve as investigative tools
[21][22][22,23]. Based on the original proposal of Willner
[23][24], animal models of human mental disorders are required to fulfill construct, face, and predictive validity. Although no model completely accounts for all three requirements and other authors have proposed different levels of validity
[24][25], researchers attempt to make their models as valuable as possible to ensure that they can translate their discoveries into
theour understanding of human disorders.
Construct validity ensures that the same etiological cause of the disease is responsible for the phenotype observed in the animal. To assess construct validity, it is necessary to understand the implicit or explicit hypothesis regarding the disease’s cause, such as the association between specific genes and their products and particular disorders. As a result, researchers have generated numerous genetically modified animals with genetic mutations similar to those found in humans or exposed animals to environmental factors associated with increased prevalence of certain pathologies. Behavioral and pharmacological analyses of these models are crucial in identifying associated phenotypic changes
[25][26]. These analyses allow the comparison of different etiological hypotheses and the exploration of pathophysiological pathways
[26][27].
Animal models are also expected to exhibit face validity, which means that the changes observed in the animal should be comparable to the symptoms seen in human patients. This requirement is based on the assumption that the phenotype observed in the animal arises from the same physiological processes as the human disease. However, this belief is not always accurate and can be difficult to confirm, especially since most psychiatric disorders have complex symptoms that may be caused by various genetic and environmental factors. To address this complexity, researchers have proposed the concept of endophenotype, which is a valuable tool that can advance
theour understanding of psychiatric disorders
[27][28].
The predictive validity of an animal model is demonstrated when a treatment tested on the animal has comparable effects on humans, as exemplified using benzodiazepines
[20][21]. Occasionally, these effects can be observed without thorough comprehension of the biological processes that underlie them. As a result, some researchers regard predictive models as only partially beneficial and subject to undesirable side effects, particularly when the mechanisms of the disease and treatment are not fully understood.
In summary, for an animal model of a psychiatric disorder to be valuable, it should have construct, face, and predictive validity, or at least some of these factors. The more the model imitates the etiological factors of a disease, manifests the phenotype/symptoms, and precisely forecasts the outcome of suggested treatments, the more advantageous it is to researchers who study the psychiatric disease.
2.2. Tools to Evaluate Face Validity of Mouse Models of ASD and Limitations Related to Sex
Mouse models of ASD are primarily evaluated based on their face validity. Initially, reports referred to three core symptoms of autism for diagnosis
[28][29], while the latest version of the Diagnostic and Statistical Manual of Mental Illnesses (DSM) focuses on two symptoms: persistent deficits in social communication and social interactions across multiple contexts, as well as the expression of restricted and repetitive behaviors, interests, and activities (including hyper- or hypo-reactivity to sensory stimuli)
[1]. Importantly, the severity of these symptoms varies highly between individuals with ASD, which is not usually considered in pre-clinical studies. Behavioral tests have been designed based on these diagnostic criteria and divided into two categories to measure these behaviors in rodents: (1) sociability and communication deficits and (2) repetitive and stereotyped behaviors
[29][30][30,31].
Initially, research into the influence of sex differences on behavior led to unintended consequences and the limiting of behavioral neuroscience research to male subjects because they were believed to be less influenced by the cyclic production of gonadal steroids that result from ovary function. Female behavior was only considered relevant in terms of understanding reproductive behavior, resulting in a poor understanding of sex differences in behavior and creating pre-clinical research biased in favor of male subjects
[31][19]. However,
theour current understanding suggests that female behavior may not be much different or difficult to study than male behavior
[32], and the extended mandate of funding agencies to include subjects of both sexes has opened a new wave of data that shows sex’s influence on a wide variety of behaviors, besides reproductive behaviors. Researchers will discuss the tests most commonly used to evaluate ASD-relevant behaviors and specify whether there are differences in their applicability or results related to the sex of the animal (
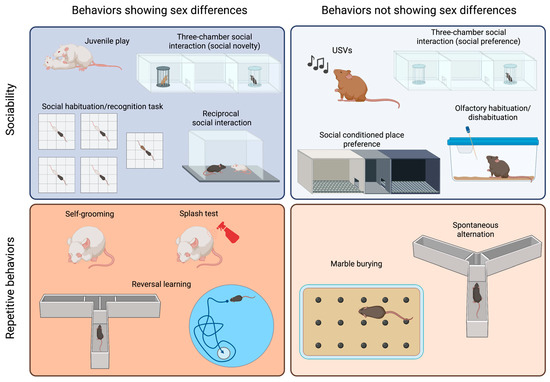
Figure 1).
