Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Erick Alves and Version 3 by Camila Xu.
Neglected Tropical Diseases (NTD) is a group of 20 diseases affecting more than 1 billion people in the world, especially those living in poor communities of tropical areas. Caused by a variety of etiologic agents such as viruses, bacteria, and parasites, these diseases have a great impact on public health, with social and economic consequences to the affected populations.
- covalent target enzyme inhibitors
- sleeping sickness
- Chagas disease
- Malária
1. Introduction
Neglected Tropical Diseases (NTD) is a group of 20 diseases affecting more than 1 billion people in the world, especially those living in poor communities of tropical areas. Caused by a variety of etiologic agents such as viruses, bacteria, and parasites, these diseases have a great impact on public health, with social and economic consequences to the affected populations [1]. These diseases are often carried by animal reservoirs, some of which are difficult to control. By affecting impoverished populations with little to no access to public health policies, NTDs do not attract enough interest for the pharmaceutical sector, due to the lack of financial profit. As a result, the chemotherapeutic arsenal is scarce for the majority of those diseases, and the lack of an efficient treatment leads to more than 170 million deaths per year [1]. Although this scenario has gradually improved, the investments keep going below the necessity. Research for new chemotherapeutic alternatives for these endemic health situations must be highly emphasized.
Surprisingly, new studies on covalent inhibitors and targeted covalent inhibitors (TCI) comprehend a potential new class of molecules with improved efficacy and security for NTDs. Compared to reversible inhibitors or mechanism-based inactivators, TCIs exhibit unique selectivity profiles. Additionally, while these previous methods are mainly limited to enzymes, the TCI approach is more versatile and can be employed for a wide range of druggable proteins [2].
TCIs can create a stable covalent bond with the target protein active site by positioning their moderately reactive electrophile warhead adjacent to a particular nucleophile amino acid residue, usually cysteine. Their pharmacological impact is sustained, while the unbound drug is quickly and continuously removed from the body from a pharmacokinetic perspective, which makes the latter less toxic [3][4][3,4]. TCIs have been employed for the development of new molecules for these three diseases over the past few years, leading to promising results and new alternatives for their treatment.
It is important to notice that the complex TCI-target is formed only after a molecular recognition of the ligand by its target molecule. TCIs are developed to not only have a reactive warhead to create a covalent bond but also must bear structural features for selective and specific interactions with its target; otherwise, no good inhibition is obtained [3].
2. Covalent Inhibitors for Neglected Diseases
Numerous covalent inhibitors and their corresponding warheads are examined. On that matter, Ki, k2nd, kinact are some of the parameters discussed and will be briefly introduced here. It is important to stress that covalent inhibition is a two-step procedure as pictured in Scheme 1. Some of the constants involved in the process are described below.
Scheme 1. Simplified scheme showing the formation of a covalent complex and its constants. P-I is the complex in which P and I form non-covalent interactions between them. P-I* consists of covalent binding between the two molecules.
2.1. Covalent Inhibitors Studies on Human African Trypanosomiasis (HAT)
The actual therapy for HAT is based on a few available drugs: pentamidine and suramin for first-stage HAT, while second-stage HAT is treated with eflornithine, melarsoprol, and nifurtimox (Nfx); Currently, as no efficient vaccine is available, treatments have been focused on combination therapies, but, still, several adverse effects have been observed, such as melarsoprol high neurotoxicity [6][97].2.1.1. Oxidizing Cycle
One of the most known pathways studied against HAT is the parasite oxidizing cycle since oxidative stress plays a major role in parasite survival [7][8][98,99]. Figure 1 shows this pathway; components shown in red are the ones currently being targeted for covalent inhibition explored.
Figure 1.
Trypanothione Reductase (TR)
Trypanothione reductase is the most thoroughly studied enzyme of the trypanothione metabolism, as it is not expressed in mammalian hosts, and many of its inhibitors have been identified. As the parasite does not have a glutathione reductase system, which is essential for cellular redox homeostasis [11][32], the only structure that connects NADPH and thiol-based redox systems is TR [12][33]. TR is an oxidoreductase that catalyzes NADPH-dependent reduction of trypanothione (bis(glutathionyl)spermidine) disulfide TS2 to trypanothione T(SH)2 (Scheme 2). Inhibition of this enzyme makes the parasite susceptible to oxidative stress and cellular damage.
Scheme 2.
TS
2
reduction reaction.