Figure 1. Sex differences in tests commonly used to assess sociability and repetitive behaviors in ASD models. The tests are categorized based on the core behavior that they measure (sociability or repetitive behaviors) and whether they yield different results in male and female animals. Created with BioRender.com.
2.2.1. Tests to Evaluate Sociability and Communication Deficits in Mice
The species
Mus musculus is known to be highly social, exhibiting reciprocal social interactions, communal nesting, sexual and parenting behaviors, territorial scent marking, and aggressive behaviors
[33][34][35][36][33,34,35,36]. Many social assays have been reported in the field of behavioral neuroscience. However,
rein this
earchers article, we will specifically discuss the tests commonly used to evaluate social impairments in mouse models of ASD.
It is probable that the most popular test to assess sociability is the
three-chamber social interaction test, which has been chosen by numerous scientists because it allows for automatization of measurements and can be easily standardized
[37][38][37,38]. In this test, animals are initially allowed to explore a cage divided into three compartments. A stimulus animal is then placed in one of the lateral compartments, known as the “social side”, and either the time that the test mouse spends exploring the social stimulus or the time spent in the social chamber is measured. Different modifications to this test have been proposed, including changing the size of the cage and placing an object into the other lateral compartment
[39][40][39,40]. Usually, a young mouse of the same sex as the test mouse is employed as a stimulus. Although strain differences have been reported, most mice will prefer the social side and, therefore, spend more time in that compartment than in that containing an inanimate object
[37][39][41][37,39,41]. This test has the advantage of placing the stimulus mouse in a cage, which is unreachable by the test mouse, and it can be performed with aggressive animals, with many male adult mice being aggressive. In addition, this test is routinely used to measure social memory, adding a third phase in which a novel stimulus replaces the object, and the test mouse then shows a preference for such novelty. This test has been used to assess sociability in both males and females, and both sexes show similar levels of exploration of the young and same-sex social stimulus
[42][43][44][42,43,44] (
Figure 1, right panel). However, social novelty recognition is influenced by the sex and strain of the animal
[44] (
Figure 1, left panel). Additionally, this test can be employed to assess sex preference by presenting a female and a male adult mouse. Sex preference is also influenced by the sex and the strain of the animal
[44].
The
reciprocal social interaction test is certainly more ethologically relevant, as it allows the animals to freely interact with each other
[30][31]. However, it is also more time consuming, as videos should be scored manually (although automated measurement has been achieved using machine learning approaches
[45]). Besides the advantage of allowing the characterization of the mode of social engagement between the mice, a main concern is how animals should be paired, as the partner may influence the outcome of the test
[46][47][46,47]. Moreover, in strains such as CF1, CBA or CD1, which have been reported to exhibit high levels of aggressive behavior
[48], this test cannot be applied to adult males, as their aggression may overshadow any potential differences in sociability.
A similar paradigm can be used to explore
juvenile play, which is a social behavior typical of young animals. Although previous work on rats showed that females played less than males (reviewed in
[49]), reports on mice are heterogenous. C57BL/6J (B6) and CD1 females solicit play activities more often than males
[36][50][36,50], while outbred CF1 males performed more of these behaviors than females
[43] (
Figure 1, left panel). An evaluation of juvenile play in the four-core genotype (FCG) model in B6 background showed that XY males solicited play activities more often than any other group, suggesting that both gonadal hormones and sex chromosomes are relevant to the expression of this behavior
[36].