Trypanothione Synthetase (TryS)
Trypanothione synthetase catalyzes the synthesis of N1,N8-bis(glutathionyl)spermidine (trypanothione), the main low molecular thiol that supports many redox functionalities in trypanosomatids. Although TryS was proved to be druggable, the design of inhibitors has been challenging due to specific characteristics of this enzyme. TryS active site accommodates four substrates and contains several regions involved in substrate binding that are highly dynamic [14][35]. To explore TryS inhibition, Benítez and colleagues utilized ebselen (1), a previously identified multi-TryS molecule [9][38]. The ENAMINE library(with a true-hit rate of 0.056% and 51,624 compounds) screening identified select hits that showed significant specificity against TryS. However, only ebselen (1) displayed a balanced antitrypanosomal and TryS inhibitory activity. The researchers conducted experiments to determine its mode of action and confirmed its covalent inhibition through the formation of Se-Cys (S) bonds, primarily through MS/MS analysis.Tryparedoxin
Another step of the oxidizing cycle is catalyzed by tryparedoxin (Tpx). This enzyme is a distant member of the human thioredoxin family, and, along with T(SH)2, catalyzes the synthesis of compounds required for DNA synthesis, making it important for parasite replication. A HTS investigation by Fueler et al. led to 36 compounds that inhibited the peroxiding system by ≥20% at a concentration of 40 μM [10][39]. As the following experiments suggested, these authors proved Tpx to be an in vivo and in vitro target and characterized compounds 6, 7 and 8 (Figure 2) as time-dependent ones through a MAL-PEG-based gel shift assay. It was also observed that the presence of the chloromethyl substituent was essential for covalent binding. Tpx Cys40 likely attacks the electrophilic carbon linked to the chlorine groups in a nucleophilic substitution, facilitated by some non-covalent interactions, especially hydrogen bonds (Figure 2b). A detailed study of the mechanism of these types of molecules was then performed by [17][100].2.1.2. Rhodesain (RD)
Rhodesain is an essential protein for parasites crossing the blood brain barrier (BBB), reaching the central nervous system. It is a key cysteine protease for the survival and infectiousness of T. brucei rhodesiense. It is also predominant in lysosomes, where it exerts its protease activity. RD has demonstrated to be a validated target because of its significance for the survival of Trypanosoma sp., and several efforts, especially those linked to covalent inhibitors, have been made in the last 10 years [18][19][5,6]. Schechter and Berger nomenclature towards sites is present in the Figure 3. Combinatorial chemistry throughout the years were applied to synthesize compounds prone to bind to the sites in several papains. That way, similar trends in cruzain and rhodesain are going to be seen, for example [20][7].
Peptidic and Pseudopeptide Compounds
Kerr et al. broadened the SAR research of peptidyl vinyl Michael acceptor derivatives for RD inhibition by synthesizing a series of peptide-based compounds with the vinyl sulfones, such as K11777 [18][5], as lead compounds considering their structures cocrystallized with RD available in PDB [18][19][5,6]. Compound 9 presented Kinact of 0.00255 ± 0.00075 min−1, and a high k2nd of 67,000 ± 432 × 103 M−1 min−1, with docking studies suggesting many hydrogen bonds with the active site (Figure 4).

Figure 5. (a) Docking experiments with 19 and RD. In this figure, hydrogen bonds between several residues and 11 can be observed, and possibly facilitate the nucleophilic action of Cys25. Adapted from [26][9]; (b) Docking experiments carried out by Ettari et al. with 11 and RD [25][8]; (c) Docking experiments carried out by Previti et al. with 15 and RD [27][10]; (d) 14 and RD, active site [28][11].
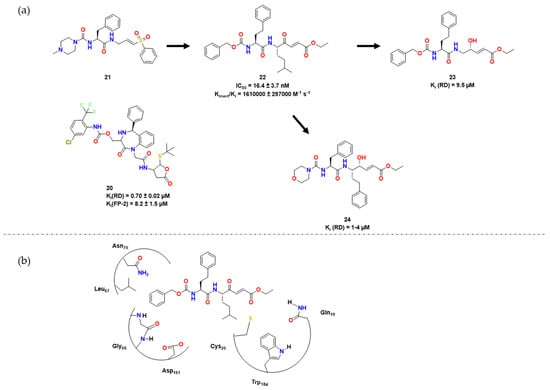


Figure 8.
Compound
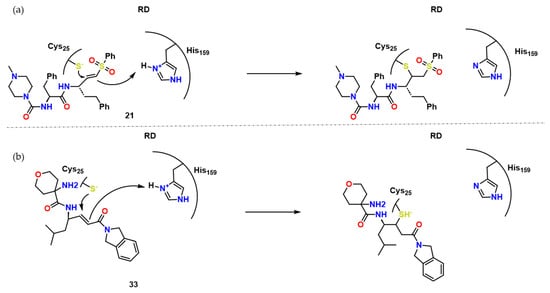

Figure 10.