In addition to performing a social novelty trial in the three-chamber social interaction test, social memory can be evaluated via a
social habituation/recognition task [51]. In this test, animals are sequentially exposed to a stimulus mouse for a short period, showing habituation, and then exposed to a novel animal to evaluate their response to novelty. As for free social interaction, when aggressive animals are being assessed, stimuli can be presented in a small cage to avoid attacks.
Social conditioned place preference is a useful tool for the evaluation of the rewarding nature of social interactions in young mice
[52][53][52,53]. However, some strains (e.g., BALB/c) do not display social conditioning. This test shows no sex differences when performed with young animals
[52]. However, female and male adult mice exhibit a differential response to social conditioning that depends on previous social conditions, such as isolation versus group housing
[53].
Olfactory habituation/dishabituation to social odors is a valuable tool for the assessment of the response to a social stimulus, as it lacks the confounding effect of a second mouse with its own sociability levels. Mice tend to sniff a novel odor and then reduce their exploration as they acclimatize to the stimulus
[54]. A dishabituation effect is observed when a different odor is introduced, and animals reinstate a high level of sniffing. Using social odors, it is possible to evaluate whether animals can discriminate between the same and different social odors and assess whether these odors are more salient to some animals than others
[55]. No sex differences were observed in response to non-social odors
[56]. Females, however, are more sensitive than male to social odors
[57], possibly due to sex differences in the development of the olfactory system
[58].
Although it is not yet well understood how mice communicate,
ultrasonic vocalizations (USVs) appear to contribute to the communication of information and social bonding
[59][60][61][62][63][64][59,60,61,62,63,64]. This observation is especially relevant to pups, and the analysis of ultrasonic vocalizations in pups separated from their dam and nest is the gold standard method for the measurement of alterations in sociability in newborns
[61][64][61,64]. Pups of different strains perform different numbers of USVs, and each strain seems to have a unique repertoire of syllables
[59]. In young, adolescent, and adult mice, both males and females vocalize, but the frequency and quality of the vocalizations depend on the eliciting stimulus and strain
[60][65][60,65].
2.2.2. Tests to Evaluate Repetitive and Stereotyped Behaviors in Mice
Mice show spontaneous motor stereotypies, including self-grooming and burying, which can develop into repetitive behaviors if they persist for prolonged periods
[66].
Self-grooming is an innate behavior involved in hygiene maintenance and thermoregulation, which can be easily assessed in a home cage or novel environment
[40][43][55][67][68][40,43,55,67,68]. Adult male mice have been reported to spend more time self-grooming than females
[43]. The
splash test is an alternative method used to elicit self-grooming behavior, where a sucrose solution is splashed on the back of the animal, and the sweetness of the solution sustains the grooming behavior
[69]. The time spent grooming after the splash can be affected by the sex of the animal, although such an effect depends on the mouse strain being studied
[70].
In the
marble burying test, animals are exposed to an environment in which marble balls have been placed on top of a thick floor of bedding, which elicits the burying behavior in most mouse strains
[71][72][71,72]. The test involves quantifying the marbles buried at different times, typically lasting 20 min
[71]. Although most reports on marble burying behavior only involve males
[72], it has been shown that the estrous cycle alters the burying response in rats
[73].
Spontaneous alternation in the Y maze or T maze can also be evaluated in an attempt to measure repetitive behaviors because mice typically alternate at levels significantly above chance, indicating their willingness to explore novel environments
[74][75][74,75]. These tests do not require training, though animals need to be active and explorative. However, an important confounder of these tests is that they depend on spatial working memory
[76]. Male and female mice similarly alternate in the Y maze
[43].