Compound

Non-Peptidic Compounds
In recent years, RD inhibition has been also explored with non-peptidic compounds as an attempt to improve pharmacokinetics characteristics in comparison to what was obtained with peptidic compounds [40][25]. McShan et al. used the irreversible tethering method to explore a library of 200 electrophilic fragments that could potentially lead to covalent inhibitors [40][25]. In contrast to peptidic ones, compounds 34 and 35 were found to be non-peptidic covalent inhibitors of papain and might possess interesting pharmacokinetic properties. Additionally, these substances had low cytotoxicity and low micromolar activity against T. brucei (Figure 11). The authors also stated a possible mechanism of interaction between the covalent fragments studied and the target [41][26], which is depicted in Figure 12.




Figure 16.
Compound
2.1.3. Other Compounds and Targets Related to HAT
Pterine Reductase 1
Tetrahydrobiopterin [H4B] is a necessary cofactor for enzymes such as aromatic amino acid hydroxylases to catalyze different hydroxylation processes. H4B can be generated from GTP or rescued from dihydrobiopterin [H2B] by NADPH-dependent dihydrofolate reductase in humans or NAD[P]H-dependent 6,7-dihydropteridine reductase which regenerates quinonoid dihydrobiopterin, a byproduct of hydroxylation processes. Biological and genetic evidence suggests that trypanosomatids lack the ability to synthesize pterins de novo, and are totally dependent on the uptake of extracellular pterins for growth [43][41]. Pterin reductase 1 [PTR1] is an enzyme that reduces dihydrobiopterin in Trypanosoma spp., being essential for its survival. Over the past 10 years, papers have been published, in which pyrimidine-based compounds have been exploited for protein kinase inhibition, topoisomerase inhibition, antibacterial, anti-inflammatory and antiparasitic activity, and dipeptidyl peptidase IV inhibition. Besides, pyrrolopyrimidines had the advantage of carrying a pharmacophore similar to the recognition motif of the parasite P2 aminopurine transporter. Khalaf et al. considered 2,4-diaminopyrimidines to be important substructures at the outset of this work [44][42]. Recognizing that physicochemical properties also play an important role in the biological activity of substituted pyrimidines, they investigated 4-alkoxy and 4-alkylamino substituents. Also, this requirement for transport into trypanosomes and the possibility that the specific transporters prefer 4-aminopyridine to engage the hydrophobic pockets of PTR1 evident from crystallographic studies, 5-alkyl, 5-aryl, 6-alkyl, and 6-aryl pyrrolopyrimidines together with 5,6-disubstituted compounds were all studied (Figure 17a). Even though compounds 46 and 47 did not exhibit the best enzyme and in vitro activity, they might be investigated further for the inclusion of a formaldehyde motif, which could potentially form a reversible covalent bond to Cys168.
Figure 17. (a) Hit compounds with interesting activity towards pteridine reductase; (b) Hit compounds with interesting activity towards CLK1, adapted from [45][43]; (c) ligand interaction site taken from Crystal structure of CLK1 and 49. Adapted from [46][48]; (d) Compound 50—covalent inhibitor of CatL; (e) Hits compounds from [47][30].
Kinase CLK1
CLK1 is a crucial protein for mitosis in Trypanosoma spp., and is a target for the antibiotic hypothemycin, highlighting its potential as a drug target [48][44]. In their research, Saldivia and colleagues found a link between the inhibition of recombinant TbCLK1 and the death of the parasite [45][43]. Only compounds containing Michael acceptors were studied, leading to the identification of compound 48. To establish the significance of the Michael acceptor for inhibition activity, they synthesized 49 without the unsaturated bond, which in turn abolished compound activity (Figure 17b). Crystallographic analysis revealed that Cys215 forms a covalent bond with the Michael acceptor, while Asp218 appears to be involved in an ionic bond with an amine. Additionally, the nitrogen from Pro214 is close to the benzyl linked to pyrrole, and Tyr212 appears to form hydrogen bonds with oxygen through its backbone (Figure 17c).Cathepsin-like (TbCATL)
Two cathepsin-like proteins are of paramount importance inside the parasite: TbCATB and TbCATL. In recent studies, however, only TbCATL showed to be essential for the parasite [37][40]. Compound 50 selectively inhibited the activity of TbCATL in trypanosomes without affecting TbCATB. This molecule showed a dose-dependent inhibitory effect with an (IC50) of 0.39 µM, indicating prominent inhibition of TbCATL (Figure 18d). Steverding et al., lastly, stated that TbCATL is essential for the survival of bloodstream forms of T. brucei and this suggests that future drug development programs shall focus on the rational design of TbCATL inhibitors [37][40].
Glyceraldehyde-3-Phosphate Dehydrogenase [GAPDH] and TR
Considering the design of new drugs, molecular hybridization has been gaining importance in the last few years. Bellutti et al. aimed to modulate two vital targets in T. brucei and T. cruzi, TR and GAPDH [47][30]. For that, they considered two different lead compounds and synthesized a hybrid, leading to 10 different derivatives (One of the lead is compound 51, Figure 18e). An interesting feature present is the amine, holding a positive charge at physiological pH, which would present interaction with a pocket filled with negative residues. Compound 53 covalently inhibits the GAPDH through Cys166 and presented the best activity against the two enzymes with an IC50 (GAPDH) of 5.3 μM and Ki (TR) of 2.32 μM. It is worth mentioning though that this trend was not corroborated by in vivo activity, being one of the weakest inhibitors of the series [50][46] the best one considering in vivo activity, which would suggest off-target activity.2.1.4. Natural Compounds and Derivatives
The natural world is certainly a good source to start searching for new possible drug candidates [49][45]. In an isolated study, Oli et al. took advantage of the great structural variety of marine sponges and the fact that some of their extracts were tested against cysteine proteases [50][46]. From bioactivity-guided fractionation of the crude cyclohexane extract, they obtained plakortide E [51][47] and found out it had a IC50 of 5 μM in T. brucei and a time dependent IC50 towards RD, varying from 257 μM in 5 min to 77 μM in 60 min, which is indicative for a covalent inhibition given the presence of Michael acceptor. Considering various compounds previously reported for trypanocidal activity, as well as the recent discovery of furanoheliangolides with high antitrypanosomal activity, the interest in such compounds has increased indeed. Lenz et al. showed that the natural 4,15-isoatriplicolide type sesquiterpene lactones inhibited trypanothione reductase; particularly, 4,15-isoatriplicolide tiglate exhibited exceptionally potent activity against T. brucei rhodesiense with an IC50 value of 15 nM [46][48]. In 2019, this same group synthesized compound 55, which also proved to be an interesting compound, presenting TbTR inhibitory activity of more than 10% in only 15 min. Zhang et al., on the other hand, approached natural phenyl vinyl sulfone compounds-derivatives [24][31]. They observed compound 56 to be inactive against RD at 20 µM after 1 h incubation, Kinact/Ki of 99 M−1 s−1 and to show reasonable in vitro activity, with an IC50 of 5.97 µM. Another interesting feature was its inactivity towards human analogous cathepsin L. Cys25 was discovered to be adjacent to the vinyl moiety in docking investigations. The homophenyl is near the P1 site, while the quinoline is next to P2. The phenyl moiety was predicted to interact with Gln19 and His162, while quinolyl had hydrophobic interactions with Met68. On the other hand, P3 did not have a moiety of its own. Thus, regarding these experiments, it is of interest to highlight the synthesis of compound 57, which presented interesting in vivo antitrypanosomal activity, and 58, which presented a great selectivity index of over 159 (Figure 19).
Figure 19. Distribution of pockets that will be used in this part of review. Also, it is noted that the parts of molecule which are nearest to these pockets are P1, P2, and P3. pdb: 4QH6.
2.2. Covalent Inhibitors Studies on Chagas Disease [CD]
2.2.1. Cruzain [CZ]
Cruzain is the largest cysteine protease from T. cruzi, and it is associated with many crucial biological processes inside the parasite, such as differentiation, invasion, and proliferation in host cells [52][49]. The sites most explored regarding drug discovery and this enzyme are S1, S2, and S3, as shown in Figure 19.Peptidomimetics
One of the major efforts in assessing drug design against CD and CZ inhibition involves peptidomimetic compounds. The use of these structures comes from the fact that cysteine proteases target protein-like structures. CZ, as a cysteine protease per se, is not different, and using this scaffold as a lead has allowed researchers to design many potential inhibitors [53][50]. Avelar et al. used a molecule established by [54][51] as a lead (59, Figure 20), and explored nitrile as a warhead for TCI against CZ [55][52]. After many modifications in P1, P2 and P3 (as seen in Figure 18), they synthesized a derivative containing 1-methyl-3-tert-butyl pyrazole in P3 and chlorine in meta for P2 (compound 63, pKi = 7.2). Furthermore, crystallographic experiments revealed that phenylalanine did not penetrate as effectively as m-chlorine in pocket S2. The authors determined that efficient penetration of S2 was only achievable when 3-chloro and 1-methyl-3-pyrazole groups were present. Despite finding this information, they were unable to demonstrate a substantial structure-activity link for amastigote forms.