Perseverative behaviors are also relatively common in mice and can be evaluated by measuring the flexibility of a mouse in terms of switching from a learned habit to a new habit. These
reversal learning tasks are usually evaluated in mazes, such as the T-maze, Morris water maze, or Clock maze
[37][77][37,77]. These tests require a substantial amount of training, and they are time consuming and cannot be used in a short age period (e.g., adolescence), although some attempts have been made to develop shorter protocols
[78]. Using the FCG model, it was shown that the sex–chromosome complement affects learning of a reversed task, with XY animals showing more perseverative errors
[79].
The insistence on exploring a known object, subject, or area over a novel example can be interpreted as analogous to the restriction in interests or insistence on sameness observed in human subjects with ASD. To explore this observation, mice have been evaluated via the novel object recognition task
[55], the social habituation/recognition task
[51], or the nose poking in a
hole board task [80]. Although males have mostly been evaluated in these tests, sex differences have been observed, with females exhibiting better performance when objects were similar in a novel object recognition test
[81].
Hypo- and hyper-reactivity to sensory stimuli can be readily evaluated in mice. Acoustic startle, air puff startle, tail flick, and hot plate can be used to evaluate adult animals. In addition, the development of sensory capacity and its response can be evaluated during the post-natal period
[43][82][43,82]. Males show a stronger startle reactivity than females
[83], and some sex differences in the nociceptive response have been reported
[84].
2.2.3. Tests for the Evaluation of Associated Symptoms
In a subset of individuals with ASD, there are associated symptoms related to other psychiatric disorders that exhibit high comorbidity with ASD, such as anxiety and depression. Various tests can be conducted to evaluate these behaviors, many of which show sex differences (reviewed in this issue,
[85]). Additionally, ASD individuals may experience seizures, intellectual disability, sleep disruption, and gastrointestinal distress. Analogous phenotypes can be assessed in mice
[86]. Seizures can be observed directly or recorded via electroencephalography (EEG). Different memory tasks that measure spatial learning (e.g., Morris water maze), contextual and cued fear conditioning, shock avoidance, object recognition, and operant tasks can be used to evaluate cognitive abilities. Running wheels and home cage monitoring systems can be used to evaluate sleep and circadian activity. Many of these parameters exhibit sex differences that should be considered when evaluating mouse models of ASD.
Evaluating associated behaviors can help to strengthen the phenotypes that correspond to the core symptom, though they can also identify potential confounders or artifacts. For instance, strong anxiety- or depression-related behaviors may lead to low exploration of social stimuli, rendering social interaction data meaningless. The same is true for memory deficits regarding social habituation or odor hyposensitivity and the observation of affected olfactory habituation or pup USVs. Deciding which phenotypes are relevant to associated symptoms and which phenotypes are artifacts that can confound the interpretation of tests related to diagnostic symptoms presents an internal contradiction that needs to be addressed on a case-by-case basis. Furthermore, these confounders may impact one sex more than the other, thus contributing to the sex bias observed in the model.
Attention-deficit/hyperactivity disorder (ADHD) is frequently observed as a comorbid condition with ASD
[87]. However, there have been prior few studies that examined the occurrence of ADHD phenotypes in rodent models of ASD. ADHD primarily affects attention, learning, hyperactivity–impulsivity, and aggressiveness
[1]. Various behavioral tests can be employed to assess behaviors associated with these symptoms
[88][89][88,89]. Deficits in learning and memory can be evaluated using the Barnes maze or the novel object recognition task. Hyperactivity can be measured through locomotion assessments in the open-field test. Impulsivity and attention deficits can be evaluated in tests such as the spontaneous Y maze alternation test or the continuous performance test. Aggressiveness is typically assessed using the resident–intruder test. Interestingly, animals that lack the integrin CD103 exhibit both ASD- and ADHD-related behaviors, and their phenotypes also exhibit sex specificity
[90]. Investigating ADHD-related behaviors in animal models of ASD can contribute to
theour understanding of the biological connections between these neurodevelopmental disorders.