Figure 21.
Proposed mechanism of covalent inhibition concerning nitrile compounds such as the ones shown in

Figure 22.
Covalent docking experiments with

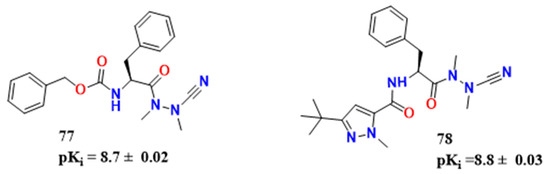
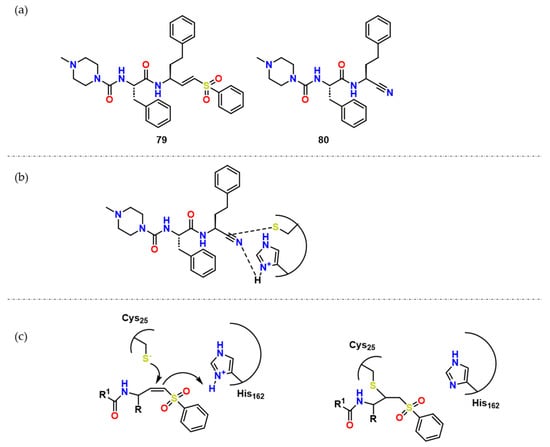
Figure 25. (a) Compound 79 and its analogue 80; (b) Possible network of interactions regarding compound 80; (c) Possible mechanism of action of covalent inhibitors and CZ.
Non-Pepditic Compounds
Non-peptide compounds have been also studied by Jimenez et al. They explored N-acyhydrazones (NAH) and benzenesulfonyl (BS) as groups with potential antitrypanosomal activity totaling 14 compounds designed [66][63], obtaining compounds with activity, having in common the presence of strong electron-withdrawing groups in para-position on the benzene ring. Maldonado et al. substituted the sulfonyl by carbonyl group to obtain new N-propionyl N′-benzeneacylhydrazone derivatives (83 and 84) and evaluated their trypanocidal activity (Figure 26) [67][64]. As a result, compound 83 presented an interesting activity against NINOA strain, and compound 84 presented LC50 comparable to Bzn.
Natural Compounds
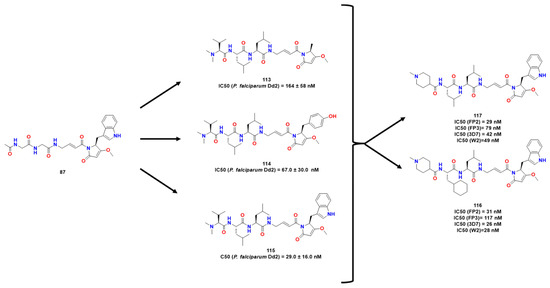
Silva et al. explored naphthoquinone analogs known to exhibit many biological properties, such as anticancer and antitrypanosomal [70][65]. In this work, they performed a virtual screening on CZ and RD. Thus, 14 molecules were identified as potentially active and were further synthesized. Compound 87 (Figure 27) was a hit compound known as a time-dependent inhibitor, and active in CZ and RD as compounds 88 and 89.
2.2.2. Other Targets Related to CD
Proline Racemase [PRAC]
Proline racemase catalyzes the interconversion of L- and D-proline enantiomers [71][66] with two different residues directly involved in isomerization: Cys130 and Cys270 [72][67]. This is important for several processes in cell function, including T. cruzi wall construction and immune escape. Also, for its presence in every single stage of T. cruzi cycle and since the parasite is no longer viable after knocking this gene down and gets more virulent if it is overexpressed, this target can be promising [73][68]. After identifying several isoforms of T. cruzi proline racemases, Berneman et al. chose 2-pyrrolecarboxylic acid (PYC) as a starting point as it is a known PRAC inhibitor with a deficit in water solubility [72][67]. This latter feature was the focus of the group and led to the synthesis of several pyrazole inhibitors that showed to be more soluble, but that did not present a good affinity for PRAC. In another approach, they performed virtual screening experiments and constructed a QSAR model, leading them to two novel, poorly reactive Michael acceptor compounds: OxoPa (95) and BrOxoPA (96) (Figure 28).
Figure 28.
Compounds
95
–
Trypanothione Reductase (TR)
TR is an essential enzyme for the trypanothione-based thiol metabolism, as stated before for Trypanosoma brucei. Beig et al. employed in vitro and in silico drug discovery methodologies, and 82 novel inhibitors showed activity in the range of nanomolar against T. cruzi TR (Figure 29) [76][71]. The best three hits were also relatively active in cell culture (IC50 < 2 μM, comparable to Nfx) and shared an aromatic core ring. Two hits (101 and 102) are shown in Figure 30, and the authors performed experiments that suggested these molecules might act as reversible inhibitors of TR. More details about TR and the structural insights of this protein are described in the reviews written by Tiwari et al. [16][37] and Battista et al. [77][72].

Figure 30.
Compounds
103
to
2.3. Malaria
P. falciparum has developed resistance to almost all antimalarial drugs used in prevention and treatment. Although most cases are in underdeveloped countries, globalization has made it possible for malaria to spread specially when left uncontrolled [80][95].2.3.1. PfGAPDH
GAPDH catalyzes the conversion of glyceraldehyde 3-phosphate (G3P) into 1,3-biphosphoglycerate (1,3-BPG), which is the sixth stage of glycolysis, reducing NAD+ to NAD through a catalytic Cys153. Its potential as a parasitic target comes from the fact that, mainly at the amastigote stage, glycolysis makes up every amount of energy storage for the parasite [78][83]. PfGADPH is inactivated by compounds capable of alkylating thiols such as 3-bromo-isoxazolines. In this regard, Bruno and coworker decided to analyze the potential activity of avicin and Br-avicin, as well as some of their analogs, and how strong and effective bromine warheads were for PfGADPH inhibition [78][83]. From their series, two compounds presented the best results: 103 and 104 (Figure 30), with the latter presenting the best covalent parameters (Kinact/Ki = 10.7 ± 2.8). Docking experiments suggested that the isoxazoline moiety, which was near His180 and Tyr323, probably interacted with NAD+ through π-stacking interactions (Figure 31). Furthermore, carbonyl oxygen carried out another hydrogen interaction with a ribose hydroxyl group, while the amine group is over sugar without any other contact. Cys153 was responsible for the nucleophilic attack, later confirmed by UV-vis spectrophotometric detection. It is worth to note that while the glycolytic function of PfGAPDH is inhibited by heme binding this function in enzyme from mammalian is not, which is indicative of difference in the catalytic center [79][81][84,85].
Figure 31. Molecule 103 in the active site post-docking carried out by [78][83]. There are π-stacking interactions between Tyr123 and His1na with molecule 103, which facilitates the nucleophilic attack of Cys153 on carbon 3, with the subsequent leave of bromine. 103 possible interactions with PfGAPDH, as shown in docking studies [4].
2.3.2. Falcipains
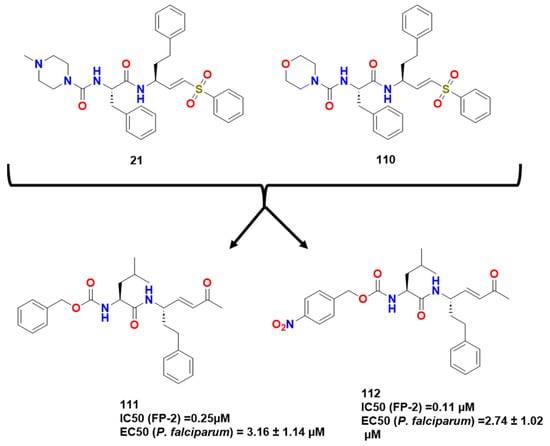
There are four papain-like cysteine proteases characterized for P. falciparum named falcipains, FP-1, FP-2 and FP-2′, FP-3. FP-2 and FP-3 diverge by 37% in amino acid sequence and are expressed in the acidic vacuoles of schizonts and trophozoites. Studies have shown that inhibiting these two latter enzymes prevents hemoglobin hydrolysis and parasite development. Also, knocked-out experiments concerning FP-2 led to the intermittent hindrance of hemoglobin hydrolysis in trophozoites, which made these parasites susceptible to inhibitors, although metabolic recovery was observed due to the probable expression of FP-3 [84][75]. In light of the studies of these targets, Ettari et al. have conducted a research on the construction of new peptidomimetics with a 1,4-benzodiazepine scaffold introduced into a dipeptide moiety, simulating a sequence of peptides necessary to bind mainly with FP-2 (Figure 32) [84][75]. From previous studies, Ettari et al. studied the effects of modifications in some parts of the structures present in 108 [86][76]. The methyl group at P1′ was replaced, which maintained irreversible inhibition but significantly reduced affinity and inhibitory potency. This difference was attributed to the divergence in lipophilicity of the compounds. Docking experiments provided further explanation, and the k2 values displayed by the compounds indicated a disfavoring of covalent adduct formation. While the carbonyl group of these compounds