2.2.4. Non-Behavioral Associated Symptoms
Mouse models of ASD recapitulate other symptoms observed in individuals with ASD, which may result from etiological factors and pathophysiological pathways and are worth investigating.
Human studies have shown the presence of activated glia, neuroinflammation, and expression of pro-inflammatory cytokines in the brains of individuals diagnosed with ASD
[91][92][91,92], as well as elevated levels of pro-inflammatory cytokines, both basally and in response to an inflammatory stimulus, in the plasma of ASD patients
[93]. Alterations in glial function, neuroinflammation, and an altered response to inflammatory stimuli have also been reported in mouse models of ASD
[67][82][67,82]. In addition, immunological dysfunction, such as T cell dysfunction, autoantibody production, and augmentation of pro-inflammatory cytokines, has been proposed in the pathogenesis of ASD (
[93][94][95][96][93,94,95,96], reviewed in
[97]). Investigating this in topic animal models has shown that cytokines can participate in the post-natal programming of adult sociability, and they can also modulate this behavior in the adult brain
[40][42][67][82][40,42,67,82]. Also, mouse models of ASD have been generated after pre- or neo-natal exposure to inflammatory stimuli (e.g., PolyI:C, LPS or virus)
[77][98][99][100][101][102][103][104][105][106][107][108][109][110][111][112][77,98,99,100,101,102,103,104,105,106,107,108,109,110,111,112]. Interestingly, sex differences in inflammatory responses and glial development and function have been reported in both humans and animals
[113].
The autistic brain is characterized by hyperconnectivity in local circuits and hypoconnectivity between brain regions
[114]. This observation is consistent with ASD being diagnosed before the age of 5, when synaptogenesis is most active in humans, a process that is next followed by active synaptic pruning and elimination
[115]. Indeed, autistic brains show increased spine density in the apical dendrites of cortical pyramidal neurons
[116], and many genes identified as providing susceptibility to ASD code for synaptic proteins or affect the morphogenesis of dendritic spines (reviewed in
[114][117][114,117]). Alterations in synaptic function and brain activity have been studied in some mouse models of ASD to understand the relevance of these findings
[117][118][119][117,118,119]. Additionally, many genetic models of ASD are built on the alteration of proteins involved in synaptic function, including neuroligins
[120][121][122][123][124][125][126][120,121,122,123,124,125,126], neurexins
[127], and shank proteins
[128][129][130][131][132][133][134][135][136][128,129,130,131,132,133,134,135,136]. As discussed below, differences in dendritic growth and synaptic formation represent a known mechanism of brain sexual differentiation.
2.3. Sex Differences in Mouse Models of ASD
Animal studies on ASD have traditionally favored male animals due to its higher incidence in boys, which has led to a lack of evidence on the impact of sex on these models, and
rwe
searchers believe that this problem may have delayed discovery in ASD. However, recent research has identified sex differences in ASD models, shedding light on possible mechanisms involved in the etiology and pathophysiology of the disorder.
ResearchersWe have included specific notations to indicate the behavioral phenotypes observed in males and females when both sexes were analyzed, and the effect of sex was considered (violet was the male symbol and was the orange female symbol). For studies in which results were obtained from both male and female subjects, but the sex effect was not reported,
researcherswe used black male and female symbols. It is important to note that when only male behavioral phenotypes are reported in a model, it indicates that
researcherswe did not come across any reports that specifically addressed females in those studies.
Among the most extensively studied pharmacological and environmental models of ASD are those generated via pre-natal exposure to valproic acid (VPA) or maternal immune activation (MIA). Various doses of VPA can be administered to animals at different gestational ages, resulting in ASD-related behaviors (as reviewed in
[137]). While most studies have used only male animals, evidence shows that VPA affects ASD-related phenotypes in males, but does not do so in females
[42][138][42,138]. For example, male mice exposed to 600 mg/kg VPA at gestational day (GD) 12.5 displayed reduced sociability in the three-chamber test, while female social interaction was not affected
[42]. However, VPA affects female mice, as they show signs of neuroinflammation during the post-natal period
[82] and in adulthood
[42]. Remarkably, repetitive behaviors were not assessed in females, and, hence, evidence of the sex-specific effect of VPA on behavior is lacking and warrants further investigation.
MIA models are generated by challenging the maternal immune system using an inflammatory stimulus. Commonly used inflammatory stimuli are the polyinosinic–polycytidylic acid (PolyI:C) that mimics viral infections and bacterial lipolysaccharides (LPS) that elicit an inflammatory response similar to the one triggered by a bacterial infection. When PolyI:C is administered at GD12, there is a consistent effect on sociability (reviewed in
[139]). Although pre-natal PolyI:C exposure can affect social behavior in both male and female mice
[107], some sex-specific differences can be observed that may depend on the gestational age at which the stimulus was administered, the strain of the mouse, and the dose and type of PolyI:C
[139][140][139,140]. Similarly, while male mice pre-natally exposed to LPS show reduced sociability, female social behavior is unaffected
[109][110][141][109,110,141]. Similar to the VPA model, LPS exposure leads to increased self-grooming in males, though this behavior has not been evaluated in females. When the human influenza virus is injected into pregnant dams at GD9.5 to elicit an inflammatory response, their adult offspring of both sexes exhibit reduced social interaction
[98].
A neonatal inflammatory challenge also results in long-lasting effects on sociability, though the extent of the effect and occurrence of sex differences depends on factors such as mouse strain, drug dose, and age at administration
[140][142][143][144][145][140,142,143,144,145].
Propionic acid (PPA) is a gut metabolite that can elicit neuroinflammatory responses
[146][147][146,147]. It has been shown that intracerebroventricular (icv), subcutaneous (sc), or intraperitoneal (ip) administration of PPA can elicit behavioral alterations related to ASD, such as reduced social interactions and repetitive patterns of behavior
[146][147][148][149][146,147,148,149]. Unfortunately, all studies on PPA have only used male subjects. However, maternal administration of PPA does not result in ASD-related behaviors in female and male offspring
[150].
Male mice pre-natally exposed to a monoclonal antibody against contactin-associated protein-like 2 (Caspr2) show reduced sociability, increased repetitive behavior (marble burying), and inflexibility in learning
[77]. However, the ASD-related behavioral phenotype of this model was not replicated when it was combined with the FCG model
[104].
Several genetic models of ASD have been proposed. Many of them are constructed based on the notion that ASD results from synaptic alterations, and, thus, key synaptic proteins, such as neuroligins 1, 3, and 4; neurexin 1α; and shank proteins, have been targeted
[120][121][122][123][124][125][126][127][128][129][130][131][132][133][134][135][136][120,121,122,123,124,125,126,127,128,129,130,131,132,133,134,135,136]. Other targeted molecules are peptides involved in social responses, such as oxytocin (OT) and vasopressin (AVP), and their receptors
[151][152][153][154][155][151,152,153,154,155]. Others models are generated by replicating genetic alterations observed in human subjects with ASD or related disorders
[156][157][158][159][160][161][162][163][164][165][166][167][168][156,157,158,159,160,161,162,163,164,165,166,167,168]. Finally, some mouse inbred strains, such as BTBR, Balb/c, and C58, exhibit reduced sociability and increased repetitive behaviors and are, thus, proposed to be studied as ASD models
[39][59][60][166][169][170][171][172][173][174][175][176][177][178][179][180][181][39,59,60,166,169,170,171,172,173,174,175,176,177,178,179,180,181]. Unfortunately, many of these models have only been studied using male subjects, while the role of sex has often not been specifically analyzed.