seemed to interact with the Trp206 side chain through hydrogen bonding, the P1′ substituent showed a different orientation. In compound 108, the methyl group was directed towards Trp206, whereas in compound 109, the groups were in proximity to residues Asn173, Val152, and Trp206, respectively, and the small distance between double bond and the sulfur atom indicating possible Michael addition. The removal of chlorine from the distal aryl moiety resulted in a derivative with reversible activity and low affinity for falcipain. Molecular dynamics studies suggested that the presence of chlorine may have contributed to a more stable conformation of compound 108 [86][76]. Previti et al. continued these lines of research by synthesizing a new series of peptide-based compounds as potential FP-2 and RD inhibitors, using compound 21 and k11002 (110) as lead compounds (Figure 33) [87][15]. The interaction of the compounds with FP-2 was lower than that presented for RD as cited previously here. They then tested several warheads and observed a trend of reactivity that followed this order: ketones > sulfones > esters > nitriles, with the vinyl ketones presenting the most potent inhibitors studied by Previti et al. (111 and 112; IC50 of 0.25 and 0.11 μM/EC50 of 3.16 ± 1.14 and 2.74 ± 1.02 μM P. falciparum respectively) (Figure 34).
Natural Compounds
With the purpose of taking advantage of natural compounds bearing privileged structures, Conroy et al. reported the synthesis and biological assay of gallinamide A [89][92] after its previous isolation [89][92]. From these efforts, the authors demonstrated interesting in vitro activity against P. falciparum with an IC50 of 50 nM of compound 90 and no harmful effect on red blood cells. They observed that replacing N-acyl pyrrolidine group had a negative impact on activity against FPs and parasites and assessed whether one or both olefinic moieties were necessary for the inhibition. They observed that the reduction in acyl pyrrolidine or in the α,β-unsaturated portion caused a detrimental effect in antiplasmodial activity. Also, the substitution of N-acyl pyrrolidine with other C-terminal groups results in a loss of activity. The derivatives in Figure 35 presented interesting activity against P. falciparum. Substitution of N-acyl pyrrolidine was attempted with benzylamide, although it was not successful. They attributed the importance of this moiety to a Michael acceptor for the active-site Cys residues in FPs in comparison with the corresponding amide. Chain extension was also attempted but failed due to the loss of activity [89][92].
Figure 35.
Figure showing compound
2.3.3. Proteasomes
Proteasomes are responsible for regulating specific processes within the cell cycle. Its composition consists of 20S catalytic units, which embrace 2 heptameric rings of beta subunits. The β1 unit behaves as caspase-like structures (ceasing itself after acidic residues), while the β2 subunit presents trypsin-like activity (cleaves after basic residues) and β5 subunit, chymotrypsin-like activity [92][80]. Proteasome inhibitors have potential activity against Malaria. Also, they have been discovered to be the cause of apoptosis in certain tumor-derived cells, which made their appearance in investigations against some types of cancer [92][80]. Furthermore, they can be active against artemisinin-sensitive and resistant parasites.Benzoxaboroles
In past endeavors, epoxyketone and vinyl sulfone warhead-based inhibitors were synthesized and bound irreversibly to proteasome active sites. Bortezomib and ixazomib are two reversible boronate-based inhibitors used clinically to treat multiple myeloma. Bortezomib presents activity against P. falciparum, albeit its activity is not entirely linked to proteasome inhibition. In addition, boronic acid derivatives are known to be serine proteases covalent inhibitors [93][81] and, usually, boronate peptides are considered for their properties as covalent inhibitors as shown by Groll et al. [92][80]. For this reason, Xie et al. used direct evolution and screening from a boronate peptide library of human proteasome inhibitors from Takeda Oncology Company to identify inhibitors of the growth of P. falciparum in vitro [90][78]. Four compounds had good antiplasmodial activity (Figure 36), but compound 121 presented the best selectivity against β2 site comparing plasmodium and human structures (Ki (Pfβ2) = 6.4 nM/Ki (Hβ2) = 487 nM). Compound 120 presented similar t½ at the human β5c site (110 min) concerning dissociation from β5 site (106 min, Ki = 0.1 nM], but only 2 min (Ki = 490 nM) at the B2c site, indicating its preference for β5c site. As opposed to it, 120 had an interesting affinity against β2 and β5 sites of Pf20S (~5 nM) and longer t1/2 (~65 min). After crystallographic assays with pf20s and bortezomib, a covalent bond between threonine and boronate was observed (Figure 36). The boronic acid derivative moiety gave hard-atom oxygen a good spot to react with as Lewis hard-soft theory states. In addition, the amine present in the threonine performed hydrogen interactions with hydroxyl from boronate, and the Gly47 also performed this interaction (Figure 36). Following these studies, Xie and collaborators attempted to enhance the oral bioavailability of MPI-1 (compound 118) and synthesized a series of compounds with a single amide-bond backbone [90][91][78,79]. As a result, they obtained 122 (Figure 36), an orally bioavailable antimalarial proteasome inhibitor with in vivo efficacy. As drawbacks and points of improvement, it was observed enhancement of in vivo half-life and possible inhibition of human T cell activation. Nevertheless, this series demonstrated the potential for a proteasome inhibitor to be used for Malaria in treatment, transmission-blocking, and chemoprophylaxis scenarios, especially when resistance to artemisinin limits the efficacy of existing combinations. In a cryo-electron microscopy (EM) experiment, the same authors observed interesting contacts between 121 and the active site of Pf20S (Figure 35). The continuous density observed between Thr1 and boronate derived structures was the main feature observed. Another point to highlight was the great number of hydrophobic interactions, particularly near the biphenyl ring. As it was observed in these assays, the size of the P1 substituent was directly related to potency and selectivity. When phenyl was replaced by biphenyl, 121 was a 5 nM inhibitor of Pfβ5 and a 21-nM inhibitor of 3d7 culture growth. As it was observed, this series presented favorable physico-chemical properties. For further studies, 121 and 122 were assessed for in vivo efficacy. Also, cryo-EM was performed in combination with MPI-5 and bortezomib. From this structure, it could be inferred that MPI-5 performed mainly hydrophobic interactions, especially considering biphenyl moiety, which probably accounted for the gain in potency in comparison with derivatives that only present monophenyl structure. Also, from further characterization, it was observed that the covalent bonding to bortezomib is 17 times faster that the one characterized by 121 for HsB5c, while the latter residence time is nearly half of the bortezomib in Hs20s. As opposed to this, the rate of dative bond formation on Pf20S is higher than for bortezomib and similar for bortezomib and Hs20 beta 5C, which is consistent with a 14-fold higher inhibition of bortezomib with Hs20s beta 5c and 2.2-fold less potent pf20s beta5 in comparison to 121. In conclusion, they could infer that 122 is a good example of orally bioavailable antimalarial compound presenting in vivo efficacy. However, it would be interesting to improve its half-life in vivo further. In a separate study, Li et al. used liquid chromatography tandem mass spectrometry to screen 228 synthetic tetradecapeptides [94][101]. This resulted in 124, in which they discovered a density continuity between this compound and two of P. falciparum 20S, as well as significant selectivity over the human enzyme, and 123 (Figure 37). These compounds explore bulky SAR in many positions in β2 and β5 subunit active sites of the proteasome.


Fk506-Binding Protein 35 (PfFKBP35)
The recruitment of PfFKBP35 is essential for immunosuppression carried out by inhibitors of calcineurin and mTOR [96][88]. Bianchin et al. offered insights through crystallographic studies of the interactions of PfFKBP-rapamycin [85][89]. They mentioned the differences between Plasmodium and human FKBP, highlighting the presence of Cys106 and Ser109 instead of His87 and Ile90 in humans (Figure 39). As a result, they confirmed that Cys106 might be investigated as a particular target for covalent inhibition, which was then done through QM/MM research by [97][90].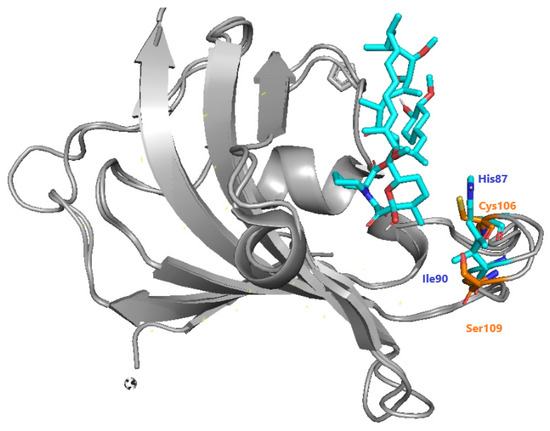
Figure 39. Specific differences between hFKBP (blue) and PfFKBP (orange), adapted from Bianchin et al. [85].
